

Comparative Analysis of Maternal Colostrum and Colostrum Replacer Effects on Immunity, Growth, and Health of Japanese Black Calves


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Colostrum Feeding
2.3. Feeding Management
2.4. Sampling and Purification of Peripheral Blood Mononuclear Cells (PBMCs)
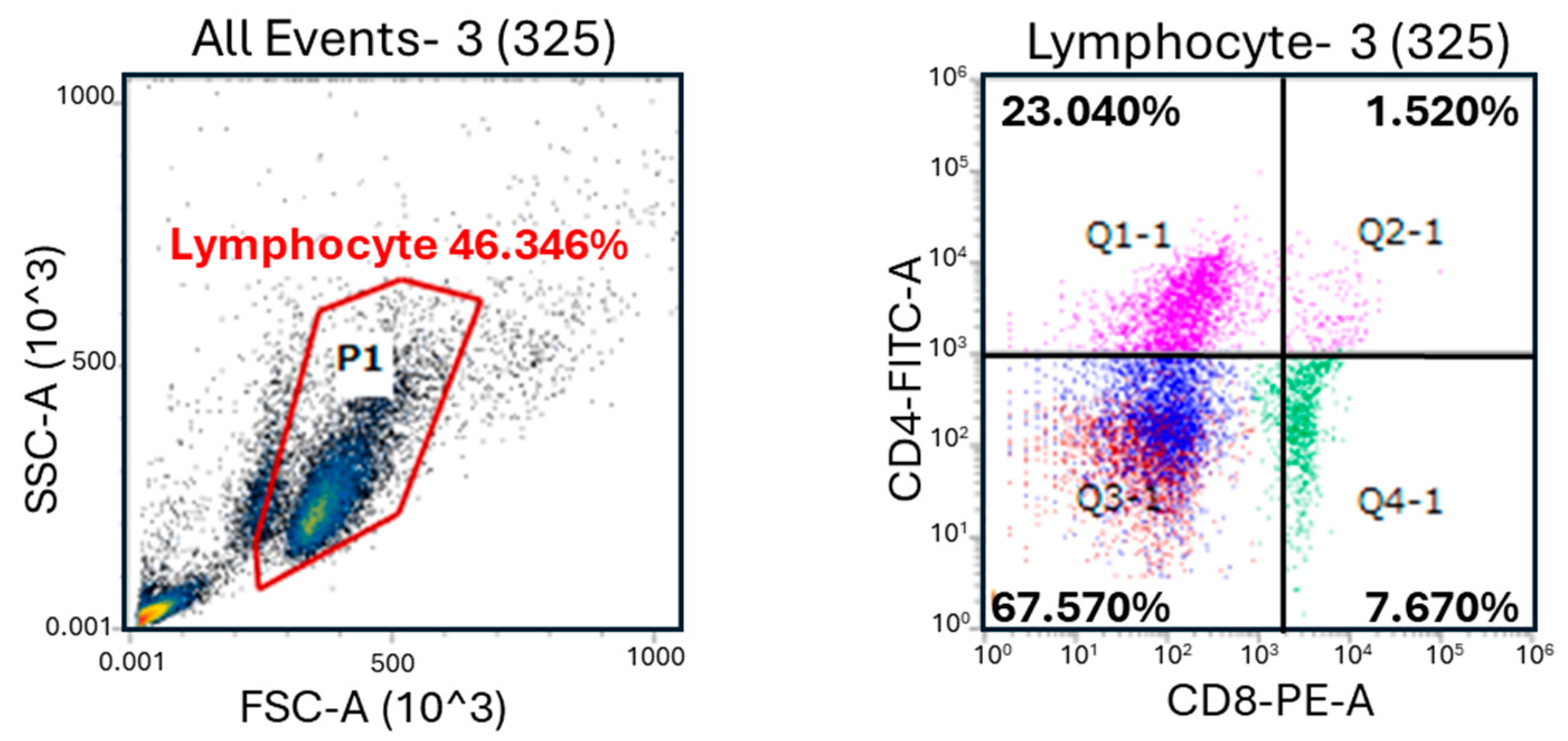
2.5. Lymphocyte Populations
2.6. Calculation of the Phagocytosis Index
2.7. Proliferation of Lymphocytes
2.8. RNA Extraction and Expression of mRNAs
2.9. Statistical Analysis
3. Results
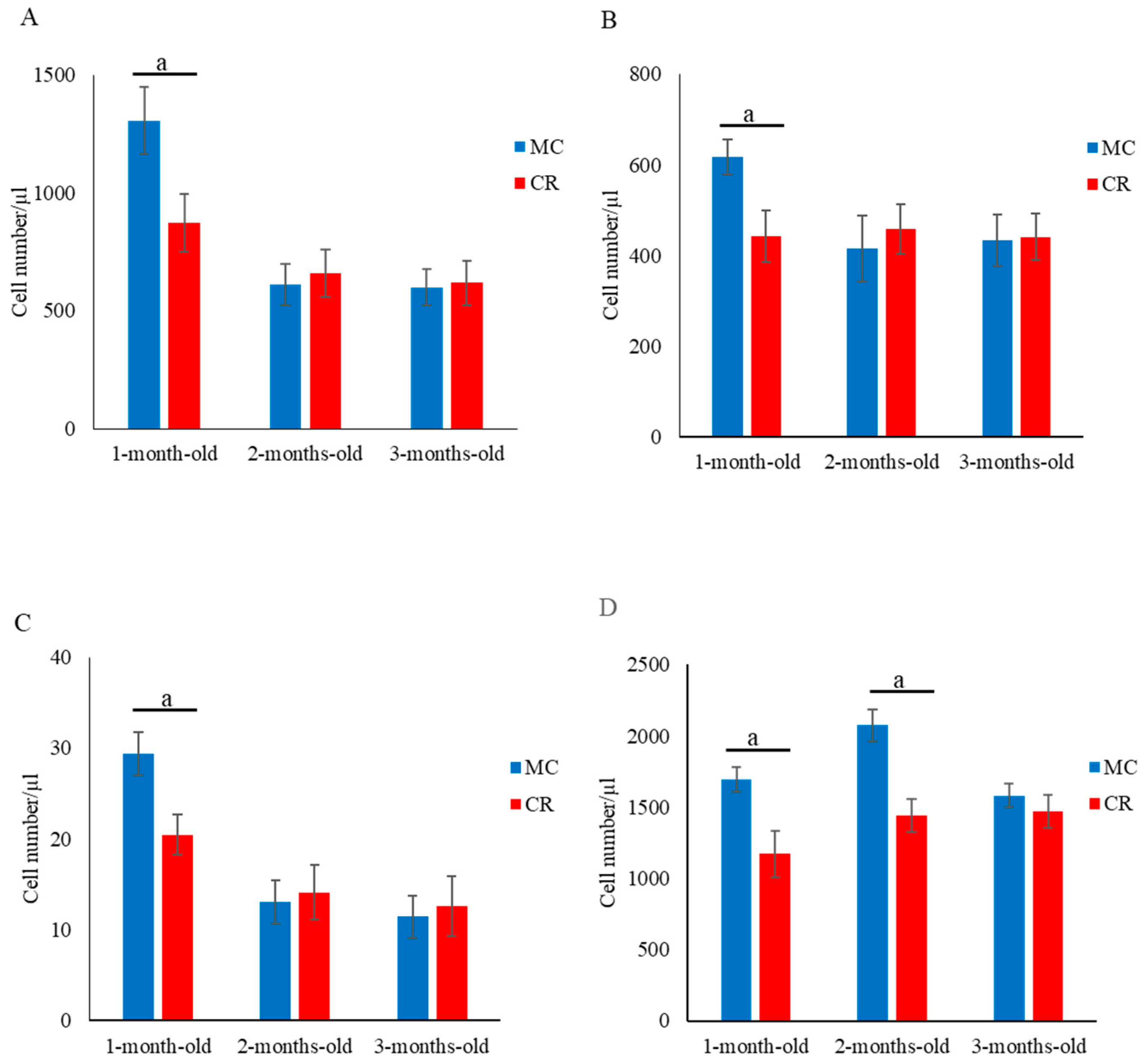
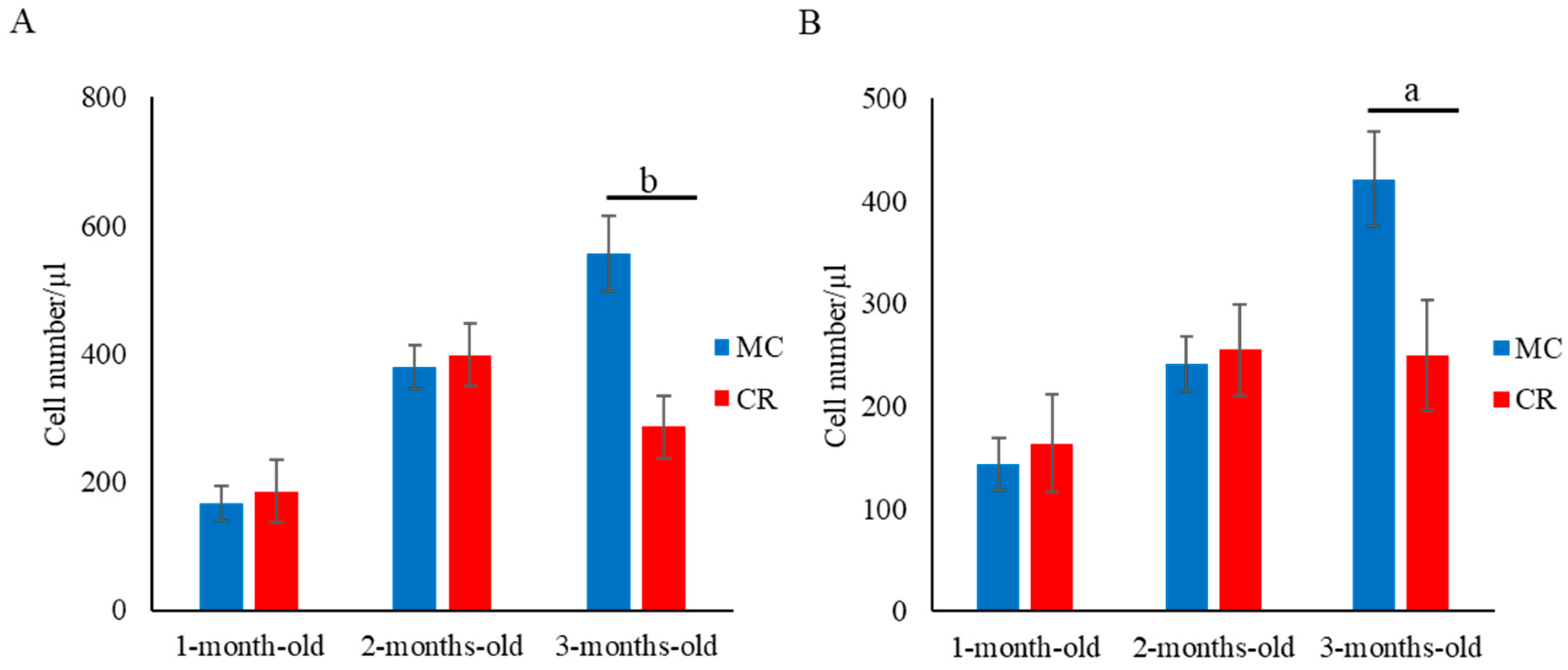
3.1. Lymphocyte Subset Analysis
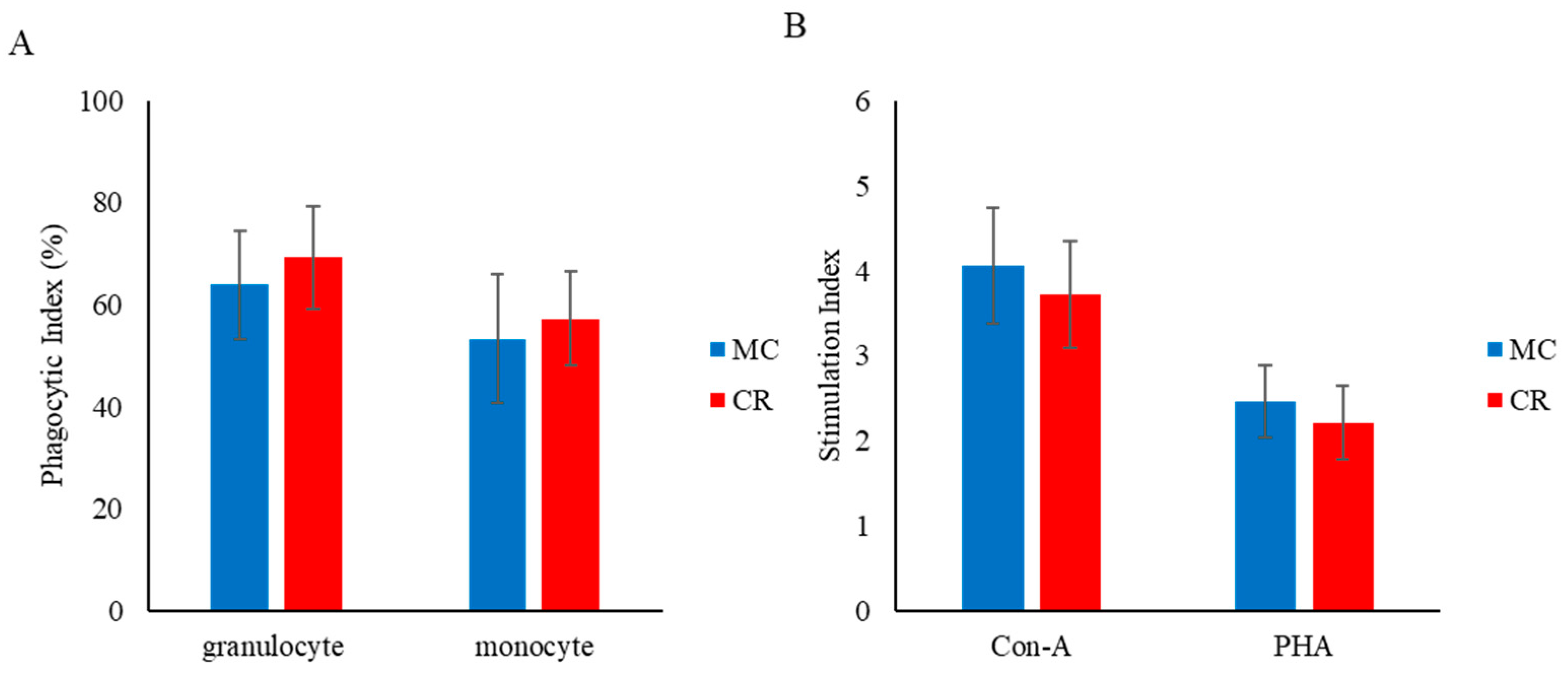
3.2. Phagocytosis and Lymphocyte Proliferation
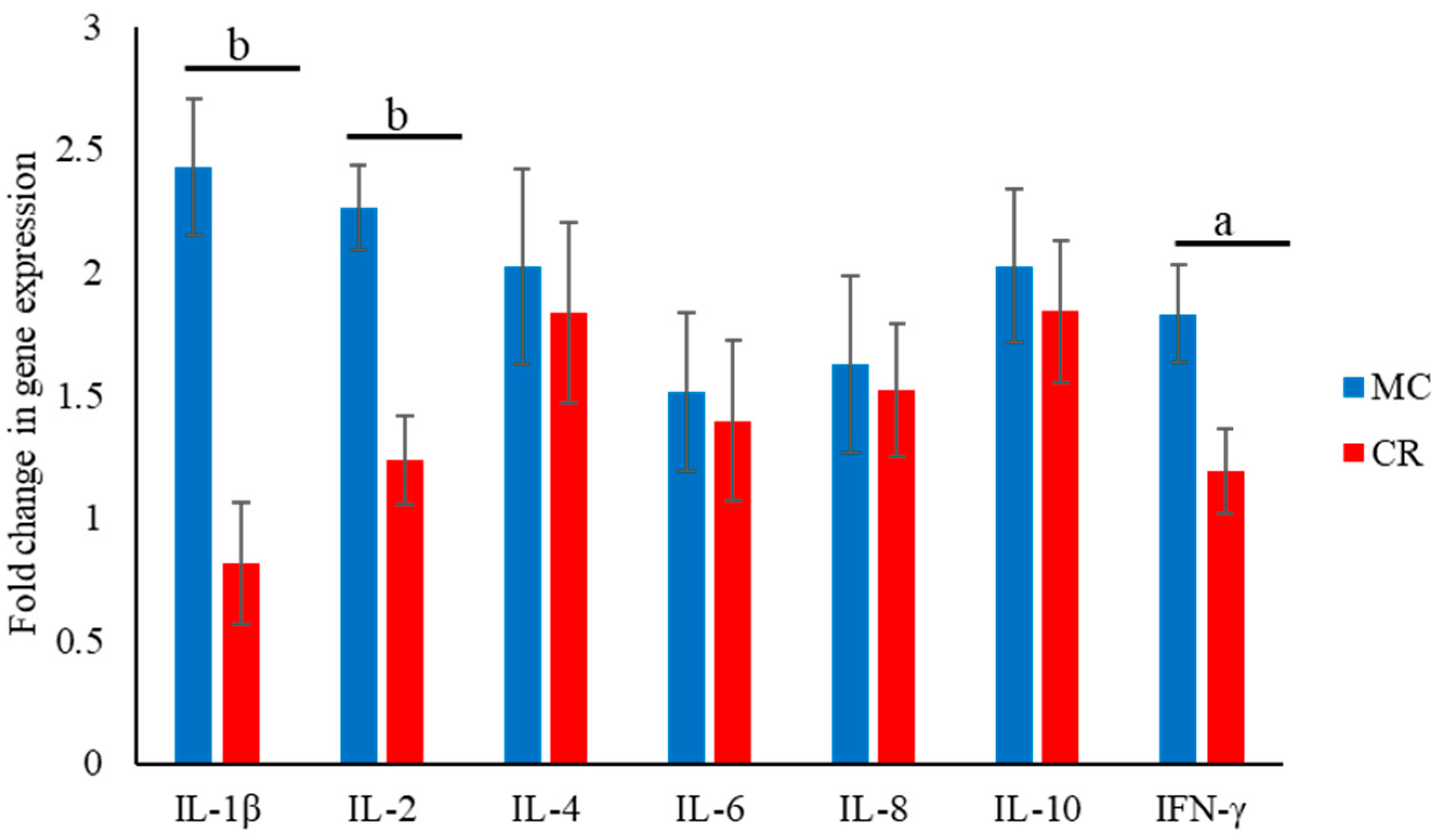
3.3. Expression of Cytokine-Encoding mRNAs
3.4. Respiratory Disease Treatment and Carcass Weight Measurement
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weaver, D.M.; Tyler, J.W.; VanMetre, D.C.; Hostetler, D.E.; Barrington, G.M. Passive Transfer of Colostral Immunoglobulins in Calves. J. Vet. Intern. Med. 2000, 14, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Faber, S.N.; Faber, N.E.; Mccauley, T.C. Effects of Colostrum Ingestion on Lactational Performance. Prof. Anim. Sci. 2005, 21, 420–425. [Google Scholar] [CrossRef]
- Donovan, D.C.; Reber, A.J.; Gabbard, J.D.; Aceves-Avila, M.; Galland, K.L.; Holbert, K.A.; Ely, L.O.; Hurley, D.J. Effect of Maternal Cells Transferred with Colostrum on Cellular Responses to Pathogen Antigens in Neonatal Calves. Am. J. Vet. Res. 2007, 68, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Mallard, B.A.; Dekkers, J.C.; Ireland, M.J.; Leslie, K.E.; Sharif, S.; Vankampen, C.L.; Wagter, L.; Wilkie, B.N. Alteration in Immune Responsiveness during the Peripartum Period and Its Ramification on Dairy Cow and Calf Health. J. Dairy Sci. 1998, 81, 585–595. [Google Scholar] [CrossRef]
- McGuirk, S.M.; Collins, M. Managing the Production, Storage, and Delivery of Colostrum. Vet. Clin. N. Am.—Food Anim. Pract. 2004, 20, 593–603. [Google Scholar] [CrossRef]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum Management for Dairy Calves. Vet. Clin. N. Am.—Food Anim. Pract. 2019, 35, 535–556. [Google Scholar] [CrossRef]
- Godden, S.M.; Haines, D.M.; Hagman, D. Improving Passive Transfer of Immunoglobulins in Calves. I: Dose Effect of Feeding a Commercial Colostrum Replacer. J. Dairy Sci. 2009, 92, 1750–1757. [Google Scholar] [CrossRef]
- Swan, H.; Godden, S.; Bey, R.; Wells, S.; Fetrow, J.; Chester-Jonest, H. Passive Transfer of Immunoglobulin G and Preweaning Health in Holstein Calves Fed a Commercial Colostrum Replacer. J. Dairy Sci. 2007, 90, 3857–3866. [Google Scholar] [CrossRef]
- Jones, C.M.; James, R.E.; Quigley, J.D.; McGilliard, M.L. Influence of Pooled Colostrum or Colostrum Replacement on IgG and Evaluation of Animal Plasma in Milk Replacer. J. Dairy Sci. 2004, 87, 1806–1814. [Google Scholar] [CrossRef]
- Foster, D.M.; Smith, G.W.; Sanner, T.R.; Busso, G.V. Serum IgG and Total [Protein] in Dairy Calves Fed Two Colostrum Replacement Products. J. Am. Vet. Med. Assoc. 2006, 27606, 1282–1285. [Google Scholar] [CrossRef]
- Pithua, P.; Godden, S.M.; Wells, S.J.; Oakes, M.J. Efficacy of Feeding Plasma-Derived Commercial Colostrum Replacer for the Prevention of Transmission of Mycobacterium avium Subsp paratuberculosis in Holstein Calves. J. Am. Vet. Med. Assoc. 2009, 234, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; LeBlanc, S.J. Factors Associated with Morbidity, Mortality, and Growth of Dairy Heifer Calves up to 3 Months of Age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, K.P.; Foley, A.L.; Collins, M.T.; McGuirk, S.M. Comparison of Passive Transfer of Immunity in Neonatal Dairy Calves Fed Colostrum or Bovine Serum-Based Colostrum Replacement and Colostrum Supplement Products. J. Am. Vet. Med. Assoc. 2010, 237, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Chigerwe, M.; Tyler, J.W.; Schultz, L.G.; Middleton, J.R.; Steevens, B.J.; Spain, J.N. Effect of Colostrum Administration by Use of Oroesophageal Intubation on Serum IgG Concentrations in Holstein Bull Calves. Am. J. Vet. Res. 2008, 69, 1158–1163. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, C.; Oie, H.; Mashimo, R.; Goto, A.; Kusaba, N.; Yamagishi, N. The Effects of Maternal Supplementation of Rumen-Protected Lysine during the Close-up Dry Period on Newborn Metabolism and Growth in Holstein Calves. J. Anim. Physiol. Anim. Nutr. 2023, 107, 28–36. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Beef Cattle, 8th ed.; National Academies Press: Washington, DC, USA, 2015; ISBN 978-0-309-31702-3. [Google Scholar]
- Baakhtari, M.; Imaizumi, N.; Kida, T.; Yanagita, T.; Ramah, A.; Ahmadi, P.; Takebe, N.; Iwamoto, Y.; Korosue, K.; Tsuzuki, N.; et al. Effects of Branched-Chain Amino Acids on Immune Status of Young Racing Horses. J. Vet. Med. Sci. 2022, 84, 558–565. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hagiwara, K.; Yamanaka, H.; Higuchi, H.; Nagahata, H.; Kirisawa, R.; Iwai, H. Oral Administration of IL-1β Enhanced the Proliferation of Lymphocytes and the O2 Production of Neutrophil in Newborn Calf. Vet. Immunol. Immunopathol. 2001, 81, 59–69. [Google Scholar] [CrossRef]
- Yamanaka, H.; Hagiwara, K.; Kirisawa, R.; Iwai, H. Transient Detection of Proinflammatory Cytokines in Sera of Colostrum-Fed Newborn Calves. J. Vet. Med. Sci. 2003, 65, 813–816. [Google Scholar] [CrossRef]
- Yamanaka, H.; Hagiwara, K.; Kirisawa, R.; Iwai, H. Proinflammatory Cytokines in Bovine Colostrum Potentiate the Mitogenic Response of Peripheral Blood Mononuclear Cells from Newborn Calves through IL-2 and CD25 Expression. Microbiol. Immunol. 2003, 47, 461–468. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The Multifaceted Role of CD4+ T Cells in CD8+ T Cell Memory. Nat. Rev. Immunol. 2016, 16, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Nascimbeni, M.; Shin, E.C.; Chiriboga, L.; Kleiner, D.E.; Rehermann, B. Peripheral CD4+CD8+ T Cells Are Differentiated Effector Memory Cells with Antiviral Functions. Blood 2004, 104, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Quandt, D.; Rothe, K.; Scholz, R.; Baerwald, C.W.; Wagner, U. Peripheral CD4CD8 Double Positive T Cells with a Distinct Helper Cytokine Profile Are Increased in Rheumatoid Arthritis. PLoS ONE 2014, 9, e93293. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Cai, B.; Feng, W.; Yang, B.; Huang, Z.; Zuo, C.; Wang, L. Double Positive CD4+CD8+ T Cells: Key Suppressive Role in the Production of Autoantibodies in Systemic Lupus Erythematosus. Indian J. Med. Res. 2014, 140, 513–519. [Google Scholar] [PubMed]
- Diedrich, C.R.; Gideon, H.P.; Rutledge, T.; Baranowski, T.M.; Maiello, P.; Myers, A.J.; Lin, P.L. CD4CD8 Double Positive T Cell Responses during Mycobacterium Tuberculosis Infection in Cynomolgus Macaques. J. Med. Primatol. 2019, 48, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Hein, W.R.; Mackay, C.R. Prominence of Γδ T Cells in the Ruminant Immune System. Immunol. Today 1991, 12, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, P.; Splitter, G.A. Γδ+ T Lymphocyte Cytotoxicity against Envelope-Expressing Target Cells Is Unique to the Alymphocytic State of Bovine Leukemia Virus Infection in the Natural Host. J. Virol. 2000, 74, 8299–8306. [Google Scholar] [CrossRef]
- Murakami, K.; Sentsui, H.; Inoshima, Y.; Inumaru, S. Increase in Γδ T Cells in the Blood of Cattle Persistently Infected with Bovine Leukemia Virus Following Administration of Recombinant Bovine IFN-γ. Vet. Immunol. Immunopathol. 2004, 101, 61–71. [Google Scholar] [CrossRef]
- Guzman, E.; Price, S.; Poulsom, H.; Hope, J. Bovine Γδ T Cells: Cells with Multiple Functions and Important Roles in Immunity. Vet. Immunol. Immunopathol. 2012, 148, 161–167. [Google Scholar] [CrossRef]
- Ohtsuka, H.; Fukunaga, N.; Hara, H.; Fukuda, S.; Hayashi, T.; Hoshi, F.; Yoshino, T.; Koiwa, M.; Kawamura, S. Changes in Peripheral Leukocyte Populations of Weak Calf Syndrome of Japanese Black Calves. J. Vet. Med. Sci. 2003, 65, 793–796. [Google Scholar] [CrossRef]
- Wyatt, C.R.; Madruga, C.; Cluff, C.; Parish, S.; Hamilton, M.J.; Goff, W.; Davis, W.C. Differential Distribution of ΓδT-Cell Receptor Lymphocyte Subpopulations in Blood and Spleen of Young and Adult Cattle. Vet. Immunol. Immunopathol. 1994, 40, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A.; Zolnai, A.; Rudas, P.; Frenyo, L.V. T-Cell Subsets in Blood and Lymphoid Tissues Obtained from Fetal Calves, Maturing Calves, and Adult Bovine. Vet. Immunol. Immunopathol. 1996, 53, 49–60. [Google Scholar] [CrossRef]
- Nonnecke, B.J.; Horst, R.L.; Waters, W.R.; Dubeski, P.; Harp, J.A. Modulation of Fat-Soluble Vitamin Concentrations and Blood Mononuclear Leukocyte Populations in Milk Replacer-Fed Calves by Dietary Vitamin A and β-Carotene. J. Dairy Sci. 1999, 82, 2632–2641. [Google Scholar] [CrossRef] [PubMed]
- Maidment, D.C. Changes in Immunoglobulin Levels during Storage of Fermented Bovine Colostrum. Br. Vet. J. 1982, 138, 18–22. [Google Scholar] [CrossRef]
- Eckblad, W.P.; Hendrix, K.M.; Olson, D.P. Total Complement Hemolytic Activity of Colostral Whey and Sera from Dairy Cows. Cornell Vet. 1981, 71, 54–58. [Google Scholar]
- Tsuji, S.; Hirata, Y.; Matsuoka, K. Two Apparent Molecular Forms of Bovine Lactoferrin. J. Dairy Sci. 1989, 72, 1130–1136. [Google Scholar] [CrossRef]
- Kishikawa, Y.; Watanabe, T.; Watanabe, T.; Kubo, S. Purification and Characterization of Cell Growth Factor in Bovine Colostrum. J. Vet. Med. Sci. 1996, 58, 47–53. [Google Scholar] [CrossRef]
- Straussberg, R.; Sirota, L.; Hart, J.; Amir, Y.S.; Djaldetti, M.; Bessler, H. Phagocytosis-Promoting Factor in Human Colostrum. Biol. Neonate 1995, 681, 15–18. [Google Scholar] [CrossRef]
- Hidalgo, A.; Chilvers, E.R.; Summers, C.; Koenderman, L. The Neutrophil Life Cycle. Trends Immunol. 2019, 40, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Nonnecke, B.J.; Foote, M.R.; Smith, J.M.; Pesch, B.A.; Van Amburgh, M.E. Composition and Functional Capacity of Blood Mononuclear Leukocyte Populations from Neonatal Calves on Standard and Intensified Milk Replacer Diets. J. Dairy Sci. 2003, 86, 3592–3604. [Google Scholar] [CrossRef]
- Maciag, S.S.; Bellaver, F.V.; Bombassaro, G.; Haach, V.; Morés, M.A.Z.; Baron, L.F.; Coldebella, A.; Bastos, A.P. On the Influence of the Source of Porcine Colostrum in the Development of Early Immune Ontogeny in Piglets. Sci. Rep. 2022, 12, 15630. [Google Scholar] [CrossRef] [PubMed]
- Grigaleviciute, R.; Planciuniene, R.; Prikockyte, I.; Radzeviciute-Valciuke, E.; Baleviciute, A.; Zelvys, A.; Zinkeviciene, A.; Zigmantaite, V.; Kucinskas, A.; Matusevicius, P.; et al. The Influence of Feeding with Colostrum and Colostrum Replacer on Major Blood Biomarkers and Growth Performance in Dairy Calves. Vet. Sci. 2023, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- O’Garra, A. Cytokines Induce the Development of Functionally Heterogeneous T Helper Cell Subsets. Immunity 1998, 8, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Biologic Basis for Interleukin-1 in Disease. J. Am. Soc. Hematol. 1996, 87, 2095–2147. [Google Scholar] [CrossRef]
- Ferrick, D.A.; Schrenzel, M.D.; Mulvania, T.; Hsieh, B.; Ferlin, W.G.; Lepper, H. Differential Production of Interferon-γ and Interleukin-4 in Response to Th1- and Th2-Stimulating Pathogens by Γδ T Cells in Vivo. Nature 1995, 373, 255–257. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic Self-Tolerance Maintained by Activated T Cells Expressing IL-2 Receptor Alpha-Chains (CD25). Breakdown of a Single Mechanism of Self-Tolerance Causes Various Autoimmune Diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Thornton, A.M.; Shevach, E.M. CD4+CD25+ Immunoregulatory T Cells Suppress Polyclonal T Cell Activation in vitro by Inhibiting Interleukin 2 Production. J. Exp. 1998, 188, 287–296. [Google Scholar] [CrossRef]
- Quigley, J.D.; Strohbehn, R.E.; Kost, C.J.; O’Brien, M.M. Formulation of Colostrum Supplements, Colostrum Replacers and Acquisition of Passive Immunity in Neonatal Calves. J. Dairy Sci. 2001, 84, 2059–2065. [Google Scholar] [CrossRef]
- Robison, J.D.; Stott, G.H.; DeNise, S.K. Effects of Passive Immunity on Growth and Survival in the Dairy Heifer. J. Dairy Sci. 1988, 71, 1283–1287. [Google Scholar] [CrossRef]
- Donovan, G.A.; Dohoo, I.R.; Montgomery, D.M.; Bennett, F.L. Associations between Passive Immunity and Morbidity and Mortality in Dairy Heifers in Florida, USA. Prev. Vet. Med. 1998, 34, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Priestley, D.; Bittar, J.H.; Ibarbia, L.; Risco, C.A.; Galvão, K.N. Effect of Feeding Maternal Colostrum or Plasma-Derived or Colostrum-Derived Colostrum Replacer on Passive Transfer of Immunity, Health, and Performance of Preweaning Heifer Calves. J. Dairy Sci. 2013, 96, 3247–3256. [Google Scholar] [CrossRef] [PubMed]
- Misaka, M.; Uematsu, M.; Hashimoto, K.; Kitahara, G.; Osawa, T.; Sasaki, Y. Impact of Dystocia and Cow/Calf Characteristics on Mortality from 0 to 120 Days of Age in Japanese Black Calves in Commercial Cow-Calf Operations. Prev. Vet. Med. 2022, 207, 105716. [Google Scholar] [CrossRef] [PubMed]
- Uetake, K. Newborn Calf Welfare: A Review Focusing on Mortality Rates. Anim. Sci. J. 2013, 84, 101–105. [Google Scholar] [CrossRef]
- Hordofa, D.; Abunna, F.; Megersa, B.; Abebe, R. Incidence of Morbidity and Mortality in Calves from Birth to Six Months of Age and Associated Risk Factors on Dairy Farms in Hawassa City, Southern Ethiopia. Heliyon 2021, 7, e08546. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequences | Accession Number | Length |
|---|---|---|---|---|
| (Base Pairs) | ||||
| GAPDH | F | GTTCAACGGCACAGTCAAGGCAGAG | NM_001034034 | 123 |
| R | ACCACATACTCAGCACCAGCATCAC | |||
| IL-1 β | F | GCCTACGCACATGTCTTCCA | NM_174093 | 111 |
| R | TGCGTCACACAGAAACTCGTC | |||
| IL-2 | F | TGCTGGATTTACAGTTGCTT | XM_024976996 | 111 |
| R | TCAATTCTGTAGCGTTAACCT | |||
| IL-4 | F | ATCAAAACGCTGAACATCCTC | NM_173921 | 142 |
| R | TCCTGTAGATACGCCTAAGCTC | |||
| IL-6 | F | AGCTCTCATTAAGCGCATGG | NM_173923 | 168 |
| R | ATCGCCTGATTGAACCCAG | |||
| IL-10 | F | GGCCTGACATCAAGGAGCAC | NM_174088 | 103 |
| R | CTCTTGTTTTCGCAGGGCAGA | |||
| IFN-γ | F | TGATTCAAATTCCGGTGGAT | NM_174086 | 108 |
| R | TCTTCCGCTTTCTGAGGTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urakawa, M.; Baakhtari, M.; Ramah, A.; Imatake, S.; Ahmadi, P.; Deguchi, Y.; Uematsu, M.; Nakama, Y.; Imabeppu, K.; Nomura, Y.; et al. Comparative Analysis of Maternal Colostrum and Colostrum Replacer Effects on Immunity, Growth, and Health of Japanese Black Calves. Animals 2024, 14, 346. https://doi.org/10.3390/ani14020346
Urakawa M, Baakhtari M, Ramah A, Imatake S, Ahmadi P, Deguchi Y, Uematsu M, Nakama Y, Imabeppu K, Nomura Y, et al. Comparative Analysis of Maternal Colostrum and Colostrum Replacer Effects on Immunity, Growth, and Health of Japanese Black Calves. Animals. 2024; 14(2):346. https://doi.org/10.3390/ani14020346
Chicago/Turabian StyleUrakawa, Marimu, Mahmoud Baakhtari, Amany Ramah, Shoichiro Imatake, Parnian Ahmadi, Yuichiro Deguchi, Mizuho Uematsu, Yoshiki Nakama, Kazunari Imabeppu, Yusuke Nomura, and et al. 2024. "Comparative Analysis of Maternal Colostrum and Colostrum Replacer Effects on Immunity, Growth, and Health of Japanese Black Calves" Animals 14, no. 2: 346. https://doi.org/10.3390/ani14020346
APA StyleUrakawa, M., Baakhtari, M., Ramah, A., Imatake, S., Ahmadi, P., Deguchi, Y., Uematsu, M., Nakama, Y., Imabeppu, K., Nomura, Y., & Yasuda, M. (2024). Comparative Analysis of Maternal Colostrum and Colostrum Replacer Effects on Immunity, Growth, and Health of Japanese Black Calves. Animals, 14(2), 346. https://doi.org/10.3390/ani14020346