
Small Island Effects on the Thermal Biology of the Endemic Mediterranean Lizard Podarcis gaigeae
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
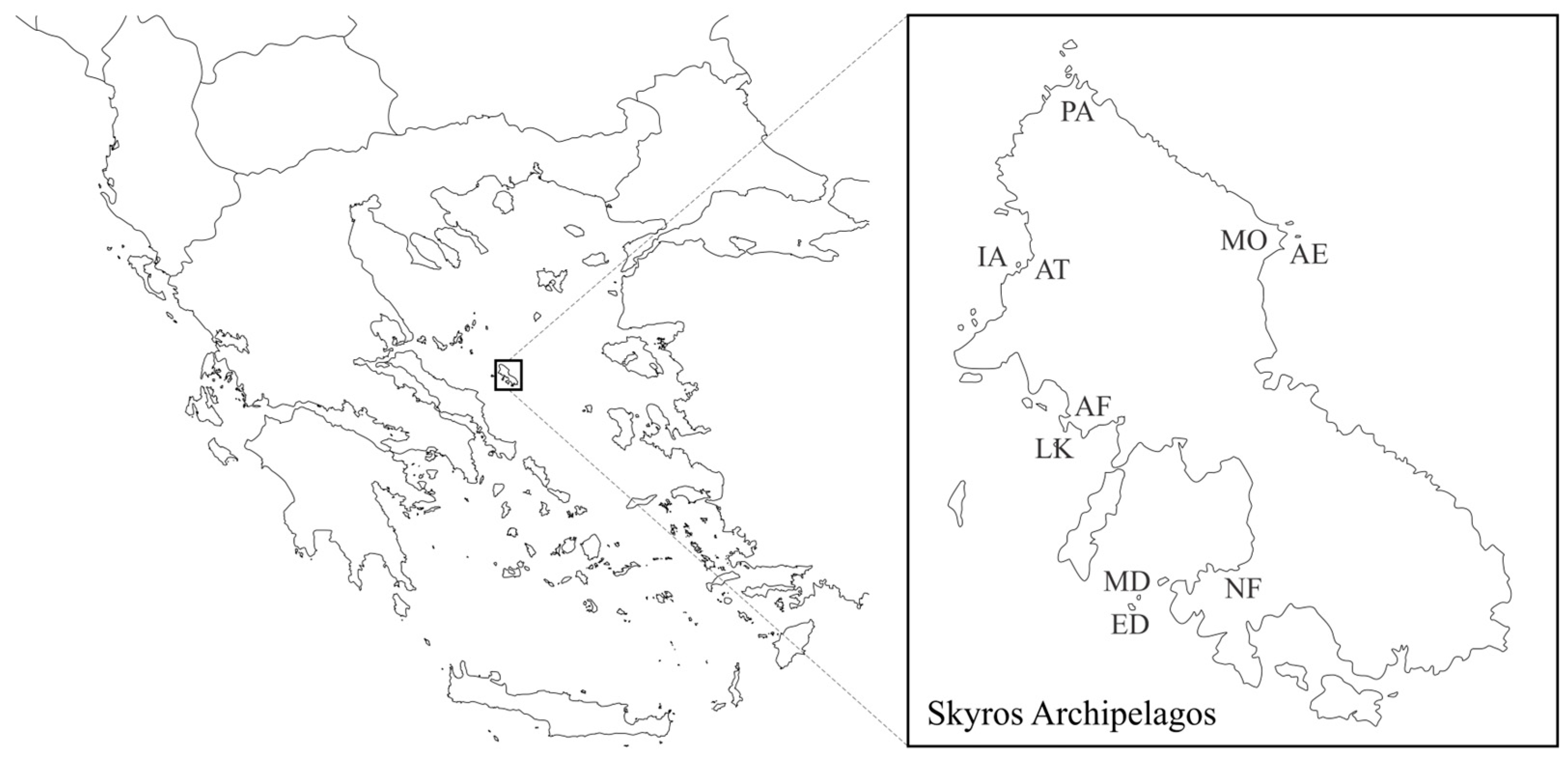
2.1. Study System
2.2. Thermal Measurements
2.2.1. Field Temperatures (Body and Operative Temperatures; Tb and Te)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | SVL (mm) | Mass (g) | Tb (°C) | Tpref (°C) | Te (°C) | db (°C) | de (°C) | EH | EB | |
|---|---|---|---|---|---|---|---|---|---|---|
| Skyros Island (“mainland”) | Agios Fokas (AF) | 6.30 ± 0.50 (5.40–7.30) N = 27 | 5.52 ± 1.63 (3.31–8.95) N = 27 | 31.39 ± 2.98 (25.40–38.00) N = 43 | 32.56 ± 1.35 (29.07–35.42) N = 27 | 39.11 ± 8.86 (23.90–70.80) N = 28 | 1.17 ± 1.41 (0.0–5.57) N = 43 | 5.87 ± 7.11 (0.0–35.51) N = 28 | 0.80 | 4.70 |
| Atsitsa (AT) | 4.54 ± 1.67 (2.10–8.10) N = 31 | 5.57 ± 0.99 (2.65–7.10) N = 31 | 30.24 ± 3.72 (21.30–35.90) N = 48 | 31.31 ± 1.74 (28.15–35.13) N = 31 | 34.92 ± 8.35 (19.40–62.90) N = 28 | 1.65 ± 2.21 (0.0–8.47) N = 48 | 5.07 ± 6.40 (0.0–29.77) N = 28 | 0.67 | 3.42 | |
| Molos (MO) | 5.70 ± 0.42 (4.50–6.50) N = 25 | 4.81 ± 1.32 (2.35–7.77) N = 25 | 33.06 ± 2.48 (26.80–39.10) N = 60 | 32.97 ± 0.90 (31.37–34.80) N = 25 | 41.48 ± 8.65 (22.40–73.30) N = 28 | 0.95 ± 1.29 (0.0–5.20) N = 60 | 8.00 ± 7.59 (0.0–38.83) N = 28 | 0.88 | 7.05 | |
| Nyfi (NF) | 6.19 ± 0.53 (5.30–7.10) N = 23 | 5.98 ± 1.64 (3.93–9.36) N = 23 | 32.03 ± 3.22 (24.90–37.80) N = 52 | 32.23 ± 1.96 (28.82–35.52) N = 23 | 38.79 ± 8.23 (22.30–66.80) N = 28 | 0.98 ± 1.31 (0.0–5.21) N = 52 | 6.53 ± 6.69 (0.0–32.34) N = 28 | 0.85 | 5.56 | |
| Palamari (PA) | 5.54 ± 0.54 (4.70–6.70) N = 18 | 4.00 ± 1.42 (2.57–7.70) N = 18 | 33.20 ± 2.69 (27.20–37.40) N = 57 | 33.27 ± 1.28 (30.77–35.37) N = 18 | 42.06 ± 9.11 (25.40–72.30) N = 28 | 0.66 ± 1.12 (0.0–4.34) N = 57 | 7.29 ± 8.19 (0.0–36.60) N = 28 | 0.91 | 6.63 | |
| Islet | Agios Emolaos (AE) | 5.79 ± 0.64 (4.50–6.80) N = 29 | 4.58 ± 1.58 (2.60–7.49) N = 29 | 32.73 ± 2.57 (28.70–37.70) N = 39 | 33.54 ± 0.85 (31.93–35.40) N = 29 | 40.29 ± 8.83 (26.30–69.50) N = 28 | 1.29 ± 1.38 (0.0–4.03) N = 39 | 6.44 ± 7.55 (0.0–34.14) N = 28 | 0.80 | 5.16 |
| Atsitsa Islet (AI) | 6.42 ± 0.52 (5.10–7.40) N = 28 | 6.03 ± 1.73 (3.24–9.73) N = 28 | 30.39 ± 1.85 (28.00–35.20) N = 38 | 32.42 ± 0.63 (31.30–33.65) N = 28 | 37.66 ± 9.77 (24.90–71.20) N = 28 | 1.40 ± 1.08 (0.0–3.30) N = 38 | 4.61 ± 9.11 (0.0–36.99) N = 28 | 0.70 | 3.21 | |
| Exo Diavates (ED) | 7.67 ± 0.93 (6.30–8.80) N = 28 | 12.15 ± 5.01 (6.60–19.51) N = 28 | 32.05 ± 2.87 (25.00–37.60) N = 49 | 34.07 ± 0.94 (32.08–35.20) N = 28 | 40.03 ± 8.53 (24.80–72.60) N = 28 | 1.84 ± 2.32 (0.0–8.23) N = 49 | 5.87 ± 7.89 (0.0–37.60) N = 28 | 0.69 | 4.03 | |
| Lakonisi (LK) | 6.93 ± 0.81 (5.50–8.10) N = 25 | 9.07 ± 3.45 (3.55–15.24) N = 25 | 32.12 ± 2.39 (27.00–38.80) N = 45 | 33.85 ± 0.83 (32.08–35.20) N = 25 | 37.84 ± 8.62 (25.20–70.90) N = 28 | 1.54 ± 1.60 (0.0–5.99) N = 45 | 5.29 ± 6.60 (0.0–35.30) N = 28 | 0.71 | 3.75 | |
| Mesa Diavates (MD) | 5.97 ± 0.65 (4.90–7.00) N = 26 | 4.90 ± 1.75 (2.54–8.04) N = 26 | 32.78 ± 2.79 (28.10–38.60) N = 42 | 33.70 ± 0.95 (31.73–35.27) N = 26 | 38.85 ± 10.62 (25.70–66.70) N = 28 | 1.49 ± 1.50 (0.0–4.64) N = 42 | 6.88 ± 8.55 (0.0–31.66) N = 28 | 0.78 | 5.39 |
2.2.2. Lab Measurements (Preferred and Set-Point Temperatures; Tpref and Tset)
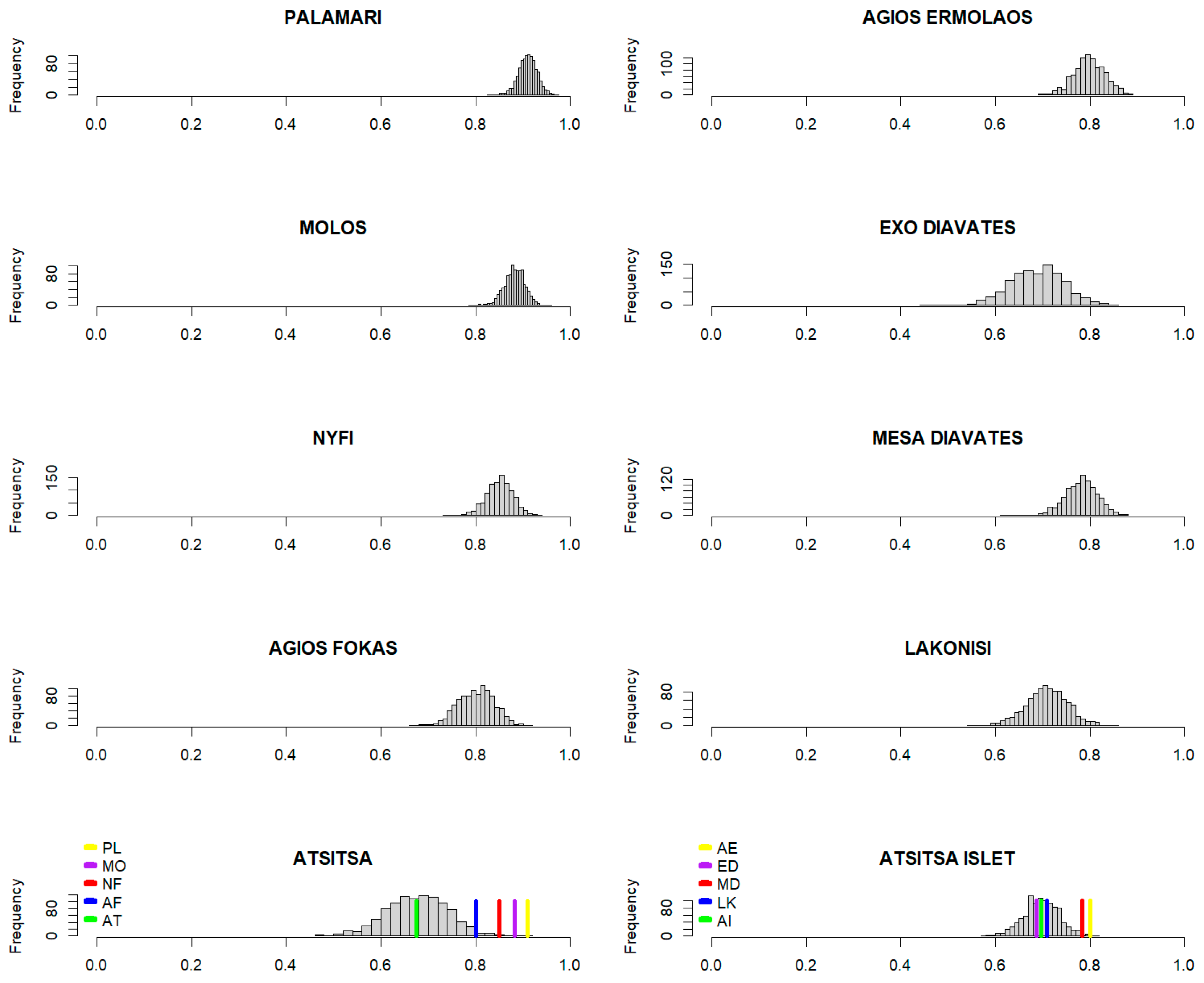
2.2.3. Effectiveness of Thermoregulation (E)
2.3. Statistical Analysis
3. Results
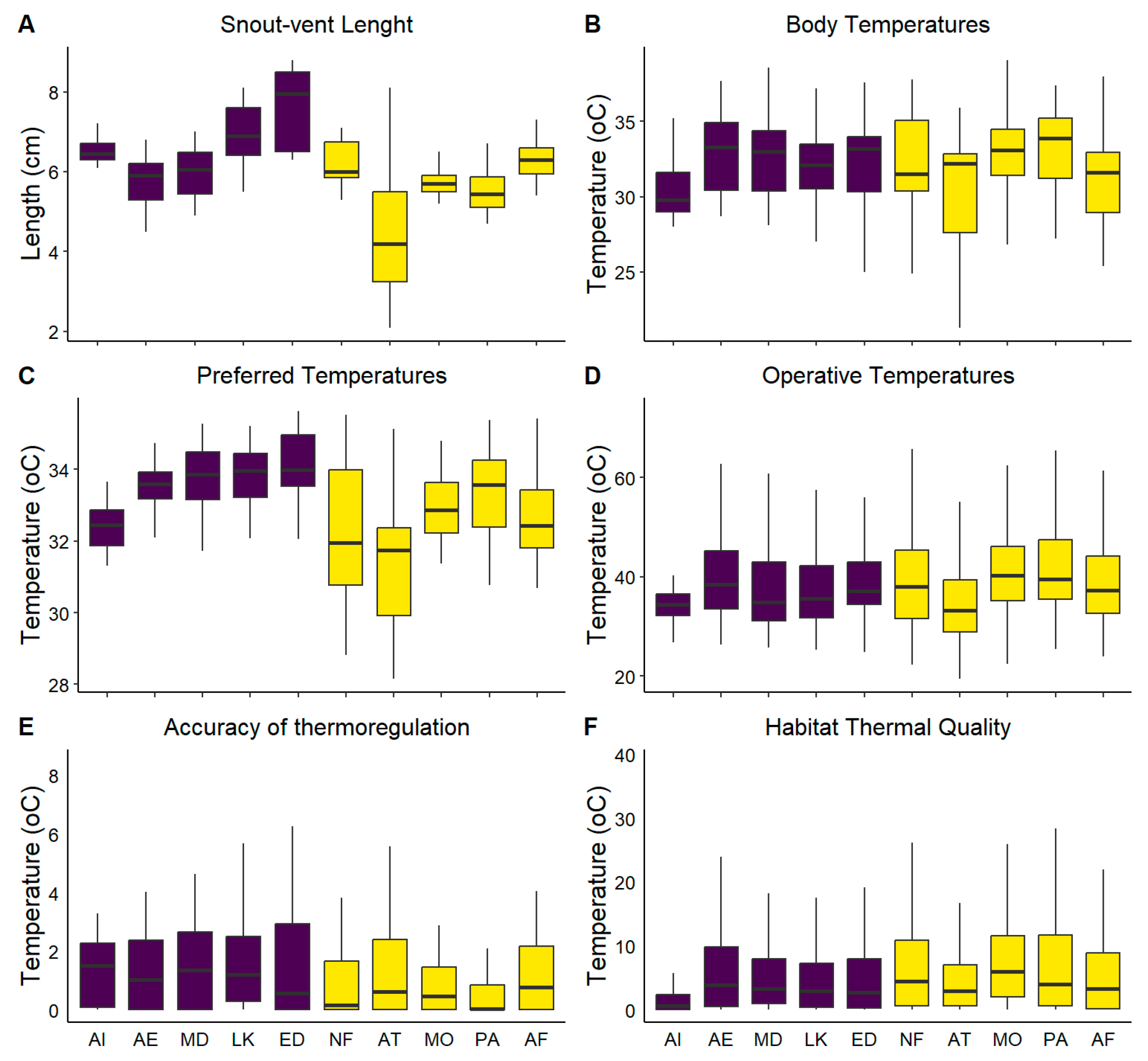
3.1. Body Size and Field Measurements (Tb and Te)
3.2. Lab Measurements (Tpref and Tset)
3.3. Effectiveness of Thermoregulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Novosolov, M.; Raia, P.; Meiri, S. The island syndrome in lizards. Glob. Ecol. Biogeogr. 2013, 22, 184–191. [Google Scholar] [CrossRef]
- Benítez-López, A.; Santini, L.; Gallego-Zamorano, J.; Milá, B.; Walkden, P.; Huijbregts, M.A.J.; Tobias, J.A. The island rule explains consistent patterns of body size evolution in terrestrial vertebrates. Nat. Ecol. Evol. 2021, 5, 768–786. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution and Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Vervust, B.; Pafilis, P.; Valakos, E.D.; Van Damme, R. Anatomical and physiological changes associated with a recent dietary shift in the lizard Podarcis sicula. Physiol. Biochem. Zool. 2010, 83, 632–642. [Google Scholar] [CrossRef]
- Rozzi, R.; Varela, S.; Bover, P.; Martin, J.M. Causal explanations for the evolution of ‘low gear’ locomotion in insular ruminants. J. Biogeogr. 2020, 47, 2274–2285. [Google Scholar] [CrossRef]
- Sagonas, K.; Pafilis, P.; Lymberakis, P.; Donihue, C.M.; Herrel, A.; Valakos, E.D. Insularity affects head morphology, bite force and diet in a Mediterranean lizard. Biol. J. Linn. Soc. 2014, 112, 469–484. [Google Scholar] [CrossRef]
- Schwaner, T.D. A field study of thermoregulation in black tiger snakes (Notechis ater niger: Elapidae) on the Franklin Islands, South Australia. Herpetologica 1989, 45, 393–401. [Google Scholar]
- Gaudenti, N.; Nix, E.; Maier, P.; Westphal, M.F.; Taylor, E.N. Habitat heterogeneity affects the thermal ecology of an endangered lizard. Ecol. Evol. 2021, 11, 14843–14856. [Google Scholar] [CrossRef]
- Sagonas, K.; Valakos, E.D.; Pafilis, P. The impact of insularity on the thermoregulation of a Mediterranean lizard. J. Therm. Biol. 2013, 38, 480–486. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Herpetology: An Introductory Biology of Amphibians and Reptiles, 4th ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Meiri, S.; Bauer, A.M.; Chirio, L.; Colli, G.R.; Das, I.; Doan, T.M.; Feldman, A.; Herrera, F.-C.; Novosolov, M.; Pafilis, P.; et al. Are lizards feeling the heat? A tale of ecology and evolution under two temperatures. Glob. Ecol. Biogeogr. 2013, 22, 834–845. [Google Scholar] [CrossRef]
- Aragón, P.; Lobo, J.M.; Olalla-Tárraga, M.Á.; Rodríguez, M.Á. The contribution of contemporary climate to ectothermic and endothermic vertebrate distributions in a glacial refuge. Glob. Ecol. Biogeogr. 2010, 19, 40–49. [Google Scholar] [CrossRef]
- Sinervo, B.; Méndez-de-la-Cruz, F.; Miles, D.B.; Heulin, B.; Bastiaans, E.; Villagrán-Santa Cruz, M.; Lara-Resendiz, R.; Martínez-Méndez, N.; Calderón-Espinosa, M.L.; Meza-Lázaro, R.N.; et al. Erosion of Lizard Diversity by Climate Change and Altered Thermal Niches. Science 2010, 328, 894–899. [Google Scholar] [CrossRef] [PubMed]
- De Chazal, J.; Rounsevell, M.D.A. Land-use and climate change within assessments of biodiversity change: A review. Glob. Environ. Chang. 2009, 19, 306–315. [Google Scholar] [CrossRef]
- Mi, C.; Ma, L.; Wang, Y.; Wu, D.; Du, W.; Sun, B. Temperate and tropical lizards are vulnerable to climate warming due to increased water loss and heat stress. Proc. R. Soc. Lond. B 2022, 289, 20221074. [Google Scholar] [CrossRef] [PubMed]
- Pontes-da-Silva, E.; Magnusson, W.E.; Sinervo, B.; Caetano, G.H.; Miles, D.B.; Colli, G.R.; Diele-Viegas, L.M.; Fenker, J.; Santos, J.C.; Werneck, F.P. Extinction risks forced by climatic change and intraspecific variation in the thermal physiology of a tropical lizard. J. Therm. Biol. 2018, 73, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Angilletta, M.J.J. Thermal Adaptation. A Theoritical and Empirical Synthesis; Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- Logan, M.L.; Cox, R.M.; Calsbeek, R. Natural selection on thermal performance in a novel thermal environment. Proc. Natl. Acad. Sci. USA 2014, 111, 14165–14169. [Google Scholar] [CrossRef]
- Hertz, P.E.; Huey, R.B.; Stevenson, R.D. Evaluating temperature regulation by field-active ectotherms: The fallacy of the inappropriate question. Am. Nat. 1993, 142, 796–818. [Google Scholar] [CrossRef]
- Pafilis, P.; Foufopoulos, J.; Sagonas, K.; Runemark, A.; Svensson, E.; Valakos, E.D. Reproductive biology of insular reptiles: Marine subsidies modulate expression of the “Island Syndrome”. Copeia 2011, 2011, 545–552. [Google Scholar] [CrossRef]
- Runemark, A.; Gabirot, M.; Svensson, E.I. Population divergence in chemical signals and the potential for premating isolation between islet- and mainland populations of the Skyros wall lizard (Podarcis gaigeae). J. Evol. Biol. 2011, 24, 795–809. [Google Scholar] [CrossRef]
- Sagonas, K.; Rota, I.A.; Tsitsilonis, O.E.; Pafilis, P.; Valakos, E.D. Infection risk dictates immunological divergence among populations in a Mediterranean lizard. J. Evol. Biol. 2016, 29, 1680–1688. [Google Scholar] [CrossRef]
- Shmida, A.; Wilson, M.V. Biological Determinants of Species Diversity. J. Biogeogr. 1985, 12, 1–20. [Google Scholar] [CrossRef]
- Garland, J.T.; Adolph, S.C. Why not to do two-species comparative studies: Limitations on inferring adaptation. Physiol. Zool. 1994, 67, 797–828. [Google Scholar] [CrossRef]
- Waples, R.S.; Gaggiotti, O. INVITED REVIEW: What is a population? An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity. Mol. Ecol. 2006, 15, 1419–1439. [Google Scholar] [CrossRef] [PubMed]
- Pafilis, P.; Lymberakis, P.; Sagonas, K.; Valakos, E. The particularities of a remote islet shape the thermoregulatory profile of an endemic Mediterranean lizard. J. Therm. Biol. 2016, 61, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Triantis, K.A.; Vardinoyannis, K.; Tsolaki, E.P.; Botsaris, I.; Lika, K.; Mylonas, M. Re-approaching the small island effect. J. Biogeogr. 2006, 33, 914–923. [Google Scholar] [CrossRef]
- Sagonas, K.; Meiri, S.; Valakos, E.D.; Pafilis, P. The effect of body size on the thermoregulation of lizards on hot, dry Mediterranean islands. J. Therm. Biol. 2013, 38, 92–97. [Google Scholar] [CrossRef]
- Valakos, E.D.; Pafilis, P.; Sotiropoulos, K.; Lymberakis, P.; Maragou, P.; Foufopoulos, J. The Amphibians and Reptiles of Greece; Chimaira: Frankfurt am Main, Germany, 2008. [Google Scholar]
- Runemark, A.; Hey, J.; Hansson, B.; Svensson, E.I. Vicariance divergence and gene flow among islet populations of an endemic lizard. Mol. Ecol. 2012, 21, 117–129. [Google Scholar] [CrossRef]
- Pafilis, P.; Foufopoulos, J.; Poulakakis, N.; Lymberakis, P.; Valakos, E.D. Tail shedding in island lizards [Lacertidae, Reptilia]: Decline of antipredator defenses in relaxed predation environments. Evolution 2009, 63, 1262–1278. [Google Scholar] [CrossRef]
- Pafilis, P.; Herrel, A.; Kapsalas, G.; Vasilopoulou-Kampitsi, M.; Fabre, A.-C.; Foufopoulos, J.; Donihue, C.M. Habitat shapes the thermoregulation of Mediterranean lizards introduced to replicate experimental islets. J. Therm. Biol. 2019, 84, 368–374. [Google Scholar] [CrossRef]
- Veríssimo, C.V.; Carretero, M.A. Preferred temperatures of Podarcis vaucheri from Morocco: Intraspecific variation and interspecific comparisons. Amphibia-Reptilia 2009, 30, 17–23. [Google Scholar] [CrossRef]
- Bakken, G.S. Measurement and application of operative and standard operative temperatures in ecology. Am. Zool. 1992, 32, 194–216. [Google Scholar] [CrossRef]
- Díaz, J.A. Ecological correlates of the thermal quality of an ectotherm’s habitat: A comparison between two temperate lizard populations. Funct. Ecol. 1997, 18, 867–875. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Reinert, H.K. Modeling body temperature and thermal inertia of large-bodied reptiles: Support for water-filled biophysical models in radiotelemetric studies. J. Therm. Biol. 2012, 37, 282–285. [Google Scholar] [CrossRef]
- Dzialowski, E.M. Use of operative temperature and standard operative temperature models in thermal biology. J. Therm. Biol. 2005, 30, 317–334. [Google Scholar] [CrossRef]
- Huey, R. Physiological consequences of habitat selection. Am. Nat. 1991, 137, S91–S115. [Google Scholar] [CrossRef]
- Blouin-Demers, G.; Weatherhead, P.J. Thermal ecology of black rat snakes (Elaphe obsoleta) in a thermally challenging environment. Ecology 2001, 82, 3025–3043. [Google Scholar] [CrossRef]
- Labocha, M.K.; Schutz, H.; Hayes, J.P. Which body condition index is best? Oikos 2014, 123, 111–119. [Google Scholar] [CrossRef]
- Van Damme, R.; Bauwens, D.; Verheyen, R.F. Selected body temperatures in the lizard Lacerta vivipara: Variation within and between populations. J. Therm. Biol. 1986, 11, 219–222. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Maia-Carneiro, T.; Dorigo, T.A.; Rocha, C.F.D. Influences of Seasonality, Thermal Environment and Wind Intensity on the Thermal Ecology of Brazilian Sand Lizards In A Restinga Remnant. South Am. J. Herpetol. 2012, 211, 241–251. [Google Scholar] [CrossRef]
- Ortega, Z.; Pérez-Mellado, V.; Garrido, M.; Guerra, C.; Villa-García, A.; Alonso-Fernández, T. Seasonal changes in thermal biology of Podarcis lilfordi (Squamata, Lacertidae) consistently depend on habitat traits. J. Therm. Biol. 2014, 39, 32–39. [Google Scholar] [CrossRef]
- Vrahnakis, M.; Fotiadis, G.; Kazoglou, Y. Skyros: A very “dry grassland” Greek island. Bull. 14—Eurasian Dry Grassl. Group 2012, 14, 9–10. [Google Scholar]
- Pafilis, P.; Maragou, P.; Sagonas, K.; Valakos, E. Partitioning thermal habitat on a vertical rock, a herculean task. J. Therm. Biol. 2017, 70, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Sears, M.W.; Angilletta, M.J.J. Costs and benefits of thermoregulation revisited: Both the heterogeneity and spatial structure of temperature drive energetic costs. Am. Nat. 2015, 185, E94–E102. [Google Scholar] [CrossRef] [PubMed]
- Žagar, A.; Carretero, M.A.; Osojnik, N.; Sillero, N.; Vrezec, A. A place in the sun: Interspecific interference affects thermoregulation in coexisting lizards. Behav. Ecol. Sociobiol. 2015, 69, 1127–1137. [Google Scholar] [CrossRef]
- Bauwens, D.; Hertz, P.E.; Castilla, A.M. Thermoregulation in a lacertid lizard: The relative contributions of distinct behavioral mechanisms. Ecology 1996, 77, 1818–1830. [Google Scholar] [CrossRef]
- Huey, R.B. Temperature, physiology, and the ecology of reptiles. In Biology of the Reptilia; Gans, C., Pough, F.H., Eds.; Academic Press: New York, NY, USA, 1982; Volume 12, pp. 25–91. [Google Scholar]
- Runemark, A.; Sagonas, K.; Svensson, E.I. Ecological explanations to island gigantism: Dietary niche divergence, predation, and size in an endemic lizard. Ecology 2015, 96, 2077–2092. [Google Scholar] [CrossRef] [PubMed]
- Wilms, T.M.; Wagner, P.; Shobrak, M.; Rödder, D.; Böhme, W. Living on the edge?—On the thermobiology and activity pattern of the large herbivorous desert lizard Uromastyx aegyptia microlepis Blanford, 1875 at Mahazat as-Sayd Protected Area, Saudi Arabia. J. Arid Environ. 2011, 75, 636–647. [Google Scholar] [CrossRef]
- Adamopoulou, C.; Valakos, E.D. Thermal ecology and activity cycle of Podarcis milensis in a sandy coastal area. Isr. J. Zool. 2005, 51, 39–52. [Google Scholar] [CrossRef]
- Corbalán, V.; Debandi, G.; Kubisch, E. Thermal ecology of two sympatric saxicolous lizards of the genus Phymaturus from the Payunia region (Argentina). J. Therm. Biol. 2013, 38, 384–389. [Google Scholar] [CrossRef]
- Díaz de la Vega-Pérez, A.H.; Jiménez-Arcos, V.H.; Manríquez-Morán, N.L.; Méndez-de la Cruz, F.R. Conservatism of thermal preferences between parthenogenetic Aspidoscelis cozumela complex (Squamata: Teiidae) and their parental species. J. Herpetol. 2013, 23, 93–104. [Google Scholar]
- Grover, M.C. Microhabitat use and thermal ecology of two narrowly sympatric sceloporus (Phrynosomatidae) lizards. J. Herpetol. 1996, 30, 152–160. [Google Scholar] [CrossRef]
- Hertz, P.E.; Huey, R.B.; Nevo, E. Homage to Santa Anita: Thermal sensitivity of sprint speed in agamid lizards. Evolution 1983, 37, 1075–1084. [Google Scholar] [CrossRef]
- Crowley, S. Thermal sensitivity of sprint-running in the lizard Sceloporus undulatus: Support for a conservative view of thermal physiology. Oecologia 1985, 66, 219–225. [Google Scholar] [CrossRef]
- Truter, J.C.; van Wyk, J.H.; Mouton, P.l.F.N. An evaluation of daily, seasonal and population-level variation in the thermal preference of a group-living lizard, Ouroborus cataphractus (Sauria: Cordylidae). Amphibia-Reptilia 2014, 35, 391–403. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Rubiño-Hispán, M.V.; Reguera, S.; Moreno-Rueda, G. Thermal dependence of sprint performance in the lizard Psammodromus algirus along a 2200-meter elevational gradient: Cold-habitat lizards do not perform better at low temperatures. J. Therm. Biol. 2015, 52, 90–96. [Google Scholar] [CrossRef]
- Angilletta, M.J.J. Thermal and physiological constraints on energy assimilation in a widespread lizard (Sceloporus undulatus). Ecology 2001, 82, 3044–3056. [Google Scholar] [CrossRef]
- McConnachie, S.; Alexander, G.J. The effect of temperature on digestive and assimilation efficiency, gut passage time and appetite in an ambush foraging lizard, Cordylus melanotus melanotus. J. Comp. Physiol. B 2004, 174, 99–105. [Google Scholar] [CrossRef]
- Shine, R.; Harlow, P. Maternal thermoregulation influences offspring viability in a viviparous lizard. Oecologia 1993, 96, 122–127. [Google Scholar] [CrossRef]
- Zhang, Y.-P.; Ji, X. The thermal dependence of food assimilation and locomotor performance in southern grass lizards, Takydromus sexlineatus (Lacertidae). J. Therm. Biol. 2004, 29, 45–53. [Google Scholar] [CrossRef]
- Scheers, H.; Van Damme, R. Micro-scale differences in thermal habitat quality and a possible case of evolutionary flexibility in the thermal physiology of lacertid lizards. Oecologia 2002, 132, 323–331. [Google Scholar] [CrossRef]
- Van Damme, R.; Bauwens, D.; Verheyen, R.F. The thermal dependence of feeding behaviour, food consumption and gut-passage time in the lizard Lacerta vivipara Jacquin. Funct. Ecol. 1991, 5, 507–517. [Google Scholar] [CrossRef]
- Marquet, P.A.; Ortíz, J.C.; Božinović, F.; Jaksié, F.M. Ecological aspects of thermoregulation at high altitudes: The case of andean Liolaemus lizards in northern Chile. Oecologia 1989, 81, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Case, T.J. Ecology and evolution of the insular giant chuckawallas, Sauromalus hispidus and Sauromalus varius. In Iguanas of the World: Their Behavior, Ecology and Conservation; Burghardt, G.M., Rand, A.S., Eds.; Noyes Publications: Park Ridge, NJ, USA, 1982; pp. 184–212. [Google Scholar]
- Huey, R.B.; Slatkin, M. Cost and benefits of lizard thermoregulation. Q. Rev. Biol. 1976, 51, 363–384. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reppa, A.; Agori, A.F.; Santikou, P.; Parmakelis, A.; Pafilis, P.; Valakos, E.D.; Sagonas, K. Small Island Effects on the Thermal Biology of the Endemic Mediterranean Lizard Podarcis gaigeae. Animals 2023, 13, 2965. https://doi.org/10.3390/ani13182965
Reppa A, Agori AF, Santikou P, Parmakelis A, Pafilis P, Valakos ED, Sagonas K. Small Island Effects on the Thermal Biology of the Endemic Mediterranean Lizard Podarcis gaigeae. Animals. 2023; 13(18):2965. https://doi.org/10.3390/ani13182965
Chicago/Turabian StyleReppa, Aikaterini, Ariadne Faidra Agori, Panayiota Santikou, Aristeidis Parmakelis, Panayiotis Pafilis, Efstratios D. Valakos, and Kostas Sagonas. 2023. "Small Island Effects on the Thermal Biology of the Endemic Mediterranean Lizard Podarcis gaigeae" Animals 13, no. 18: 2965. https://doi.org/10.3390/ani13182965