
Transcriptomics in Rare Minnow (Gobiocypris rarus) towards Attenuated and Virulent Grass Carp Reovirus Genotype II Infection
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Virus
2.3. Virus Infection and Sample Collecting
2.4. RNA Extraction, cDNA Library Construction, and Sequencing
2.5. De Novo Assembly and Annotation
- (1)
- removing reads containing adapters;
- (2)
- removing reads containing more than 10% of unknown nucleotides (N);
- (3)
- removing low-quality reads containing more than 50% of low-quality (Q-value ≤ 20) bases.
2.6. Gene Expression Analysis and Enrichment Analysis
2.7. Validation of DEGs by RT-qPCR
3. Results
3.1. Transcriptome Sequencing, De Novo Assembly, and Annotation
3.2. Identification and Enrichment of Differentially Expressed Genes
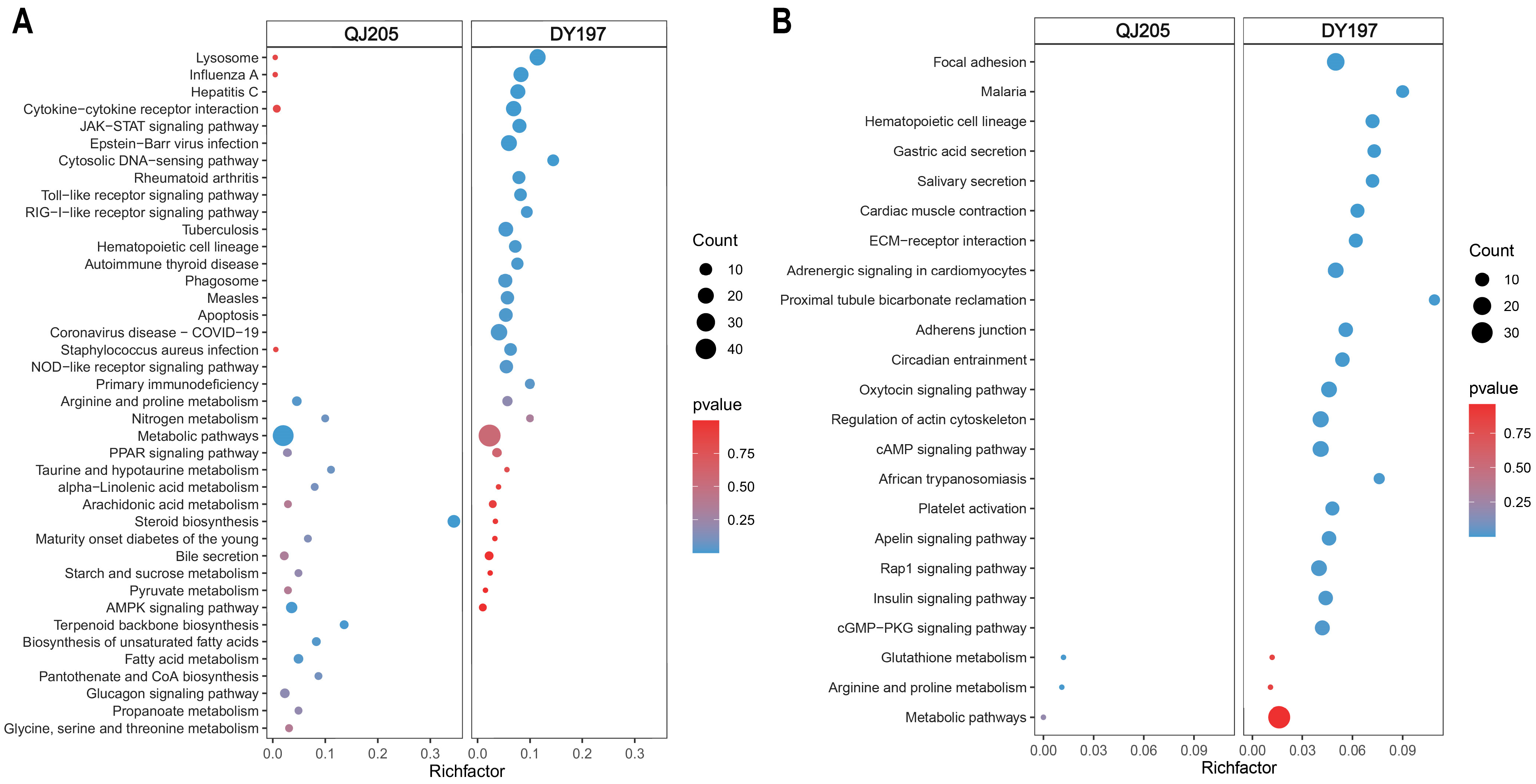
3.3. Enrichment Analysis of Differentially Expressed Genes in Spleen
3.4. Enrichment Analysis of Differentially Expressed Genes in Liver
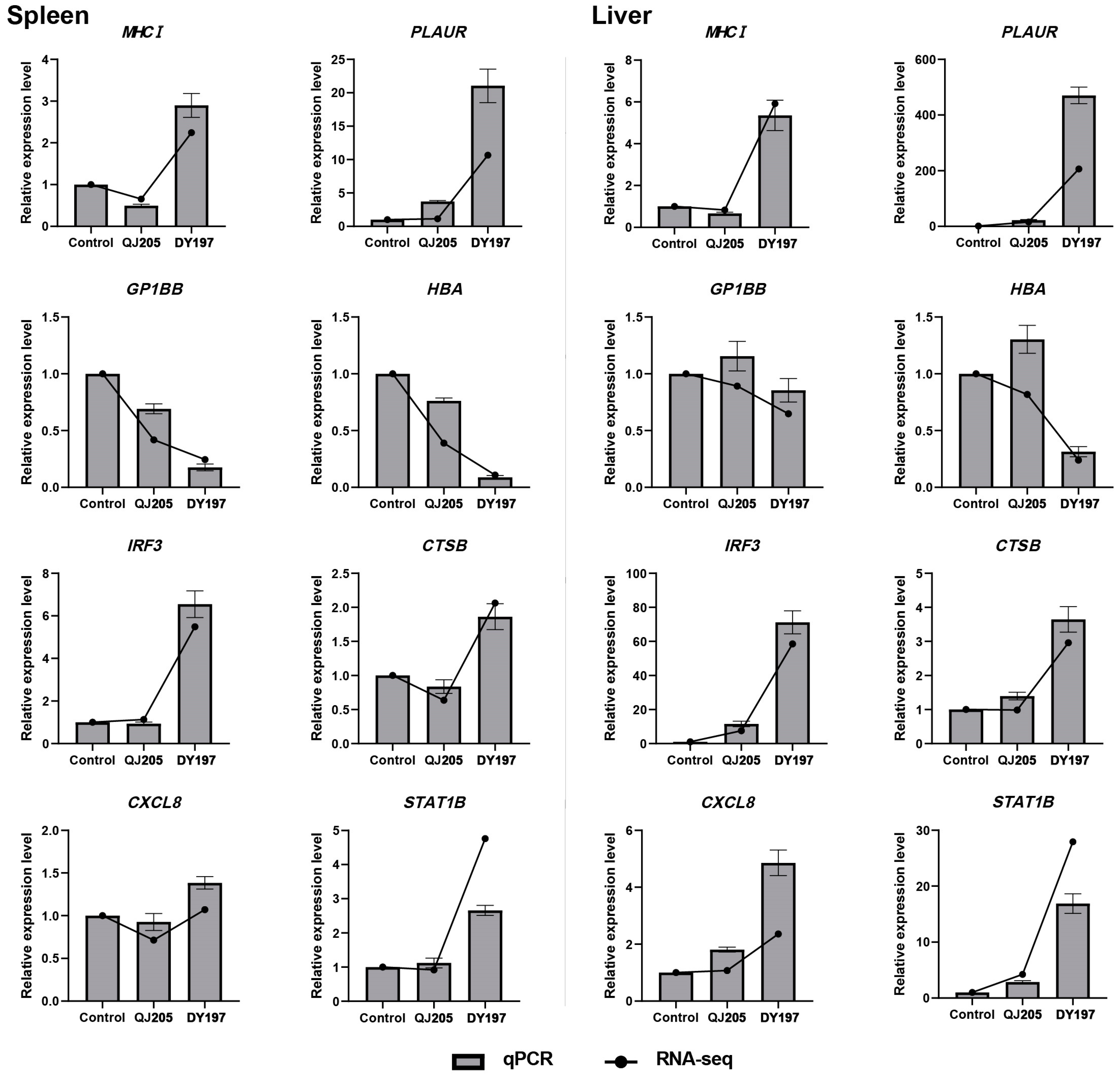
3.5. Validation of Differentially Expressed Genes by qPCR
4. Discussion
4.1. Immune Response
4.2. Apoptosis and Necroptosis-Related Pathways
4.3. Protein Digestion and Absorption
4.4. Cell Proliferation and Migration
4.5. Hemorrhage-Related Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, L.; Fang, Q.; Shah, S.; Atanasov, I.; Zhou, Z. Subnanometer-resolution structures of the grass carp reovirus core and virion. J. Mol. Biol. 2008, 382, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Y.; Su, J. Insights into the antiviral immunity against grass carp (Ctenopharyngodon idella) reovirus (GCRV) in grass carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Zhang, A.; Pei, Y.; Chu, P.; Li, Y.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Differences in responses of grass carp to different types of grass carp reovirus (GCRV) and the mechanism of hemorrhage revealed by transcriptome sequencing. BMC Genom. 2017, 18, 452. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zeng, W.; Liu, C.; Zhang, C.; Wang, Y.; Shi, C.; Wu, S. Complete Genome Sequence of a Reovirus Isolated from Grass Carp, Indicating Different Genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, W.; Li, Y.; Zhou, Y.; Meng, Y.; Zeng, L.; Vakharia, V.N.; Fan, Y. Isolation, Identification, and Genomic Analysis of a Novel Reovirus from Healthy Grass Carp and Its Dynamic Proliferation In Vitro and In Vivo. Viruses 2021, 13, 690. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, J.; Zhou, Y.; Jiang, N.; Liu, W.; Zeng, L. Establishment and application of a universal RT-PCR assay for detection of different genotype grass carp reovirus. Freshw. Fish. 2018, 48, 8. [Google Scholar]
- Zeng, W.; Wang, Q.; Wang, Y.; Zhao, C.; Li, Y.; Shi, C.; Wu, S.; Song, X.; Huang, Q.; Li, S. Immunogenicity of a cell culture-derived inactivated vaccine against a common virulent isolate of grass carp reovirus. Fish Shellfish. Immunol. 2016, 54, 473–480. [Google Scholar] [CrossRef]
- Xing, Y.; Tian, Y.; Deng, G.; Chi, Y.; Jiang, X. Complete genomic sequence of a reovirus isolated from grass carp in China. Virus Res. 2012, 163, 275–283. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, S.; Ji, G.; Liu, J.; Shi, L.; Zhou, B. Research and Discussion of Rare Minnow as an Aquatic Model Organism. Asian J. Ecotoxicol. 2017, 12, 38–46. [Google Scholar]
- Wang, T.; Yi, Y.; Chen, H.; Liu, H.; Wen, G. Histopathological observation on hemorrhagic rare minnow (gobiocypris rarus) infected artificially with hemorrhagic virus of grass carp. Acta Hydrobiol. Sin. 1995, 19, 350–353. [Google Scholar]
- Chen, J.; Chang, O.; Li, Y.; Wang, Y.; Liu, C.; Yin, J.; Bergmann, S.M.; Zeng, W.; Wang, Q. Establishment of a rare minnow (Gobiocypris rarus) disease model for grass carp reovirus genotype II. Aquaculture 2020, 533, 736133. [Google Scholar] [CrossRef]
- Zapata, A.; Díez, B.; Cejalvo, T.; Frías, C.; Cortés, A. Ontogeny of the immune system of fish. Fish Shellfish. Immunol. 2006, 20, 126–136. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Muñoz, P.; López-Muñoz, A.; Pelegrín, P.; Ayala, A.G.; Mulero, V.; Meseguer, J. Early innate immune response and redistribution of inflammatory cells in the bony fish gilthead seabream experimentally infected with Vibrio anguillarum. Cell Tissue Res. 2005, 320, 61–68. [Google Scholar] [CrossRef]
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish. Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef]
- Freitas-Lopes, M.A.; Mafra, K.; David, B.A.; Carvalho-Gontijo, R.; Menezes, G.B. Differential Location and Distribution of Hepatic Immune Cells. Cells 2017, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Zeng, W.; Wang, Y.; Wang, Q.; Wu, S. Pathogenicity and tissue distribution of grass carp reovirus after intraperitoneal administration. Virol. J. 2014, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Fu, X.; Li, N.; Liu, L.; Lin, Q.; Li, Y.; Peng, Y.; Huang, Z.; Wu, S. The distribution of different virulence grass carp reovirus strains in some neglected tissues. Pol. J. Vet. Sci. 2016, 19, 763–770. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zeng, W.; Wang, Y.; Wang, Q.; Yin, J.; Li, Y.; Wang, C.; Bergmann, S.M.; Gao, C.; Hu, H. Comparison of the blood parameters and histopathology between grass carp infected with a virulent and avirulent isolates of genotype Ⅱ grass carp reovirus. Microb. Pathog. 2019, 139, 103859. [Google Scholar] [CrossRef]
- Ye, H.; Lin, Q.; Luo, H. Applications of transcriptomics and proteomics in understanding fish immunity. Fish Shellfish. Immunol. 2018, 77, 319–327. [Google Scholar] [CrossRef]
- Bersanelli, M.; Mosca, E.; Remondini, D.; Giampieri, E.; Sala, C.; Castellani, G.; Milanesi, L. Methods for the integration of multi-omics data: Mathematical aspects. BMC Bioinform. 2016, 17 (Suppl. 2), 15. [Google Scholar] [CrossRef] [Green Version]
- Dettleff, P.; Moen, T.; Santi, N.; Martinez, V. Transcriptomic analysis of spleen infected with infectious salmon anemia virus reveals distinct pattern of viral replication on resistant and susceptible Atlantic salmon (Salmo salar). Fish Shellfish. Immunol. 2017, 61, 187–193. [Google Scholar] [CrossRef]
- Ling, X.; Dong, W.; Zhang, Y.; Qian, X.; Zhang, W.; He, W.; Zhao, X.; Liu, J. Comparative transcriptomics and histopathological analysis of crucian carp infection by atypical Aeromonas salmonicida. Fish Shellfish. Immunol. 2019, 94, 294–307. [Google Scholar] [CrossRef]
- Zhu, J.; Li, C.; Ao, Q.; Tan, Y.; Luo, Y.; Guo, Y.; Lan, G.; Jiang, H.; Gan, X. Trancriptomic profiling revealed the signatures of acute immune response in tilapia (Oreochromis niloticus) following Streptococcus iniae challenge. Fish Shellfish. Immunol. 2015, 46, 346–353. [Google Scholar] [CrossRef]
- Wang, P.; Wang, J.; Su, Y.; Mao, Y.; Zhang, J.; Wu, C.; Ke, Q.; Han, K.; Zheng, W.; Xu, N. Transcriptome analysis of the Larimichthys crocea liver in response to Cryptocaryon irritans. Fish Shellfish. Immunol. 2016, 48, 1–11. [Google Scholar] [CrossRef]
- Chen, G.; He, L.; Luo, L.; Huang, R.; Liao, L.; Li, Y.; Zhu, Z.; Wang, Y. Transcriptomics Sequencing Provides Insights into Understanding the Mechanism of Grass Carp Reovirus Infection. Int. J. Mol. Sci. 2018, 19, 488. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, B.; Wang, N.; Ouyang, G.; Cao, H. Transcriptome analysis of rare minnow (Gobiocypris rarus) infected by the grass carp reovirus. Fish Shellfish. Immunol. 2019, 89, 337–344. [Google Scholar] [CrossRef]
- Ma, J.; Xu, C.; Zhou, Y.; Jiang, N.; Xue, M.; Cao, J.; Li, S.; Fan, Y. Metabolomics in rare minnow (Gobiocypris rarus) after infection by attenuated and virulent grass carp reovirus genotype II. Fish Shellfish. Immunol. 2023, 138, 108840. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, A.; Raza, S.Q.; Voisin, L.; Law, F.; Pipoli da Fonseca, J.; Caillet, M.; Kroemer, G.; Perfettini, J.L. Multifaceted roles of purinergic receptors in viral infection. Microbes Infect. 2012, 14, 1278–1283. [Google Scholar] [CrossRef]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef] [PubMed]
- Siebler, J.; Wirtz, S.; Klein, S.; Protschka, M.; Blessing, M.; Galle, P.R.; Neurath, M.F. A key pathogenic role for the STAT1/T-bet signaling pathway in T-cell-mediated liver inflammation. Hepatology 2003, 38, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Goerge, T.; Ho-Tin-Noe, B.; Carbo, C.; Benarafa, C.; Remold-O’Donnell, E.; Zhao, B.; Cifuni, S.M.; Wagner, D.D. Inflammation induces hemorrhage in thrombocytopenia. Blood 2008, 111, 4958–4964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, Y.; Zhang, Y.; Liu, Y.; Xiang, Z.; Qu, F.; Yu, Z. Cloning and characterization of three suppressors of cytokine signaling (SOCS) genes from the Pacific oyster, Crassostrea gigas. Fish Shellfish. Immunol. 2015, 44, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Sun, L. Comparative analysis of the expression patterns of eight suppressors of cytokine signaling in tongue sole, Cynoglossus semilaevis. Fish Shellfish. Immunol. 2016, 55, 595–601. [Google Scholar] [CrossRef]
- Schwake, M.; Schröder, B.; Saftig, P. Lysosomal membrane proteins and their central role in physiology. Traffic 2013, 14, 739–748. [Google Scholar] [CrossRef]
- Wang, R.; Song, L.; Su, B.; Zhao, H.; Zhang, D.; Peatman, E.; Li, C. Mucosal expression signatures of two Cathepsin L in channel catfish (Ictalurus punctatus) following bacterial challenge. Fish Shellfish. Immunol. 2015, 47, 582–589. [Google Scholar] [CrossRef]
- Gao, C.; Fu, Q.; Su, B.; Zhou, S.; Liu, F.; Song, L.; Zhang, M.; Ren, Y.; Dong, X.; Tan, F.; et al. Transcriptomic profiling revealed the signatures of intestinal barrier alteration and pathogen entry in turbot (Scophthalmus maximus) following Vibrio anguillarum challenge. Dev. Comp. Immunol. 2016, 65, 159–168. [Google Scholar] [CrossRef]
- He, L.; Zhu, D.; Liang, X.; Li, Y.; Liao, L.; Yang, C.; Huang, R.; Zhu, Z.; Wang, Y. Multi-Omics Sequencing Provides Insights Into Age-Dependent Susceptibility of Grass Carp (Ctenopharyngodon idellus) to Reovirus. Front. Immunol. 2021, 12, 694965. [Google Scholar] [CrossRef]
- Adams, O.; Besken, K.E.; Oberdörfer, C.; MacKenzie, C.R.; Rüssing, D.; Däubener, W. Inhibition of human herpes simplex virus type 2 by interferon gamma and tumor necrosis factor alpha is mediated by indoleamine 2,3-dioxygenase. Microbes Infect. 2004, 6, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Yi, X.; Sun, Y.; Bi, X.; Du, J.; Zhang, C.; Quan, S.; Zhang, F.; Sun, R.; Qian, L.; et al. Proteomic and Metabolomic Characterization of COVID-19 Patient Sera. Cell 2020, 182, 59–72.e15. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Vandenabeele, P. Necroptosis and its role in inflammation. Nature 2015, 517, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.J.; Nogusa, S.; Chen, P.; Maki, J.L.; Lerro, A.; Andrake, M.; Rall, G.F.; Degterev, A.; Balachandran, S. Interferon-induced RIP1/RIP3-mediated necrosis requires PKR and is licensed by FADD and caspases. Proc. Natl. Acad. Sci. USA 2013, 110, E3109–E3118. [Google Scholar] [CrossRef] [Green Version]
- Varfolomeev, E.E.; Schuchmann, M.; Luria, V.; Chiannilkulchai, N.; Beckmann, J.S.; Mett, I.L.; Rebrikov, D.; Brodianski, V.M.; Kemper, O.C.; Kollet, O.; et al. Targeted disruption of the mouse Caspase 8 gene ablates cell death induction by the TNF receptors, Fas/Apo1, and DR3 and is lethal prenatally. Immunity 1998, 9, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baritaud, M.; Cabon, L.; Delavallée, L.; Galán-Malo, P.; Gilles, M.E.; Brunelle-Navas, M.N.; Susin, S.A. AIF-mediated caspase-independent necroptosis requires ATM and DNA-PK-induced histone H2AX Ser139 phosphorylation. Cell Death Dis. 2012, 3, e390. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Usui, K.; Hiraki, T. Fluconazole modulates membrane rigidity, heterogeneity, and water penetration into the plasma membrane in Saccharomyces cerevisiae. Biochemistry 2009, 48, 8494–8504. [Google Scholar] [CrossRef]
- Verri, T.; Barca, A.; Pisani, P.; Piccinni, B.; Storelli, C.; Romano, A. Di- and tripeptide transport in vertebrates: The contribution of teleost fish models. J. Comp. Physiol. B 2017, 187, 395–462. [Google Scholar] [CrossRef]
- Ge, W.; Hou, C.; Zhang, W.; Guo, X.; Gao, P.; Song, X.; Gao, R.; Liu, Y.; Guo, W.; Li, B.; et al. Mep1a contributes to Ang II-induced cardiac remodeling by promoting cardiac hypertrophy, fibrosis and inflammation. J. Mol. Cell. Cardiol. 2021, 152, 52–68. [Google Scholar] [CrossRef]
- Meyer-Schwesinger, C. The ubiquitin-proteasome system in kidney physiology and disease. Nat. Rev. Nephrol. 2019, 15, 393–411. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; Bertolotti, A. Regulation of proteasome assembly and activity in health and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 697–712. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Alonso, J.L.; Ostuni, E.; Whitesides, G.M.; Ingber, D.E. Cell shape provides global control of focal adhesion assembly. Biochem. Biophys. Res. Commun. 2003, 307, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Frame, M.C.; Patel, H.; Serrels, B.; Lietha, D.; Eck, M.J. The FERM domain: Organizing the structure and function of FAK. Nat. Rev. Mol. Cell Biol. 2010, 11, 802–814. [Google Scholar] [CrossRef]
- Hsia, D.A.; Mitra, S.K.; Hauck, C.R.; Streblow, D.N.; Nelson, J.A.; Ilic, D.; Huang, S.; Li, E.; Nemerow, G.R.; Leng, J.; et al. Differential regulation of cell motility and invasion by FAK. J. Cell Biol. 2003, 160, 753–767. [Google Scholar] [CrossRef]
- Fouquet, B.; Nikolic, J.; Larrous, F.; Bourhy, H.; Wirblich, C.; Lagaudrière-Gesbert, C.; Blondel, D. Focal adhesion kinase is involved in rabies virus infection through its interaction with viral phosphoprotein P. J. Virol. 2015, 89, 1640–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Ruan, L.; Xu, X. An immediate-early protein of white spot syndrome virus modulates the phosphorylation of focal adhesion kinase of shrimp. Virology 2011, 419, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.H. Examining coagulation-complement crosstalk: Complement activation and thrombosis. Thromb. Res. 2016, 141 (Suppl. 2), S50–S54. [Google Scholar] [CrossRef]
- Calame, D.G.; Mueller-Ortiz, S.L.; Wetsel, R.A. Innate and adaptive immunologic functions of complement in the host response to Listeria monocytogenes infection. Immunobiology 2016, 221, 1407–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamad, O.A.; Bäck, J.; Nilsson, P.H.; Nilsson, B.; Ekdahl, K.N. Platelets, complement, and contact activation: Partners in inflammation and thrombosis. Adv. Exp. Med. Biol. 2012, 946, 185–205. [Google Scholar] [CrossRef]
- Meri, S. Complement activation in diseases presenting with thrombotic microangiopathy. Eur. J. Intern. Med. 2013, 24, 496–502. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines for Malaria, 13 July 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| β-actin | ATGGATGATGAAATTGCCGC | CTGTCCCATACCAACCATGA |
| GP1BB | GAACTCCACCTGAACGACAA | TCAGATAAAGGATGCCGCAG |
| MHCⅠ | CCTATGCTGGACAACACTCT | TTTGGCACAGCTTTCATTGC |
| PLAUR | TAACGTGCCCAATGGAAAGA | TACAACCAAAGACTGGCCTC |
| IRF3 | TCAGTGGGAAGATCAACGAG | TGAGGACGGATAATGCGAAA |
| CTSB | CTGGCTACAGCCCTTCTTAC | AGGAAGTCCTCATACACGGT |
| CXCL8 | CAGTCTTTGTTATCGCTGGC | ATGTGTTTACCAATGCGTCG |
| HBA | AGACCTATTTCGCTCACTGG | GAATGCATGAAGTTCGCTCA |
| STAT1B | AGCACTACAGCCGTCTCAATCT | CCTGTGATGAGTTACCGCTACCTT |
| Pathway | Gene Name | Log2 (Fold Change) | |
|---|---|---|---|
| QJ205 | DY197 | ||
| Representative upregulated differentially expressed genes | |||
| RIG-Ⅰ/TOLL/NOD-like receptor signaling pathway, JAK-STAT signaling pathway | Retinoic acid-inducible protein I | 0.11 | 2.01 |
| DEXH (Asp-Glu-X-His) box polypeptide 58 | 0.28 | 2.47 | |
| Melanoma differentiation associated gene 5 | −0.07 | 1.72 | |
| Toll-like receptor 3 | −0.17 | 1.03 | |
| Toll-like receptor 8 | −0.97 | 1.02 | |
| Interferon regulatory factor 3 | 0.09 | 1.06 | |
| Interferon regulatory factor 7 | 0.16 | 2.44 | |
| Interferon alpha | −0.81 | 1.60 | |
| Interferon beta | −0.56 | 3.85 | |
| Signal transducer and activator of transcription 1b | −0.13 | 2.24 | |
| Signal transducer and activator of transcription 2 | −0.34 | 1.09 | |
| Tyrosine-protein kinase JAK1 | −0.61 | 1.27 | |
| Chemokine (C-X-C motif) ligand 11 | −0.37 | 1.79 | |
| Chemokine (C-X-C motif) ligand 20 | −0.11 | 2.23 | |
| Suppressor of cytokine signaling 1 | 0.08 | 2.06 | |
| Suppressor of cytokine signaling 3 | −0.44 | 1.18 | |
| Suppressor of cytokine signaling 6 | −0.84 | 1.38 | |
| B cell receptor signaling pathway | Immunoglobulin omega chain | −0.70 | 1.96 |
| B-cell receptor CD22 | −1.03 | 1.05 | |
| Secreted immunoglobulin heavy chain IgM | −0.73 | 1.16 | |
| Immunoglobulin heavy variable 1–3 | −0.59 | 1.81 | |
| Tryptophan metabolism | kynurenine-oxoglutarate transaminase 1 | −0.62 | 1.34 |
| Lysosome and phagosome | Cathepsin A | −0.69 | 1.10 |
| Cathepsin B | −0.65 | 1.05 | |
| Cathepsin D | −0.82 | 1.17 | |
| Cathepsin K | −0.30 | 2.33 | |
| Cathepsin L | 0.16 | 1.83 | |
| Cathepsin S | −0.71 | 1.06 | |
| Cathepsin Z | −0.64 | 1.08 | |
| MHC class I antigen | −0.61 | 1.17 | |
| Apoptosis and necroptosis | Bcl-2 associated X protein | −0.19 | 1.64 |
| Caspase-7 | −0.66 | 1.81 | |
| Interferon-induced, double-stranded RNA-activated protein kinase | 0.10 | 1.56 | |
| Complement and coagulation cascades | Coagulation factor IXa | 1.05 | 0.21 |
| Urokinase plasminogen activator surface receptor | 0.18 | 3.41 | |
| Complement factor H | 0.58 | 1.48 | |
| Representative downregulated differentially expressed genes | |||
| T cell receptor signaling pathway | T-cell surface glycoprotein CD3 zeta chain | −1.26 | −1.62 |
| CD8 alpha chain | −0.93 | −2.17 | |
| Tyrosine-protein kinase ZAP-70 | −0.63 | −1.45 | |
| Steroid biosynthesis | Squalene synthase | 1.12 | −0.48 |
| Lanosterol synthase | 1.39 | −1.50 | |
| Delta(24)-sterol reductase | 1.37 | −0.77 | |
| Squalene monooxygenase | 1.86 | −2.10 | |
| Malaria | Hemoglobin subunit alpha | −1.36 | −3.21 |
| Hemoglobin subunit beta-2 | −0.33 | −1.98 | |
| Thrombospondin-1 | −1.08 | −1.79 | |
| Platelet activation | Platelet glycoprotein V | −1.15 | −1.96 |
| Platelet glycoprotein Ib alpha chain | −0.95 | −1.80 | |
| Platelet glycoprotein Ib beta chain | −1.26 | −2.04 | |
| Focal adhesion and regulation of actin cytoskeleton | Myosin regulatory light polypeptide 9b | −1.31 | −1.67 |
| Myosin light chain kinase | −0.92 | −1.62 | |
| Integrin alpha-2 | −1.09 | −1.66 | |
| Platelet-derived growth factor subunit B | −1.42 | −2.06 | |
| Laminin subunit beta-1b | −0.69 | −1.67 | |
| Vascular endothelial growth factor C | −1.07 | −1.48 | |
| Pathway | Gene Name | Log2 (Fold Change) | |
|---|---|---|---|
| QJ205 | DY197 | ||
| Representative upregulated differentially expressed genes | |||
| RIG-Ⅰ/TOLL/NOD-like receptor singaling pathway, JAK-STAT singaling pathway | Retinoic acid-inducible protein I | 1.95 | 3.99 |
| DEXH (Asp-Glu-X-His) box polypeptide 58 | 2.29 | 4.70 | |
| Melanoma differentiation associated gene 5 | 0.93 | 3.13 | |
| Nucleotide-binding oligomerization domain-containing protein | 0.02 | 1.67 | |
| Nicotinamide phosphoribosyltransferase-like protein | 1.05 | 3.54 | |
| Toll-like receptor 8 | 0.52 | 3.10 | |
| TNF receptor-associated factor 5 | 0.16 | 2.47 | |
| Interferon regulatory factor 7 | 2.85 | 5.77 | |
| Interferon regulatory factor 9 | 0.56 | 2.49 | |
| Interferon a | 0.71 | 3.86 | |
| Signal transducer and activator of transcription 1b | 2.08 | 4.79 | |
| Tyrosine-protein kinase JAK1 | −0.04 | 1.74 | |
| Cytokine receptor family member b4 | −0.03 | 1.96 | |
| C-X-C motif chemokine 9 | 1.10 | 3.90 | |
| Chemokine (C-X-C motif) ligand 20 | 0.56 | 4.47 | |
| Suppressor of cytokine signaling 1 | 0.87 | 2.52 | |
| Suppressor of cytokine signaling 3 | 0.21 | 1.79 | |
| B cell receptor signaling pathway | Immunoglobulin omega chain | 0.00 | 8.06 |
| B-cell receptor CD22 | 0.58 | 2.81 | |
| Secreted immunoglobulin heavy chain IgM | 0.73 | 2.08 | |
| Tryptophan metabolism | kynurenine-oxoglutarate transaminase 1 | 0.61 | 1.81 |
| Lysosome and phagosome | Cathepsin A | −0.05 | 1.15 |
| Cathepsin B | −0.02 | 1.56 | |
| Cathepsin H | 0.27 | 1.81 | |
| Cathepsin K | 1.19 | 3.87 | |
| Cathepsin L | 0.07 | 1.53 | |
| Cathepsin Z | −0.12 | 1.82 | |
| MHC class I antigen | 0.20 | 3.28 | |
| Apoptosis and necroptosis | Bcl-2 associated X protein | −0.19 | 1.64 |
| Caspase-7 | 0.29 | 2.69 | |
| Caspase-8 | 1.65 | 3.75 | |
| Interferon-induced, double-stranded RNA-activated protein kinase | 1.16 | 2.80 | |
| Tumor necrosis factor receptor superfamily member 1A | 0.24 | 1.98 | |
| Tumor necrosis factor (ligand) superfamily, member 10 | 1.94 | 3.00 | |
| Proteasome | Proteasome subunit alpha type-3 | 0.10 | 1.50 |
| Proteasome subunit alpha type-5 | −0.06 | 1.27 | |
| Proteasome subunit alpha type-6 | 0.51 | 2.25 | |
| Proteasome subunit beta type-2 | −0.33 | 1.36 | |
| Proteasome subunit beta type-7 | 0.99 | 2.97 | |
| Proteasome subunit beta type-8 | 0.54 | 2.97 | |
| Proteasome activator complex subunit 1 | 0.38 | 2.33 | |
| Proteasome activator complex subunit 2 | 0.46 | 2.81 | |
| Protein digestion and absorption | Meprin A subunit alpha | 2.14 | 3.49 |
| Meprin A subunit beta | 2.80 | 3.19 | |
| Solute carrier family 15 member 1 | 3.26 | 3.05 | |
| Complement and coagulation cascades | Complement C1q subcomponent subunit A | 0.51 | 2.49 |
| Proteinase-activated receptor 1 | −0.08 | 1.79 | |
| Urokinase plasminogen activator surface receptor | 3.96 | 7.69 | |
| Complement factor H | −0.19 | 1.44 | |
| Representative downregulated differentially expressed genes | |||
| Steroid biosynthesis | Squalene synthase | −0.24 | −1.52 |
| Lanosterol synthase-like isoform X2 | −0.75 | −2.02 | |
| Delta(24)-sterol reductase | −0.02 | −1.14 | |
| Squalene monooxygenase | −0.98 | −2.59 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Xu, C.; Jiang, N.; Meng, Y.; Zhou, Y.; Xue, M.; Liu, W.; Li, Y.; Fan, Y. Transcriptomics in Rare Minnow (Gobiocypris rarus) towards Attenuated and Virulent Grass Carp Reovirus Genotype II Infection. Animals 2023, 13, 1870. https://doi.org/10.3390/ani13111870
Ma J, Xu C, Jiang N, Meng Y, Zhou Y, Xue M, Liu W, Li Y, Fan Y. Transcriptomics in Rare Minnow (Gobiocypris rarus) towards Attenuated and Virulent Grass Carp Reovirus Genotype II Infection. Animals. 2023; 13(11):1870. https://doi.org/10.3390/ani13111870
Chicago/Turabian StyleMa, Jie, Chen Xu, Nan Jiang, Yan Meng, Yong Zhou, Mingyang Xue, Wenzhi Liu, Yiqun Li, and Yuding Fan. 2023. "Transcriptomics in Rare Minnow (Gobiocypris rarus) towards Attenuated and Virulent Grass Carp Reovirus Genotype II Infection" Animals 13, no. 11: 1870. https://doi.org/10.3390/ani13111870