Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure
by
Amandine Thépault
1,2, 
Valérie Rose
1,
Marilyne Queguiner
1,
Marianne Chemaly
1 and
Katell Rivoal
1,* 1
ANSES, Ploufragan-Plouzané-Niort Laboratory, Hygiene and Quality of Poultry and Pig Products Unit, BP53, 22440 Ploufragan, France
2
French Agency for Food, Environmental and Occupational Health & Safety, Rennes 1 University, rue du Thabor, 35000 Rennes, France
*
Author to whom correspondence should be addressed.
Animals 2020, 10(5), 838; https://doi.org/10.3390/ani10050838
Submission received: 27 March 2020
/
Revised: 23 April 2020
/
Accepted: 30 April 2020
/
Published: 12 May 2020
(This article belongs to the Section Companion Animals)
Abstract
:Simple Summary
Campylobacter is among the most common causes of bacterial gastroenteritis and is associated with post-infectious neuropathies. This organism is part of the commensal microbiota of numerous host species, including companion animals. Molecular typing approaches have been used to attribute the source of human campylobacteriosis and showed that broilers are a major infection source, while pets are a non-negligible one (10–25% of clinical cases). Since the assessment of animal colonization by Campylobacter is crucial to better understand its epidemiology, we determined the Campylobacter carriage by dogs and cats, hereafter defined as pets, and characterized genetically Campylobacter jejuni (C. jejuni) isolates. Pets appeared to frequently carry Campylobacter. The comparison of genetic profiles of C. jejuni isolates with isolates from several animal reservoirs and clinical cases revealed an overlap between profiles. These results suggest potential pets’ contamination by livestock or inversely, as well as their potential role in Campylobacter transmission to humans. However, some pets’ profiles were not isolated in livestock, which suggests the existence of other sources of pet contamination by C. jejuni or implies that pets may constitute a reservoir for Campylobacter with specific profiles. Since frequent contact occurs between pets and humans, this work emphasized the potential role of pets as a source of human exposure to Campylobacter.
Abstract
Assessing the carriage of Campylobacter in animal reservoirs is essential to better understand Campylobacter epidemiology. Here, we evaluated the prevalence of thermophilic Campylobacter spp. in dogs and cats, hereafter defined as pets, and characterized Campylobacter jejuni (C. jejuni) isolates to assess their genetic diversity and their potential link with isolates from other animals or human cases. During a 6-month period, 304 feces samples were collected from pets. A significantly higher prevalence of thermophilic Campylobacter spp. was found in dogs compared with cats, as well as in dogs ≤ 1-year-old compared with older dogs. C. jejuni was the predominant species found in pets, and its genomic characterization revealed a high genetic diversity. Genotypes comparison with previously characterized isolates revealed a partial overlap between C. jejuni isolates from pets, chicken, cattle, and clinical cases. This overlap suggests the potential role of livestock and humans in pets’ exposure to Campylobacter, or vice versa. The isolation of pets’ specific profiles may suggest the existence of other sources of pet contamination or imply that pets may constitute a reservoir for Campylobacter. Because of the proximity between humans and pets, along with their frequent carriage of C. jejuni, human exposure to Campylobacter from pets can be more important than previously thought.
1. Introduction
Campylobacter spp. are the leading cause of the main bacterial foodborne zoonosis in Europe, with about 246,571 cases reported in 2018 [1]. Campylobacter jejuni (C. jejuni) is the most frequent causative species of campylobacteriosis followed by C. coli. In France, these species were respectively responsible for nearly 80 and 15% of human infections, while C. fetus, C. lari and C. upsaliensis accounted for 4, 0.4, and 0.1% of campylobacteriosis cases that occurred between 2003 and 2010, respectively [2]. Several risk factors for human infection by Campylobacter spp. are related to the consumption of contaminated foodstuffs, such as chicken meat, unpasteurized milk, or untreated water. However, traveling abroad and contact with farm animals or pets also constituted risk factors for human campylobacteriosis [3,4,5]. Several source attribution studies quantifying the relative importance of animal reservoirs in campylobacteriosis identified chicken and cattle as the main source of human infection by Campylobacter spp. [6,7,8,9,10]. However, 1 to 25% of human campylobacteriosis are attributable to pets (including cats and dogs) in several European countries [11,12,13], where heterogeneous levels of Campylobacter spp. carriage by pets were highlighted [12,14,15].
Thus, cats and dogs (hereafter defined as pets) appear to be potential important sources of human infection by Campylobacter. To better understand the epidemiology of Campylobacter spp. in their natural reservoirs, several prevalence and genotyping studies have been performed in chicken, cattle, or environmental water in France. These studies revealed high rates of Campylobacter spp. isolation and the overlap between genetic profiles found in C. jejuni isolates from animal reservoirs and human campylobacteriosis, emphasizing the potential involvement of these sources in human infection [16,17,18,19]. However, no such study has been performed in pets in France, despite a possible implication of pets in transmission of Campylobacter spp. to humans, as previously suggested in Europe [12].
Therefore, this study aimed to evaluate the carriage of thermophilic Campylobacter spp. in pets. Then, C. jejuni isolates circulating in these animals were genotyped to assess their genetic diversity and their potential link with animal and clinical isolates through the comparison of their genetic profiles. For this purpose, Comparative Genomic Fingerprinting, using 40 assay genes (CGF40) [20], was used to describe C. jejuni isolates since this technique shows good concordance [17,20] with Multilocus Sequence Typing (MLST) [21].
2. Materials and Methods
2.1. Collection of Feces Samples from Pets and Bacterial Analyses
In December 2014 and from April to October 2015, 304 feces samples from cats (n = 70) and dogs (n = 234) were collected on a voluntary basis (from 4 veterinarians, 2 breeders/ owners of cat and dog boarding house, and 26 private owners) in the administrative department of Côtes d’Armor in Brittany, France. Each feces sample collected was accompanied by a questionnaire informing on the age of the animal, the health status, the pet living conditions (indoor/outdoor/both), and the season of sampling. For the detection of thermophilic Campylobacter spp., ten grams of feces were diluted (ratio 1:10) in Bolton broth (Thermofisher Diagnostic, Dardilly, France) and homogenized for 45 s. Ten microliters of each inoculated Bolton broth were streaked onto mCCDA (modified Charcoal Cefoperazone Deoxycholate Agar) and Butzler n°2 (virion) (Thermofisher Diagnostic, Dardilly, France) plates and incubated for 72 h at 41.5 °C under microaerobic conditions. After an enrichment of the inoculated Bolton broths for 24 h at 41.5 °C, 10 µL of the mixtures were plated onto mCCDA and Butzler n°2 (virion) agar media and incubated for 48 h at 41.5 °C under microaerobic conditions. For each positive plate of each media, up to five typical Campylobacter colonies were subcultured onto blood agar plates for species determination by Maldi-Tof (Matrix-Assisted Laser Desorption/Ionization-Time of Flight). The isolates were stored at −70 °C in peptone broth containing 20% (v/v) glycerol, and only C. jejuni isolates were further characterized in this study.
2.2. CGF40 Typing
DNA was extracted using InstagenMatrix® (Biorad, Marnes-la-Coquette, France) according the supplier recommendations. The forty accessory genes for CGF [20] were amplified according to previously published experimental conditions [17]. The CGF fingerprints were visualized using a standard gel electrophoresis containing 2% of agarose colored with GelRed® (Interchim, Montluçon, France) in accordance with supplier recommendations. DNA from the reference strain NCTC11168, known to contain the 40 assay genes, was used as positive control in all assays.
The PCR results were converted into binary data corresponding to the absence (0) or the presence (1) of the marker in bacterial genomes and were stored into BioNumerics® software (v6.5, Applied Maths, Belgium). A dendrogram was built using the simple matching distance coefficient and unweighted-pair group method using average linkages (UPGMA) of clustering in BioNumerics® as previously described [20] and using 100% of similarity for cluster definition to describe the population structure. Then, a cut-off of 90% of similarity was used for cluster definition to compare pet C. jejuni isolates from this study to previously published French C. jejuni isolates from chicken, cattle and human campylobacteriosis [17,18,22]. Since no universal and fixed nomenclature exists regarding CGF cluster definition, the CGF 40-90% profiles of the previously published isolates were renumbered in this study.
2.3. Data Analysis
The Campylobacter status of the animals has been assessed with three main factors: (i) the kind of pets (cats or dogs), the season of sampling (spring, summer or fall) and the pet living condition (inside or outside the house or both). A multi-factor logistic regression was performed using the glm function of the R software. To assess the statistical difference between the carriage of Campylobacter spp. within pets according to their age, Fisher exact tests were used. In addition, the genetic diversity was assessed within C. jejuni isolates using the Simpson’s diversity index calculated with the online tool “Comparing Partitions” from the website http://www.comparingpartitions.info/ [23]. The calculation of the confidence intervals for clustering agreement measures at 95% was performed using the resampling technique jackknife [24].
3. Results
3.1. Campylobacter Isolation from Pet Feces and Species Identification
In this study, 304 feces samples from cats and dogs were collected: 234 samples were from dogs and 70 from cats. One hundred seventeen samples were collected in spring (89 dogs and 28 cats), 127 in summer (109 dogs and 18 cats), and 60 (36 dogs and 24 cats) in fall. According to the completed questionnaire, the age of animals ranged from 3 weeks to 17 years in dogs, and from 3 months to 15 years in cats, but no age information was collected for 5 dogs and 2 cats. Health status regarding diarrhea was also recorded, with only 3 dogs and 1 cat showed diarrhea symptoms at the sampling date. Among the 304 animals, 192 lived outdoor and indoor alternatively (156 dogs and 36 cats), 67 exclusively outdoor (65 dogs and 2 cats), 41 exclusively indoor pets (32 cats and 9 dogs), and no data were available for 4 dogs.
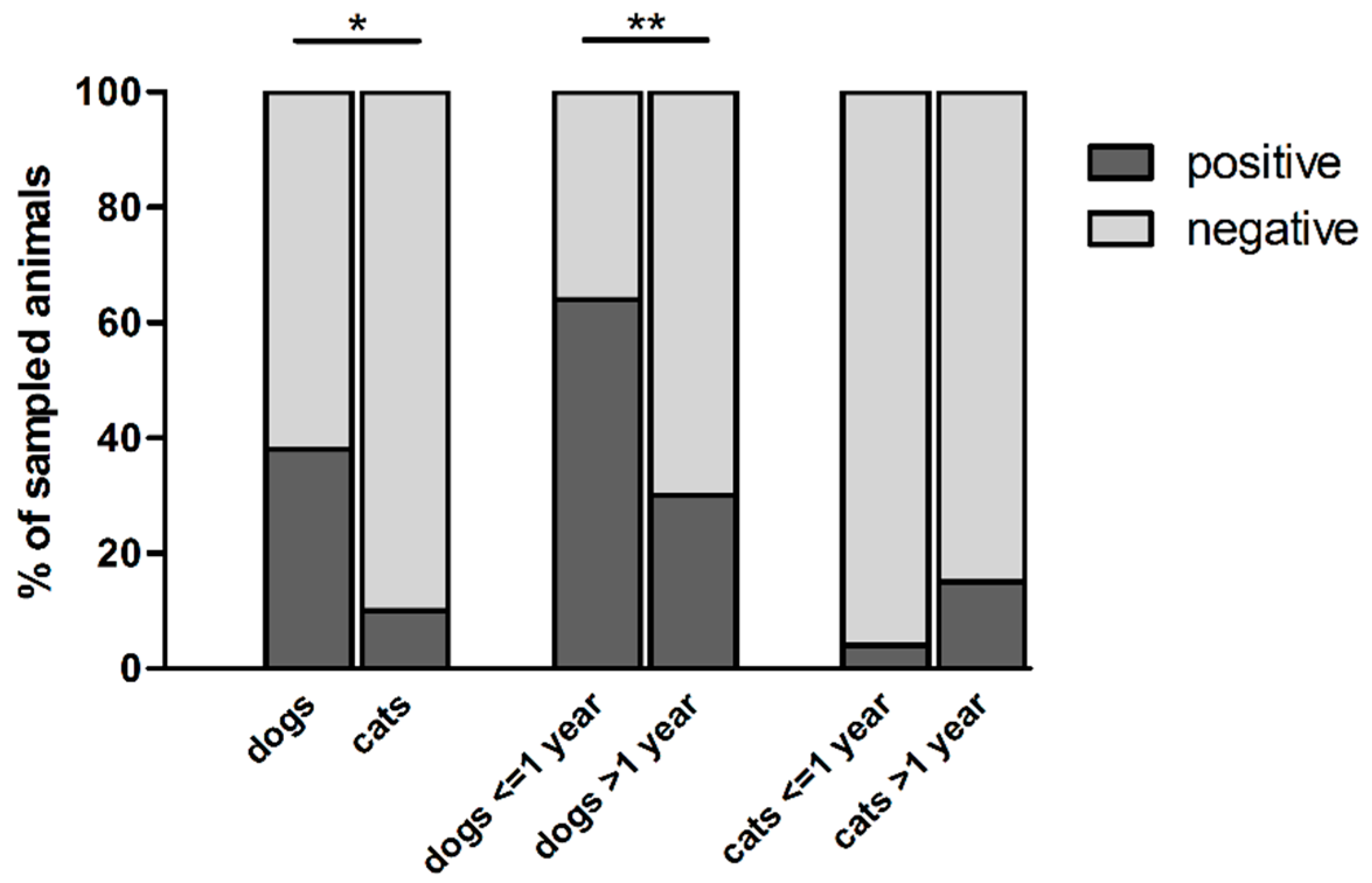
Within the 304 pet feces samples collected, thermophilic Campylobacter spp. were isolated from 89 dogs and 7 cats, which correspond to a presence in 38 and 10% of the animals, respectively (Table 1; Figure 1). Dogs carried significantly more thermophilic Campylobacter spp. than cats (p < 0.0001), as well as puppies (dogs ≤ 1-year-old) compared with adult dogs (p < 0.0001) (Figure 1). Indeed, 63.6% (35/55) of puppies carried thermophilic Campylobacter spp., in comparison with 30.5% of adult dogs (53/174), while these bacteria were isolated from 20% (1/5) of dogs without age information (Table 1). No significant differences were observed in the carriage of thermophilic Campylobacter spp. in cats according to their age (Table 1; Figure 1). Campylobacter was not isolated from feces of animals showing diarrhea (n = 4) at the sampling date (data not shown). Because of the low number of animals showing these symptoms (4/304), its impact on Campylobacter carriage cannot be assessed in our study. The season of sampling, as well as the living conditions of pets, did not influence significantly Campylobacter carriage by pets (p > 0.05).
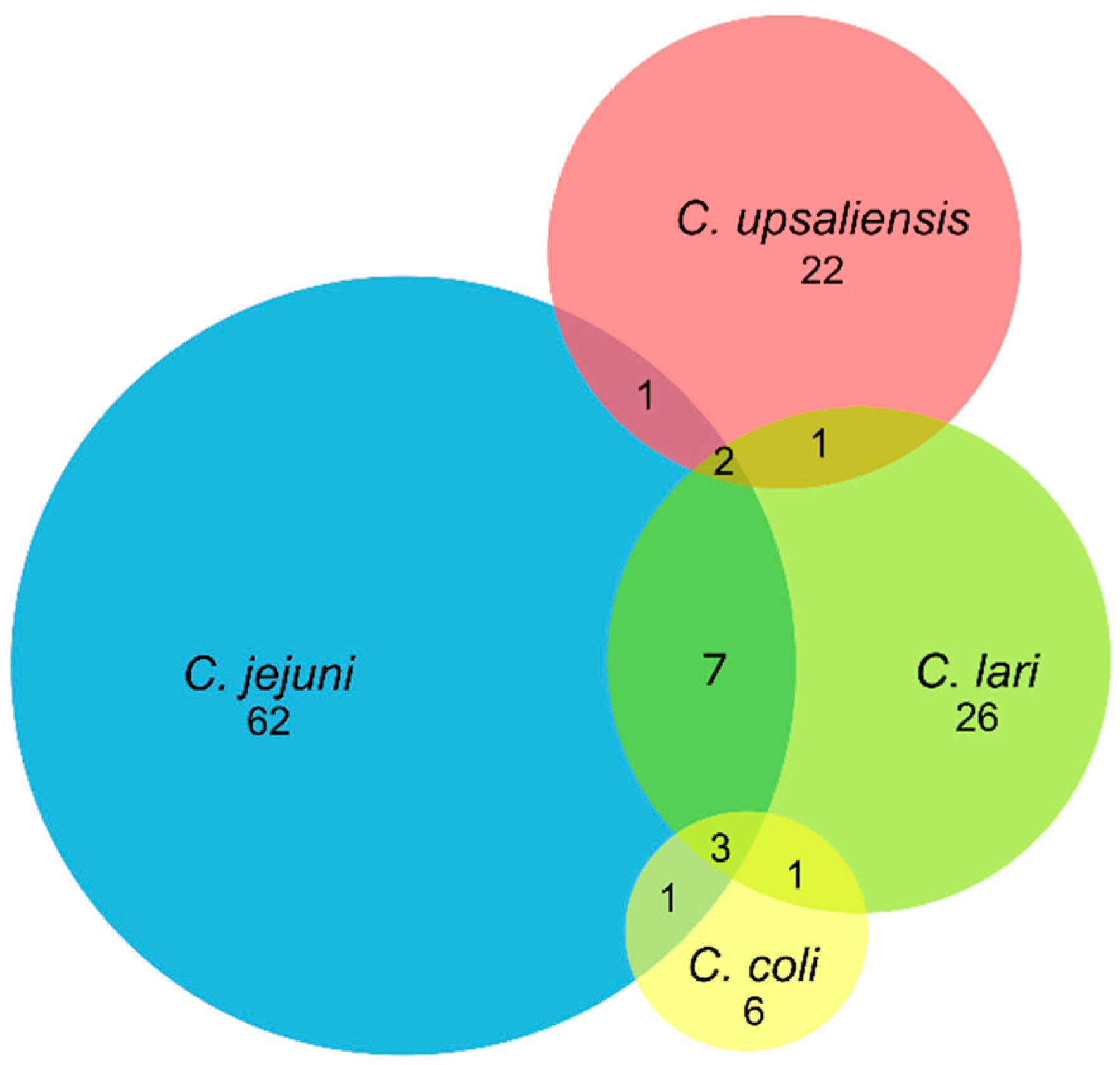
C. jejuni was the main species isolated in pets, followed by C. lari, C. upsaliensis, and C. coli in dogs, or C. upsaliensis in cats (Table 1; Figure 2). C. jejuni was found in 64.0% (57/89) and 71.4% (5/7) of Campylobacter-positive dogs and cats, which corresponds to an overall C. jejuni prevalence of 24.4 and 7.1% in these animals, respectively (Table 1). Simultaneous carriages of several species of Campylobacter were observed in 16 dogs (Figure 2). Seven of them carried C. jejuni and C. lari, while carriages of C. coli/C. lari, C. jejuni/C. coli, C. jejuni/C. upsaliensis, or C. upsaliensis/C. lari were each observed in one dog. In few animals, up to three different species of thermophilic Campylobacter were isolated. Three dogs carried C. jejuni, C. coli, and C. lari, while 2 others carried C. jejuni, C. lari and C. upsaliensis.
3.2. Genotyping of C. jejuni Isolates Using Comparative Genomic Fingerprinting, Using 40 Assay Genes (CGF40)
Among 494 isolates of C. jejuni collected from 62 C. jejuni-positive feces samples, the characterization using CGF40 was performed on 455 C. jejuni isolates, since no growth was observed during subculture on agar plates for 39 isolates collected from 6 animals. Within the 455 genotyped isolates and based on 100% of similarity for cluster definition, 135 clusters were obtained (hereafter defined as CGF40-100% clusters). In 34 animals, from 2 to 10 different CGF40-100% clusters were detected, while 22 animals carried only one CGF40-100% cluster. Since C. jejuni isolates collected from the same individual animal and showing identical CGF40-100% cluster are likely to be the same strain, because of the high discriminatory power of CGF40-100% [17,20], one isolate of each CGF40-100% cluster found was kept for each animal in order to analyze the genetic structure of C. jejuni population.
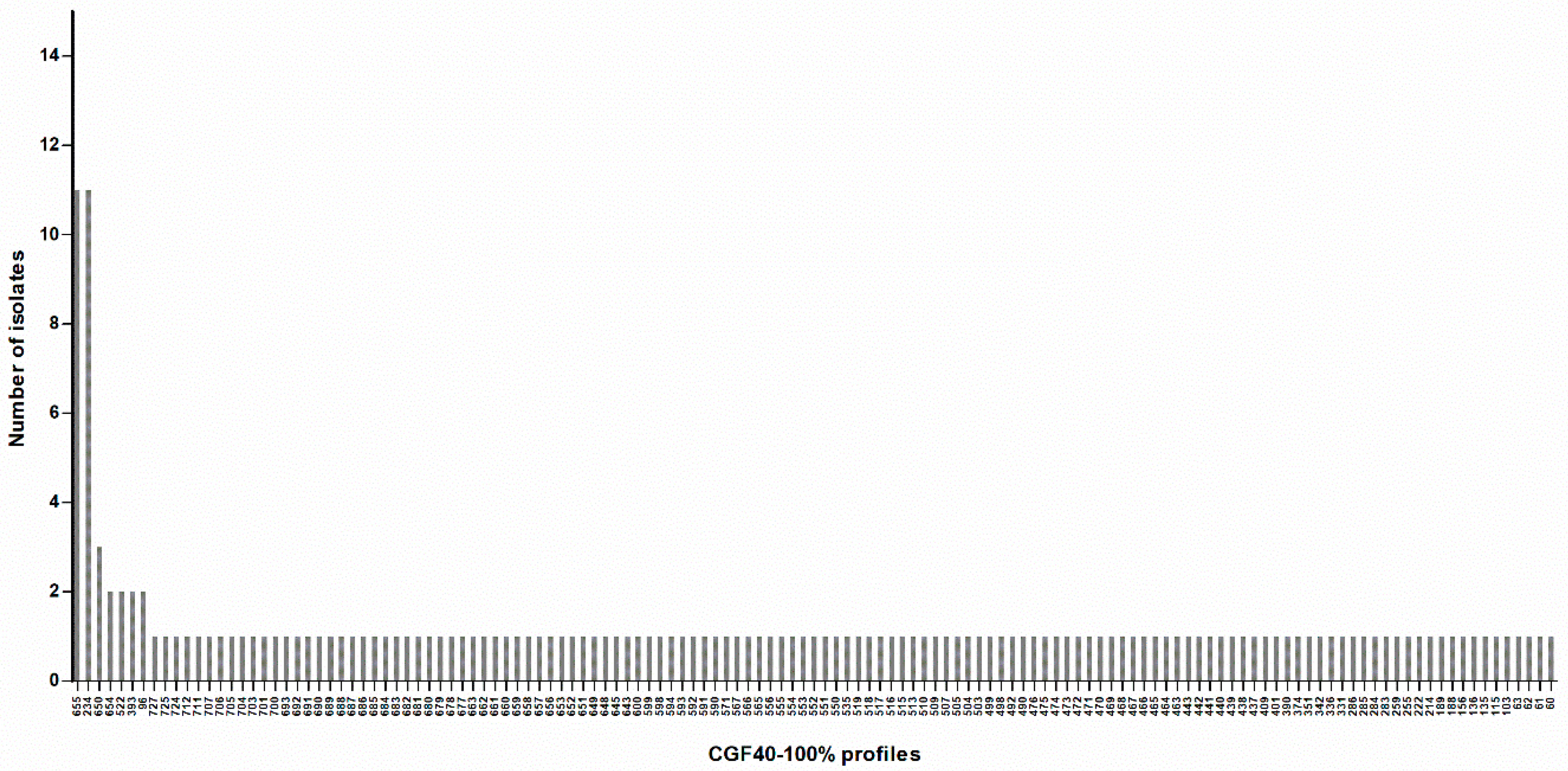
Finally, this population comprised 161 C. jejuni isolates with 135 different CGF40-100% clusters (Figure 3; Supplementary Table S1). A Simpson’s diversity index of 0.991 (CI95% 0.984–0.998) was calculated, indicating a high genetic diversity among C. jejuni isolated from pets. Indeed, 79.5% of isolates (128/161) had a CGF40-100% profile found only once in the population (Figure 3; Supplementary Table S1). However, 2 profiles were predominant (profiles 234 and 655), including each 11 isolates, which represent 6.8% of the population (Figure 3; Supplementary Table S1).
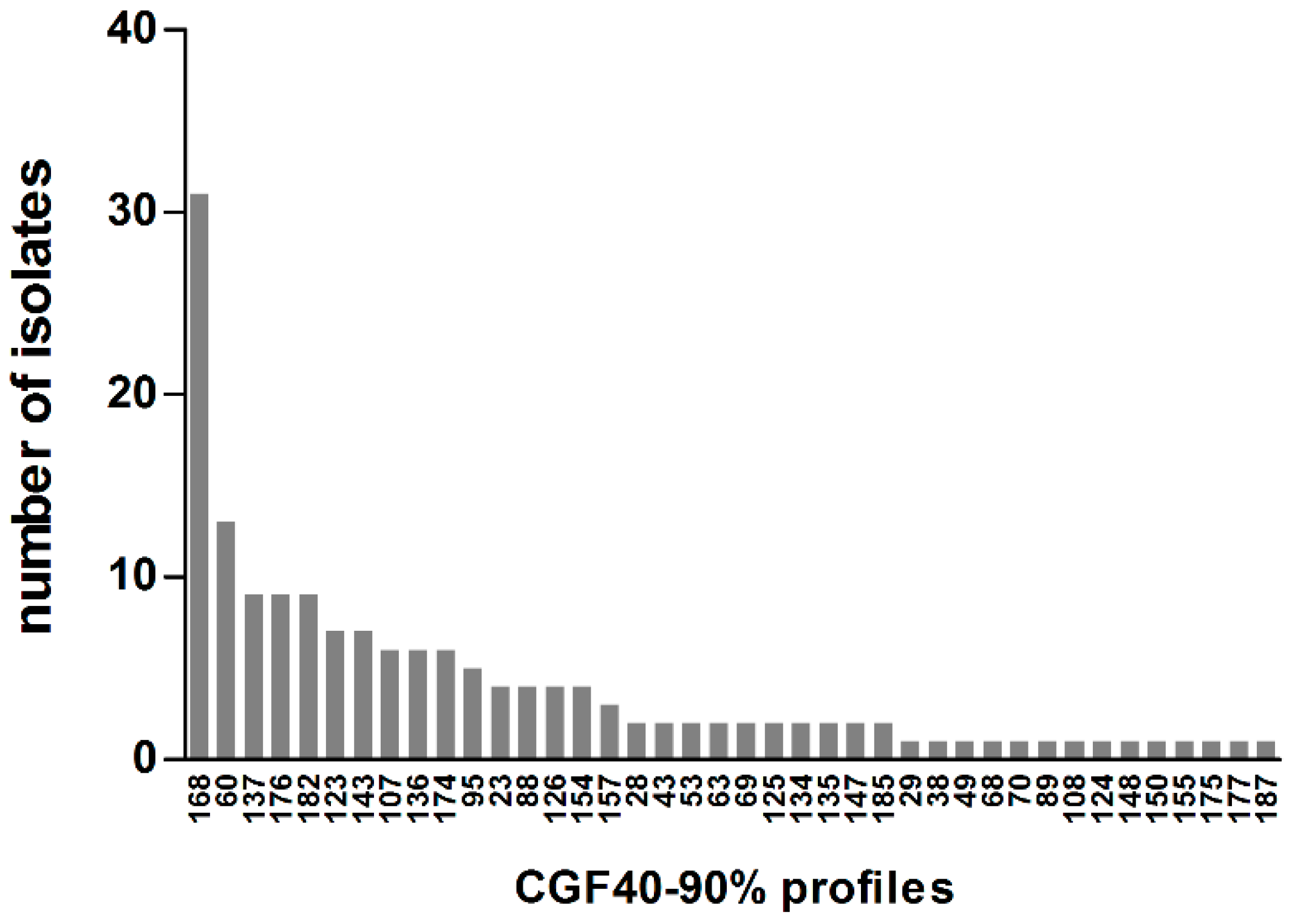
To compare C. jejuni isolates from pets with previously characterized C. jejuni from chicken, cattle, or humans in France [17,18,22], CGF40 clusters were then defined on the basis of 90% of similarity and, thereafter, termed as CGF40-90% clusters. Within the 161 C. jejuni isolated from pets, 40 CGF40-90% clusters were obtained. By looking at the distribution of CGF40-90% clusters within the C. jejuni population from pets (Figure 4), five clusters, including each at least 5% of C. jejuni isolates, were observed and were defined as predominant.
These 5 predominant CGF40-90% clusters (clusters 168, 60, 137, 176, and 182) included 44.1% of C. jejuni isolates from pets, and the most prevalent cluster (168) included 19.3% of the isolates (Table 2). Three of the 5 predominant clusters in pets were found in C. jejuni chicken isolates, while 4 of them were also isolated from cattle and human (Table 2). The cluster 168 which is the most prevalent cluster in pets, was found in a small proportion in cattle (0.9%) and clinical cases (0.6%) but was absent within the population of chicken isolates. However, the second most prevalent cluster in pets (cluster 60) was detected in chicken, cattle isolates, as well as in clinical isolates, in which it constitutes the most prevalent CGF40–90% cluster with 13.2% of prevalence. In addition and to a lesser extent, C. jejuni from clusters 137 and 182 were isolated in chicken (6.2 and 3.3% of isolates, respectively), cattle (1.2 and 1.4% of isolates, respectively), and clinical cases (4.1 and 1.2% of isolates, respectively) (Table 2). Finally, no C. jejuni belonging to the cluster 176, which includes 5.6% of pet isolates, was collected within chicken, cattle, or human (Table 2).
4. Discussion
This study conducted in Brittany is the first study to our knowledge which assesses the carriage of thermotolerant Campylobacter spp. by pets in France and describes the genetic structure of C. jejuni isolates circulating in these animals. Here, we highlighted the frequent carriage of thermotolerant Campylobacter spp. in dogs (38%), as well as their isolation in cats (10%) to a lesser extent. Wide variations were observed in Campylobacter spp. prevalence in cats and dogs between several prevalence studies performed in other countries. Indeed, from 4.8 to 73% of dogs were reported to carried Campylobacter spp., while the prevalence ranged from 9.9 to 41.9% in cats [15,25,26,27,28,29].
Here, we also reported a higher Campylobacter carriage by dogs under 1-year-old (63.6%) compared with older dogs (30.5%), while no difference was observed in cats. This result emphasized the variation of dog colonization according to their age previously described in a few studies [25,30]. In addition, several epidemiological studies highlighted that young dogs are more likely to carry Campylobacter spp. [31], especially C. upsaliensis [15,27].
C. upsaliensis is widely described as the predominant Campylobacter species isolated from pets and especially dogs, while, in some studies, C. helveticus was described as the most prevalent Campylobacter species in cats [12,32,33,34]. However, in accordance with Giacomelli et al. [35] and Amar et al. [14], the species identification of Campylobacter spp. isolates revealed in this study the predominance of C. jejuni in dogs (24.4%) and cats (7.1%). In 18% of Campylobacter-positive dogs, co-infection with up to three different species of Campylobacter was also found in our study. In their study, Chaban et al. [34] reported a simultaneous carriage in healthy and diarrheic dogs with up to 7 and 12 Campylobacter species, respectively, while Parson, et al. [28] described simultaneous carriage with 2 different Campylobacter species in dogs.
However, it should be noted that since our sampling survey was conducted locally, and despite a high number of samples, our results may not be representative of the French pet population. In addition, several factors might have influenced Campylobacter carriage rate by pets in our study, such as technical differences in the protocol used for the detection of thermophilic Campylobacter spp. In our study, conducted at the scale of the administrative department of Côtes d’Armor in Brittany, the season of sampling did not appear to influence the Campylobacter carriage by pets. However, seasonal variations in the prevalence of Campylobacter spp. in pets have been previously described. Indeed, Carbonero et al. [31] described a seasonal difference in the prevalence of C. jejuni in dogs during the spring compared with winter, while Mohan et al. [36] reported in New-Zealand a higher prevalence of C. jejuni in dogs during warmer months of the year. However, this seasonal variation is debatable since other studies did not observe significant seasonal variations in Campylobacter spp. prevalence in dogs [32,37]. Another factor, which might have impacted the carriage rate of Campylobacter within pets, is their health status, even if this is debatable since contradictory results are described. In fact, Carbonero et al. [31] showed that diarrhea in pets and especially dogs are positively associated with Campylobacter spp., C. jejuni and C. upsaliensis carriage, while Rossi et al. [33] and Parsons et al. [27] did not observe any association between intestinal disorder and Campylobacter spp. carriage rates in pets. The impact of health status on Campylobacter carriage in dogs and cats cannot be assessed in our study because of the low number of animals showing diarrhea symptoms (4/304). Finally, the living conditions of pets might also have influenced Campylobacter spp. carriage. In fact, dogs having frequent contact with birds, or outdoor cats are more likely to carry Campylobacter spp. [15], as well as stray dogs compared with family dogs [38]. This may also be the consequences of a higher exposure of these pets to Campylobacter spp. through the environment or contact with birds, which are well described to frequently carry Campylobacter species [39,40]. In our study, performed at the scale of the administrative department of Côtes d’Armor in Brittany, the living conditions of pets (inside or outside the house or both) did not appear to influence the Campylobacter carriage by pets.
Pets could therefore constitute a reservoir for Campylobacter spp. and especially for C. jejuni in France. Genotyping of C. jejuni isolates circulating in pets using CGF40 revealed in this study an important genetic diversity, with 79.5% of C. jejuni isolates belonging to a CGF40-100% profile found only once in the population. The carriage of highly diverse C. jejuni by pets in our study was also highlighted by the isolation of several genotypes (from 1 to 10) of C. jejuni within the same individual, as previously described by Koene et al. [41] using another genotyping method. This high genetic diversity in C. jejuni isolates was also highlighted during a longitudinal study by Hald et al. [32], in which different genotypes were isolated in the same animal over the time.
Within the five main “pet CGF40-90% genotypes”, three and four of them were also isolated in chicken and cattle, respectively. This finding is in accordance with Acke et al. [42], who reported an overlap of genotypes between pets and, chicken or cattle C. jejuni isolates, and may indicate a potential transmission of C. jejuni between these animals and pets. However, the absence, or the low isolation rate, of the first and fourth main “pet CGF40-90% genotypes” in cattle and chicken may suggest the existence of other sources of pet contamination by C. jejuni (e.g., environmental water or wild birds) or the existence of genotypes specific to C. jejuni from pets, therefore implying that they may constitute a reservoir for Campylobacter spp. with pet-specific genotypes. This absence of pet genotypes in cattle or chicken may also be the result of rapid genetic rearrangements in the genome of Campylobacter hosted by pets, as hypothesized by Koene et al. [41], to explain the shedding of distinct Campylobacter genotypes by pets living in the same household.
Finally, an overlap of CGF40-90% genotypes was observed in pets and human isolates, as previously described using other genotyping methods [12,14,43], suggesting a potential role of pets as a source of Campylobacter exposure for humans. This was especially supported by the identification of some risk factors for human contamination by Campylobacter spp. related to pets, such as owning a dog under 1-year-old [12]. As suggested by Mughini-Gras et al. [12], the transmission to humans of Campylobacter carried by pets may occur either directly by contact or by the household and the immediate environment contamination since these animals may eventually act as reservoirs for Campylobacter. However, the overlap between genotypes in pets and humans may also indicate a common infection of pets and humans by another source, such as cattle or chicken, which also overlapped with humans and pets, or a potential role of human as a source of pets contamination by Campylobacter spp. [12].
5. Conclusions
Pets constitute an important reservoir for Campylobacter spp. in our sampling area, and C. jejuni was the most frequent species isolated, while co-infections of individual animals with several species of Campylobacter were reported. Dogs appeared to carry more Campylobacter spp. than cats, as well as young dogs compared with dogs older than 1 year. The genetic characterization of C. jejuni isolates using CGF40 highlighted the high genetic diversity within the isolates population and the co-infection of individual animals by several genotypes of C. jejuni. Then, the overlap between pet and chicken or cattle isolates indicated the potential role of the livestock as a source of contamination for pets or inversely, while the overlap between isolates from pets and humans suggested a potential role of pets in human contamination by C. jejuni, or vice versa. Because of low attribution of human campylobacteriosis to pets in source attribution studies, the risk for human health is considered low. Nevertheless, because of the proximity between pets and humans, and the frequent carriage of C. jejuni by these animals in our study, the exposure of human to Campylobacter from pets can be more important than thought, and pets can constitute a significant source of human exposure to Campylobacter. Further analyses will be necessary to assess with accuracy the implication of pets in human campylobacteriosis.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2615/10/5/838/s1, Table S1: Genetic characteristics of C. jejuni isolates collected from pets in France.
Author Contributions
Conceptualization, M.C., K.R., A.T.; methodology, M.C., K.R., A.T.; investigation, A.T., V.R., M.Q.; writing—original draft preparation, A.T.; writing—review and editing, M.C., K.R., A.T.; supervision, M.C. and K.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the French Ministry of Defence (Direction Générale de l’Armement) and the Conseil Général des Côtes d’Armor (CG22).
Acknowledgments
Authors would like to thank veterinarians, private owners and breeders who provided samples during the 6-month period of the study. We thank the Maldi-Tof platform of the Anses Laboratory of Nancy for the reading of the protein extracts
Conflicts of Interest
The authors declare no conflict of interest.
References
- EFSA. Scientific report on the European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 17. [Google Scholar] [CrossRef] [Green Version]
- Bessède, E.; Lehours, P.; Labadi, L.; Bakiri, S.; Mégraud, F. Comparison of Characteristics of Patients Infected by Campylobacter jejuni, Campylobacter coli, and Campylobacter fetus. J. Clin. Microbiol. 2013, 52, 328–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, A.R.; Pires, S.M.; Halasa, T.; Hald, T. Source attribution of human campylobacteriosis using a meta-analysis of case-control studies of sporadic infections. Epidemiol. Infect. 2012, 140, 970–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, S.; Fournier, E.; Carrier, N.; Frost, E.; Arbeit, R.D.; Michaud, S. Campylobacteriosis in Urban versus Rural Areas: A Case-Case Study Integrated with Molecular Typing to Validate Risk Factors and to Attribute Sources of Infection. PLoS ONE 2013, 8, e83731. [Google Scholar] [CrossRef] [Green Version]
- Mossong, J.; Mughini-Gras, L.; Penny, C.; Devaux, A.; Olinger, C.; Losch, S.; Cauchie, H.-M.; Van Pelt, W.; Ragimbeau, C. Human Campylobacteriosis in Luxembourg, 2010–2013: A Case-Control Study Combined with Multilocus Sequence Typing for Source Attribution and Risk Factor Analysis. Sci. Rep. 2016, 6, 20939. [Google Scholar] [CrossRef] [Green Version]
- Gras, L.M.; Smid, J.H.; Wagenaar, J.A.; De Boer, A.G.; Havelaar, A.H.; Friesema, I.; French, N.P.; Busani, L.; Van Pelt, W. Risk Factors for Campylobacteriosis of Chicken, Ruminant, and Environmental Origin: A Combined Case-Control and Source Attribution Analysis. PLoS ONE 2012, 7, e42599. [Google Scholar] [CrossRef] [Green Version]
- Berthenet, E.; Thépault, A.; Chemaly, M.; Rivoal, K.; Ducournau, A.; Buissonnière, A.; Bénéjat, L.; Bessède, E.; Mégraud, F.; Sheppard, S.K.; et al. Source attribution of Campylobacter jejuni shows variable importance of chicken and ruminants reservoirs in non-invasive and invasive French clinical isolates. Sci. Rep. 2019, 9, 8098. [Google Scholar] [CrossRef]
- Thépault, A.; Méric, G.; Rivoal, K.; Pascoe, B.; Mageiros, L.; Touzain, F.; Rose, V.; Béven, V.; Chemaly, M.; Sheppard, S.K. Genome-Wide Identification of Host-Segregating Epidemiological Markers for Source Attribution in Campylobacter jejuni. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Thépault, A.; Rose, V.; Quesne, S.; Poëzevara, T.; Beven, V.; Hirchaud, E.; Touzain, F.; Lucas, P.; Méric, G.; Mageiros, L.; et al. Ruminant and chicken: Important sources of campylobacteriosis in France despite a variation of source attribution in 2009 and 2015. Sci. Rep. 2018, 8, 9305. [Google Scholar] [CrossRef]
- Sheppard, S.K.; Dallas, J.F.; Strachan, N.J.C.; Macrae, M.; McCarthy, N.; Wilson, D.J.; Gormley, F.J.; Falush, D.; Ogden, I.D.; Maiden, M.C.J.; et al. Campylobacter Genotyping to Determine the Source of Human Infection. Clin. Infect. Dis. 2009, 48, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Kittl, S.; Heckel, G.; Korczak, B.M.; Kuhnert, P. Source Attribution of Human Campylobacter Isolates by MLST and Fla-Typing and Association of Genotypes with Quinolone Resistance. PLoS ONE 2013, 8, e81796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gras, L.M.; Smid, J.H.; Wagenaar, J.A.; Koene, M.G.J.; Havelaar, A.H.; Friesema, I.; French, N.P.; Flemming, C.; Galson, J.; Graziani, C.; et al. Increased risk for Campylobacter jejuni and C. coli infection of pet origin in dog owners and evidence for genetic association between strains causing infection in humans and their pets. Epidemiol. Infect. 2013, 141, 2526–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, B.M.; Schielke, A.; Didelot, X.; Kops, F.; Breidenbach, J.; Willrich, N.; Gölz, G.; Alter, T.; Stingl, K.; Josenhans, C.; et al. A combined case-control and molecular source attribution study of human Campylobacter infections in Germany, 2011–2014. Sci. Rep. 2017, 7, 5139. [Google Scholar] [CrossRef] [PubMed]
- Amar, C.; Kittl, S.; Spreng, D.; Thomann, A.; Korczak, B.M.; Burnens, A.P.; Kuhnert, P. Genotypes and antibiotic resistance of canine Campylobacter jejuni isolates. Vet. Microbiol. 2014, 168, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Wieland, B.; Schüpbach-Regula, G.; Danuser, J.; Wittwer, M.; Burnens, A.P.; Wassenaar, T.M.; Stärk, K.D.C. Campylobacter spp. in Dogs and Cats in Switzerland: Risk Factor Analysis and Molecular Characterization with AFLP. J. Vet. Med. Ser. B 2005, 52, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Châtre, P.; Haenni, M.; Meunier, D.; Botrel, M.-A.; Calavas, D.; Madec, J.-Y. Prevalence and Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli Isolated from Cattle between 2002 and 2006 in France. J. Food Prot. 2010, 73, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Thépault, A.; Guyard-Nicodème, M.; Rose, V.; Quesne, S.; Queguiner, M.; Houard, E.; Mégraud, F.; Rivoal, K.; Chemaly, M. A representative overview of the genetic diversity and lipooligosaccharide sialylation in Campylobacter jejuni along the broiler production chain in France and its comparison with human isolates. Int. J. Food Microbiol. 2018, 274, 20–30. [Google Scholar] [CrossRef]
- Thepault, A.; Poezevara, T.; Quesne, S.; Rose, V.; Chemaly, M.; Rivoal, K. Prevalence of Thermophilic Campylobacter in Cattle Production at Slaughterhouse Level in France and Link Between C. jejuni Bovine Strains and Campylobacteriosis. Front. Microbiol. 2018, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Denis, M.; Tanguy, M.; Chidaine, B.; Laisney, M.-J.; Mégraud, F.; Fravalo, P. Description and sources of contamination by Campylobacter spp. of river water destined for human consumption in Brittany, France. Pathol. Boil. 2011, 59, 256–263. [Google Scholar] [CrossRef]
- Taboada, E.N.; Ross, S.L.; Mutschall, S.K.; MacKinnon, J.M.; Roberts, M.J.; Buchanan, C.J.; Kruczkiewicz, P.; Jokinen, C.C.; Thomas, J.E.; Nash, J.; et al. Development and Validation of a Comparative Genomic Fingerprinting Method for High-Resolution Genotyping of Campylobacter jejuni. J. Clin. Microbiol. 2011, 50, 788–797. [Google Scholar] [CrossRef] [Green Version]
- Dingle, K.; Colles, F.M.; Wareing, D.; Ure, R.; Fox, A.J.; Bolton, F.E.; Bootsma, H.J.; Willems, R.J.L.; Urwin, R.; Maiden, M.C.J. Multilocus Sequence Typing System for Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Rose, V.; Thépault, A.; Lemercier, A.; Mégraud, F.; Chemaly, M.; Rivoal, K. A stable and highly diverse genetic structure in clinical Campylobacter jejuni isolates in 2009 and 2015. In Proceedings of the 19th International Workshop on Campylobacter, Helicobacter and Related Organisms (CHRO), Nantes, France, 10–14 September 2017; p. 148. [Google Scholar]
- Carriço, J.A.; Silva-Costa, C.; Cristino, J.M.; Pinto, F.R.; De Lencastre, H.; Almeida, J.S.; Ramirez, M. Illustration of a Common Framework for Relating Multiple Typing Methods by Application to Macrolide-Resistant Streptococcus pyogenes†. J. Clin. Microbiol. 2006, 44, 2524–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severiano, A.; Carriço, J.A.; Robinson, D.A.; Ramirez, M.; Pinto, F.R. Evaluation of Jackknife and Bootstrap for Defining Confidence Intervals for Pairwise Agreement Measures. PLoS ONE 2011, 6, e19539. [Google Scholar] [CrossRef] [PubMed]
- Acke, E.; McGill, K.; Golden, O.; Jones, B.R.; Fanning, S.; Whyte, P. Prevalence of thermophilic Campylobacter species in household cats and dogs in Ireland. Vet. Rec. 2009, 164, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewska, M.; Szczepańska, B.; Klawe, J.; Śpica, D.; Chudzińska, M. Prevalence of Campylobacter jejuni and Campylobacter coli species in cats and dogs from Bydgoszcz (Poland) region. Pol. J. Vet. Sci. 2013, 16, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, B.; Porter, C.; Ryvar, R.; Stavisky, J.; Williams, N.; Pinchbeck, G.; Birtles, R.; Christley, R.; German, A.J.; Radford, A.D.; et al. Prevalence of Campylobacter spp. in a cross-sectional study of dogs attending veterinary practices in the UK and risk indicators associated with shedding. Vet. J. 2010, 184, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Parsons, B.; Williams, N.; Pinchbeck, G.; Christley, R.; Hart, C.; Gaskell, R.; Dawson, S. Prevalence and shedding patterns of Campylobacter spp. in longitudinal studies of kennelled dogs. Vet. J. 2011, 190, 249–254. [Google Scholar] [CrossRef]
- Torkan, S.; Vazirian, B.; Khamesipour, F.; Dida, G.O. Prevalence of thermotolerantCampylobacterspecies in dogs and cats in Iran. Vet. Med. Sci. 2018, 4, 296–303. [Google Scholar] [CrossRef]
- Rahimi, E.; Chakeri, A.; Esmizadeh, K. Prevalence of Campylobacter species in fecal samples from cats and dogs in Iran. Slov. Vet. Res. 2012, 49, 117–122. [Google Scholar]
- Carbonero, A.; Torralbo, A.; Borge, C.; García-Bocanegra, I.; Arenas, A.; Perea, A. Campylobacter spp., C. jejuni and C. upsaliensis infection-associated factors in healthy and ill dogs from clinics in Cordoba, Spain. Screening tests for antimicrobial susceptibility. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 505–512. [Google Scholar] [CrossRef]
- Hald, B.; Pedersen, K.; Wainø, M.; Jørgensen, J.C.; Madsen, M. Longitudinal study of the excretion patterns of thermophilic Campylobacter spp. in young pet dogs in Denmark. J. Clin. Microbiol. 2004, 42, 2003–2012. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Hänninen, M.-L.; Revez, J.; Hannula, M.; Zanoni, R. Occurrence and species level diagnostics of Campylobacter spp., enteric Helicobacter spp. and Anaerobiospirillum spp. in healthy and diarrheic dogs and cats. Vet. Microbiol. 2008, 129, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Chaban, B.; Ngeleka, M.; Hill, J.E. Detection and quantification of 14 Campylobacter species in pet dogs reveals an increase in species richness in feces of diarrheic animals. BMC Microbiol. 2010, 10, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomelli, M.; Follador, N.; Coppola, L.; Martini, M.; Piccirillo, A. Survey of Campylobacter spp. in owned and unowned dogs and cats in Northern Italy. Vet. J. 2015, 204, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Mohan, V.; Stevenson, M.; Marshall, J.; French, N.P.; Marshall, J. Characterisation by multilocus sequence and por A and fla A typing of Campylobacter jejuni isolated from samples of dog faeces collected in one city in New Zealand. N. Z. Vet. J. 2017, 65, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, M.; Bergsjø, B.; Hofshagen, M.; Skjerve, E.; Kruse, H. Risk factors for Campylobacter infection in Norwegian cats and dogs. Prev. Vet. Med. 2002, 55, 241–253. [Google Scholar] [CrossRef]
- Tsai, H.-J.; Huang, H.-C.; Lin, C.-M.; Lien, Y.-Y.; Chou, C.-H. Salmonellae and Campylobacters in Household and Stray Dogs in Northern Taiwan. Vet. Res. Commun. 2007, 31, 931–939. [Google Scholar] [CrossRef]
- Mohan, V. Faeco-prevalence of Campylobacter jejuni in urban wild birds and pets in New Zealand. BMC Res. Notes 2015, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Waldenström, J.; Broman, T.; Carlsson, I.; Hasselquist, D.; Achterberg, R.P.; Wagenaar, J.A.; Olsen, B. Prevalence of Campylobacter jejuni, Campylobacter lari, and Campylobacter coli in Different Ecological Guilds and Taxa of Migrating Birds. Appl. Environ. Microbiol. 2002, 68, 5911–5917. [Google Scholar] [CrossRef] [Green Version]
- Koene, M.G.; Houwers, D.J.; Dijkstra, J.R.; Duim, B.; Wagenaar, J.A. Strain variation within Campylobacter species in fecal samples from dogs and cats. Vet. Microbiol. 2009, 133, 199–205. [Google Scholar] [CrossRef]
- Acke, E.; Carroll, C.; O’Leary, A.; McGill, K.; Kelly, L.; Lawlor, A.; Madden, R.H.; Moran, L.; Scates, P.; McNamara, E.; et al. Genotypic characterisation and cluster analysis of Campylobacter jejuni isolates from domestic pets, human clinical cases and retail food. Ir. Vet. J. 2011, 64, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damborg, P.; Olsen, K.E.P.; Nielsen, E.M.; Guardabassi, L. Occurrence of Campylobacter jejuni in Pets Living with Human Patients Infected with C. jejuni. J. Clin. Microbiol. 2004, 42, 1363–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Carriage of thermophilic Campylobacter spp. in pets from France. * represents the significant difference between Campylobacter spp. carriage by dogs and cats (Fisher test, p < 0.0001); ** represents the significant difference between Campylobacter spp. carriage by dogs ≤1-year-old and older dogs (Fisher test, p < 0.0001).
Figure 1.
Carriage of thermophilic Campylobacter spp. in pets from France. * represents the significant difference between Campylobacter spp. carriage by dogs and cats (Fisher test, p < 0.0001); ** represents the significant difference between Campylobacter spp. carriage by dogs ≤1-year-old and older dogs (Fisher test, p < 0.0001).

Figure 2.
Venn diagram representing thermophilic Campylobacter spp. carried in pets (dogs and cats) from France. Numbers represent the number of pets carrying Campylobacter species. When an overlap is observed between two or more circles representing Campylobacter species, it means a co-infection of pets by these Campylobacter species.
Figure 2.
Venn diagram representing thermophilic Campylobacter spp. carried in pets (dogs and cats) from France. Numbers represent the number of pets carrying Campylobacter species. When an overlap is observed between two or more circles representing Campylobacter species, it means a co-infection of pets by these Campylobacter species.

Figure 3.
Distribution of CGF40-100% profiles within 161 C. jejuni isolates from cats and dogs in France. Labels on the X-axis correspond to the CGF40-100% cluster number.
Figure 3.
Distribution of CGF40-100% profiles within 161 C. jejuni isolates from cats and dogs in France. Labels on the X-axis correspond to the CGF40-100% cluster number.

Figure 4.
Distribution of CGF40-90% profiles within 161 C. jejuni isolates from cats and dogs in France. Labels on the X-axis correspond to the CGF40-90% cluster number.
Figure 4.
Distribution of CGF40-90% profiles within 161 C. jejuni isolates from cats and dogs in France. Labels on the X-axis correspond to the CGF40-90% cluster number.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Prevalence of thermophilic Campylobacter species from dogs and cats feces samples in France.
Table 1.
Prevalence of thermophilic Campylobacter species from dogs and cats feces samples in France.
| Bacterial Species Isolated | Dogs | Cats | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total (n = 234) | ≤1 year (n = 55) | >1 year ( n= 174) | N/A (n = 5) | Total (n = 70) | ≤1 year (n = 26) | >1 year (n = 42) | N/A (n = 2) | |||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | |
| Campylobacter spp. | 89 | 38.0 * | 35 | 63.6 ** | 53 | 30.5 | 1 | 20.0 | 7 | 10.0 | 1 | 3.8 | 6 | 14.3 | 0 | 0 |
| C. jejuni | 57 | 24.4 | 24 | 43.6 | 32 | 18.4 | 1 | 20.0 | 5 | 7.1 | 1 | 3.8 | 4 | 9.5 | 0 | 0 |
| C. coli | 6 | 2.6 | 4 | 7.3 | 2 | 1.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C. lari | 26 | 11.1 | 12 | 21.8 | 14 | 8.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C. uspaliensis | 21 | 9.0 | 8 | 14.5 | 13 | 7.5 | 0 | 0 | 1 | 1.4 | 0 | 0 | 1 | 2.4 | 0 | 0 |
| Campylobacter species not identified | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.4 | 0 | 0 | 1 | 2.4 | 0 | 0 |
* Significantly higher prevalence of Campylobacter spp. in dogs compared with cats (Fisher test, p < 0.001). ** Significantly higher prevalence of Campylobacter spp. in young dogs compared with older dogs (Fisher test, p < 0.001). Note: Co-infection with two or three Campylobacter species was observed within 16 dogs.
Table 2.
Presence of the 5 main CGF40-90% clusters of C. jejuni from pets, among chicken, cattle, or human isolates in France.
Table 2.
Presence of the 5 main CGF40-90% clusters of C. jejuni from pets, among chicken, cattle, or human isolates in France.
| CGF40-90% Cluster | Pets | Chicken | Cattle | Human |
|---|---|---|---|---|
| 168 | 31/161 (19.3%) | 0/644 (0%) | 6/649 (0.9%) | 3/514 (0.6%) |
| 60 | 13/161 (8.1%) | 51/644 (8.0%) | 66/649 (10.2%) | 68/514 (13.2%) |
| 137 | 9/161 (5.6%) | 40/644 (6.2%) | 8/649 (1.2%) | 21/514 (4.1%) |
| 176 | 9/161 (5.6%) | 0/644 (0%) | 0/649 (0%) | 0/514 (0%) |
| 182 | 9/161 (5.6%) | 21/644 (3.3%) | 9/649 (1.4%) | 6/514 (1.2%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thépault, A.; Rose, V.; Queguiner, M.; Chemaly, M.; Rivoal, K. Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure. Animals 2020, 10, 838. https://doi.org/10.3390/ani10050838
AMA Style
Thépault A, Rose V, Queguiner M, Chemaly M, Rivoal K. Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure. Animals. 2020; 10(5):838. https://doi.org/10.3390/ani10050838
Chicago/Turabian StyleThépault, Amandine, Valérie Rose, Marilyne Queguiner, Marianne Chemaly, and Katell Rivoal. 2020. "Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure" Animals 10, no. 5: 838. https://doi.org/10.3390/ani10050838
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.