Cyclospora cayetanensis and Cyclosporiasis: An Update



Abstract
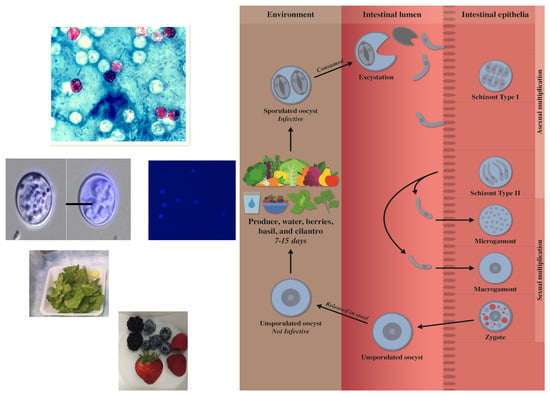
1. Introduction
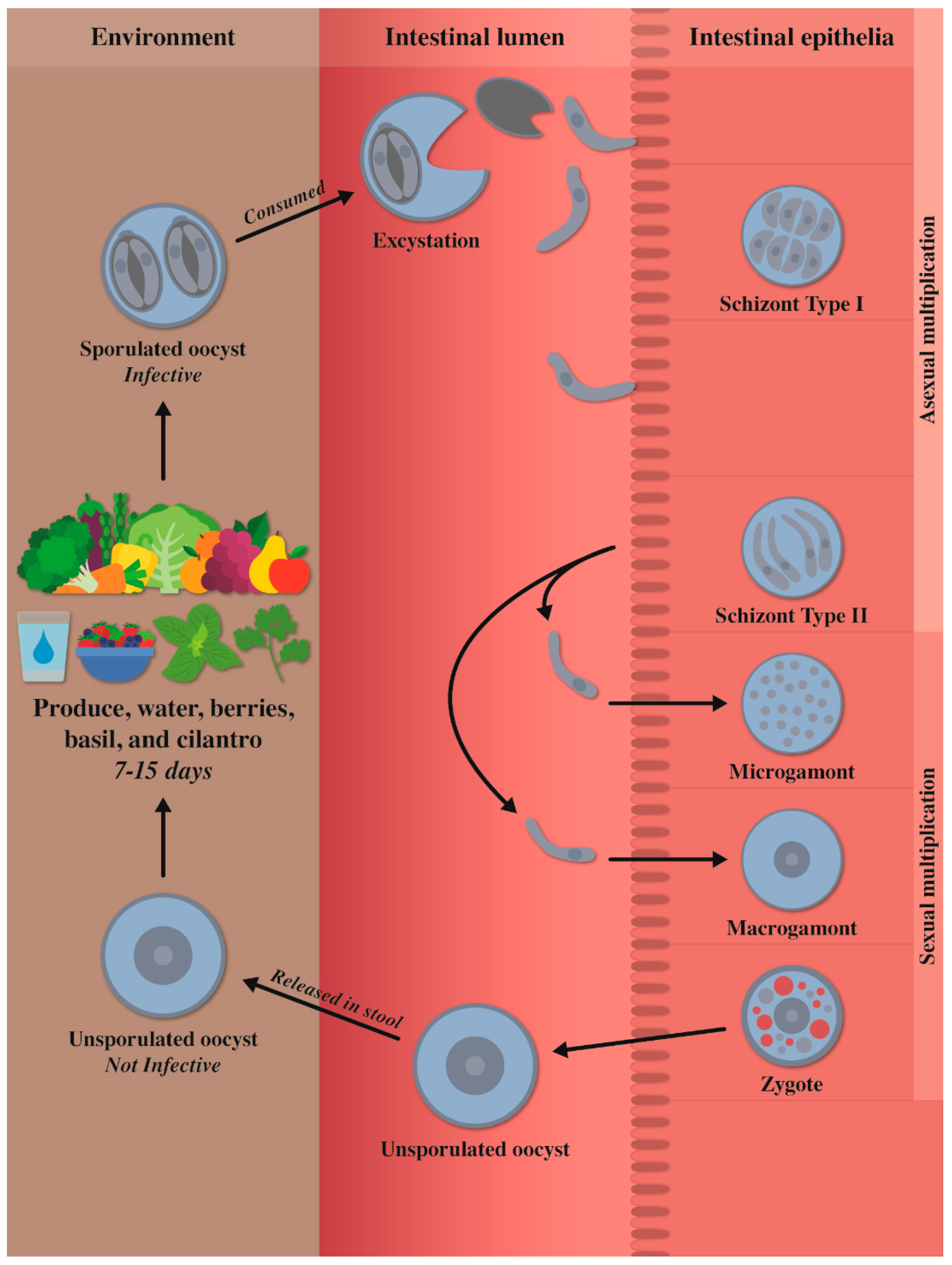
2. Morphology and Life Cycle
Oocyst Sporulation
3. Transmission and Epidemiology
3.1. Seasonality
3.2. Risk Factors
3.2.1. Contact with Animals
3.2.2. Age
3.2.3. Gender
3.2.4. Socioeconomic Status
3.2.5. Immunocompromised Hosts
3.2.6. Resident Foreigners, Expatriates and Traveler’s Diarrhea
3.2.7. Consumption of Contaminated Food
3.2.8. Waterborne Infection of C. cayetanensis
3.2.9. Soil as a Source of Infection and/or Transmission for C. cayetanensis Oocysts
4. Outbreaks Due to C. cayetanensis
5. Clinical Symptoms and Pathogenesis
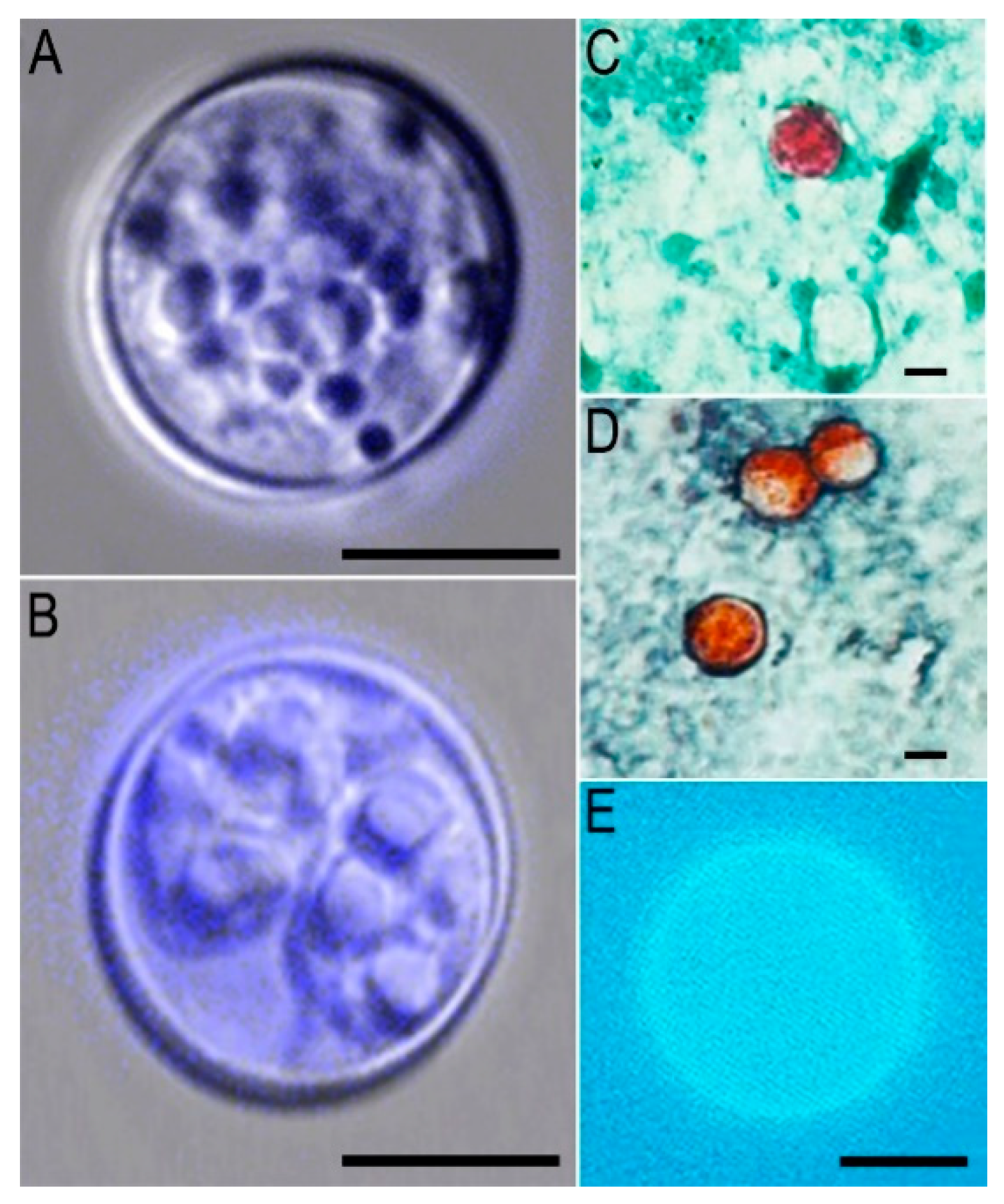
6. Diagnosis
6.1. Serological Studies
6.2. Oocyst Detection in Clinical Samples
6.2.1. Oocyst Testing
6.2.2. Modified Acid-Fast Staining
6.2.3. Heated Safranin Staining
6.2.4. Auto Fluorescence
6.3. Molecular Detection and Characterization
7. Treatment
8. Control and Prevention
9. Conclusions and Future Research Needed
Funding
Acknowledgments
Conflicts of Interest
Data availability
References
- Ortega, Y.R.; Sterling, C.R.; Gilman, R.H.; Cama, V.A.; Díaz, F. Cyclospora species: A new protozoan pathogen of humans. N. Engl. J. Med. 1993, 328, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Gilman, R.H.; Sterling, C.R. A new coccidian parasite (Apicomplexa: Eimeriidae) from humans. J. Parasitol. 1994, 80, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Sanchez, R. Update on Cyclospora cayetanensis, a food-borne and waterborne parasite. Clin. Microbiol. Rev. 2010, 23, 218–234. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.V.; Paton, C.A.; Girdwood, R.W.A.; Mtambo, M.M.A. Cyclospora in non-human primates in Gombe, Tanzania. Vet. Rec. 1996, 138, 528. [Google Scholar] [PubMed]
- Lopez, F.A.; Manglicmot, J.; Schmidt, T.M.; Yeh, C.; Smith, H.V.; Relman, D.A. Molecular characterization of Cyclospora-like organisms from baboons. J. Infect. Dis. 1999, 179, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; da Silva, A.J.; Lilley, B.G.; Pieniazek, N.J. Morphologic and molecular characterization of new Cyclospora species from Ethiopian monkeys: C. cercopitheci sp.n., C. colobi sp.n., and C. papionis sp.n. Emerg. Infect. Dis. 1999, 5, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Njenga, M.N.; DaSilva, A.J.; Owino, D.; Nace, E.K.; Won, K.Y.; Mwenda, J.M. A survey for Cyclospora spp. in Kenyan primates, with some notes on its biology. J. Parasitol. 2001, 87, 1394–1397. [Google Scholar] [CrossRef]
- Zhao, G.H.; Cong, M.M.; Bian, Q.Q.; Cheng, W.Y.; Wang, R.J.; Qi, M.; Zhang, L.X.; Lin, Q.; Zhu, X.Q. Molecular characterization of Cyclospora-like organisms from golden snub-nosed monkeys in Qinling Mountain in Shaanxi province, northwestern China. PLoS ONE 2013, 8, e58216. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Owens, J.R.; Bishop, H.S.; de Almeida, M.E.; da Silva, A.J.; Hearn, G.; Honarvar, S. Cyclospora spp. in drills, Bioko Island, Equatorial Guinea. Emerg. Infect. Dis. 2014, 20, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ye, J.; Arrowood, M.J.; Ma, J.; Wang, L.; Xu, H.; Feng, Y.; Xiao, L. Identification and morphologic and molecular characterization of Cyclospora macacae n. sp. from rhesus monkeys in China. Parasitol. Res. 2015, 114, 1811–1816. [Google Scholar] [CrossRef][Green Version]
- Marangi, M.; Koehler, A.V.; Zanzani, S.A.; Manfredi, M.T.; Brianti, E.; Giangaspero, A.; Gasser, R.B. Detection of Cyclospora in captive chimpanzees and macaques by a quantitative PCR-based mutation scanning approach. Parasites Vectors 2015, 8, 274. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Ortega, Y.R.; Hanes, D.E.; Nace, E.K.; Do, R.Q.; Robl, M.G.; Won, K.Y.; Gavidia, C.; Sass, N.L.; Mansfield, K. Attempts to establish experimental Cyclospora cayetanensis infection in laboratory animals. J. Parasitol. 2000, 86, 577–582. [Google Scholar] [CrossRef]
- Zerpa, R.; Uchima, N.; Huicho, L. Cyclospora cayetanensis associated with watery diarrhoea in Peruvian patients. J. Trop. Med. Hyg. 1995, 98, 325–329. [Google Scholar] [PubMed]
- García-López, H.L.; Rodríguez-Tovar, L.E.; Medina-De la Garza, C.E. Identification of Cyclospora in poultry. Emerg. Infect. Dis. 1996, 2, 356–357. [Google Scholar] [CrossRef] [PubMed]
- Sherchand, J.B.; Cross, J.H. Emerging pathogen Cyclospora cayetanensis infection in Nepal. Southeast Asian J. Trop. Med. Public Health 2001, 32 (Suppl. 2), 143–150. [Google Scholar] [PubMed]
- Pérez Cordón, G.; Hitos Prados, A.; Romero, D.; Sánchez Moreno, M.; Pontes, A.; Osuna, A.; Rosales, M.J. Intestinal parasitism in the animals of the zoological garden “Peña Escrita” (Almuñecar, Spain). Vet. Parasitol. 2008, 156, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Pérez Cordón, G.; Hitos Prados, A.; Romero, D.; Sánchez Moreno, M.; Pontes, A.; Osuna, A.; Rosales, M.J. Intestinal and haematic parasitism in the birds of the Almuñecar (Granada, Spain) ornithological garden. Vet. Parasitol. 2009, 165, 361–366. [Google Scholar] [CrossRef]
- Cama, V.A.; Ortega, Y.R. Cyclospora cayetanensis. In Foodborne Parasites, 2nd ed.; Ortega, Y.R., Sterling, C.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 41–56. [Google Scholar]
- Ortega, Y.R.; Sherchand, J.B. Cyclospora cayetanensis. In Biology of Foodborne Parasites, 1st ed.; Xiao, L., Ryan, U., Feng, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 97–110. [Google Scholar]
- Li, G.; Xiao, S.; Zhou, R.; Li, W.; Wadeh, H. Molecular characterization of Cyclospora-like organism from dairy cattle. Parasitol. Res. 2007, 100, 955–961. [Google Scholar] [CrossRef]
- Chu, D.M.T.; Sherchand, J.B.; Cross, J.H.; Orlandi, P.A. Detection of Cyclospora cayetanensis in animal fecal isolates from Nepal using an FTA filter-base polymerase chain reaction method. Am. J. Trop. Med. Hyg. 2004, 71, 373–379. [Google Scholar] [CrossRef]
- Sun, T.; Ilardi, C.F.; Asnis, D.; Bresciani, A.R.; Goldenberg, S.; Roberts, B.; Teichberg, S. Light and electron microscopic identification of Cyclospora species in the small intestine. Evidence of the presence of asexual life cycle in human host. Am. J. Clin. Pathol. 1996, 105, 216–220. [Google Scholar] [CrossRef]
- Nhieu, J.T.V.; Nin, F.; Fleury-Feith, J.; Chaumette, M.T.; Schaeffer, A.; Bretagne, S. Identification of intracellular stages of Cyclospora species by light microscopy of thick sections using hematoxylin. Hum. Pathol. 1996, 27, 1107–1109. [Google Scholar] [PubMed]
- Ortega, Y.R.; Nagle, R.; Gilman, R.H.; Watanabe, J.; Miyagui, J.; Quispe, H.; Kanagusuku, P.; Roxas, C.; Sterling, C.R. Pathologic and clinical findings in patients with cyclosporiasis and a description of intracellular parasite life-cycle stages. J. Infect. Dis. 1997, 176, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Connor, B.A.; Reidy, J.; Soave, R. Cyclosporiasis: Clinical and histopathologic correlates. Clin. Infect. Dis. 1999, 28, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Velásquez, J.N.; Carnevale, S.; Cabrera, M.; Kuo, L.; Chertcoff, A.; Mariano, M.; Bozzini, J.P.; Etchart, C.; Argento, R.; Di Risio, C. Cyclospora cayetanensis en pacientes con SIDA y diarrea crónica. Acta Gastroenterol. Lat. 2004, 34, 133–137. [Google Scholar]
- Tsang, O.T.Y.; Wong, R.W.C.; Lam, B.H.S.; Chan, J.M.C.; Tsang, K.Y.; Leung, W.S. Cyclospora infection in a young woman with human immunodeficiency virus in Hong Kong: A case report. BMC Res. Notes 2013, 6, 521. [Google Scholar] [CrossRef] [PubMed]
- Di Gliullo, A.B.; Cribari, M.S.; Bava, A.J.; Cicconetti, J.S.; Collazos, R. Cyclospora cayetanensis in sputum and stool samples. Rev. Inst. Med. Trop. São Paulo 2000, 42, 115–117. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hussein, E.M.; Abdul-Manaem, A.H.; El-Attary, E.S.L. Cyclospora cayetanensis oocysts in sputum of a patient with active pulmonary tuberculosis, case report in Ismailia, Egypt. J. Egypt. Soc. Parasitol. 2005, 35, 787–793. [Google Scholar] [PubMed]
- Sifuentes-Osornio, J.; Porras-Cortés, G.; Bendall, R.P.; Morales-Villarreal, F.; Reyes-Terán, G.; Ruiz-Palacios, G.M. Cyclospora cayetanensis infection in patients with and without AIDS: Biliary disease as another clinical manifestation. Clin. Infect. Dis. 1995, 21, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- De Górgolas, M.; Fortés, J.; Fernández Guerrero, M.L. Cyclospora cayetanensis cholecystitis in a patient with AIDS. Ann. Intern. Med. 2001, 134, 166. [Google Scholar] [CrossRef] [PubMed]
- Zar, F.A.; El-Bayoumi, E.; Yungbluth, M.M. Histologic proof of acalculous cholecystitis due to Cyclospora cayetanensis. Clin. Infect. Dis. 2001, 33, E140–E141. [Google Scholar] [CrossRef][Green Version]
- Smith, H.V.; Paton, C.A.; Mtambo, M.M.A.; Girdwood, R.W.A. Sporulation of Cyclospora sp. oocysts. Appl. Environ. Microbiol. 1997, 63, 1631–1632. [Google Scholar] [PubMed]
- Sathyanarayanan, L.; Ortega, Y. Effects of temperature and different food matrices on Cyclospora cayetanensis oocyst sporulation. J. Parasitol. 2006, 92, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Sathyanarayanan, L.; Ortega, Y. Effects of pesticides on sporulation of Cyclospora cayetanensis and viability of Cryptosporidium parvum. J. Food Prot. 2004, 67, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Sterling, C.R.; Gilman, R.H. Cyclospora cayetanensis. Adv. Parasitol. 1998, 40, 399–418. [Google Scholar] [CrossRef]
- Chacín-Bonilla, L. Transmission of Cyclospora cayetanensis infection: A review focusing on soil-borne cyclosporiasis. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 215–216. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Olson, B.H. Cyclospora cayetanensis: A review of an emerging parasitic coccidian. Int. J. Parasitol. 2003, 33, 371–391. [Google Scholar] [CrossRef]
- Chacin-Bonilla, L. Cyclospora cayetanensis. In Global Water Pathogens Project; (Fayer, R., Jakubowski, W., Eds. Part 3 Protists); Rose, J.B., Jiménez-Cisneros, B., Eds.; Michigan State University: East Lansing, MI, USA, 2017; Available online: http://www.waterpathogens.org/book/cyclospora-cayetanensis (accessed on 27 July 2019).
- Hoge, C.W.; Shlim, D.R.; Rajah, R.; Triplett, J.; Shear, M.; Rabold, J.G.; Echeverria, P. Epidemiology of diarrhoeal illness associated with coccidian-like organism among travellers and foreign residents in Nepal. Lancet 1993, 341, 1175–1179. [Google Scholar] [CrossRef]
- Pape, J.W.; Verdier, R.I.; Boncy, M.; Boncy, J.; Johnson, W.D., Jr. Cyclospora infection in adults infected with HIV. Clinical manifestations, treatment, and prophylaxis. Ann. Intern. Med. 1994, 121, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Fryauff, D.J.; Krippner, R.; Prodjodipuro, P.; Ewald, C.; Kawengian, S.; Pegelow, K.; Yun, T.; von Heydwolff-Wehnert, C.; Oyofo, B.; Gross, R. Cyclospora cayetanensis among expatriate and indigenous populations of West Java, Indonesia. Emerg. Infect. Dis. 1999, 5, 585–588. [Google Scholar] [CrossRef]
- Ooi, W.W.; Zimmerman, S.K.; Needham, C.A. Cyclospora species as a gastrointestinal pathogen in immunocompetent hosts. J. Clin. Microbiol. 1995, 33, 1267–1269. [Google Scholar]
- Wurtz, R. Cyclospora: A newly identified intestinal pathogen of humans. Clin. Infect. Dis. 1994, 18, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.C.; McIntyre, M. The incidence of Cyclospora cayetanensis in stool samples submitted to a district general hospital. Epidemiol. Infect. 1996, 117, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Chacín-Bonilla, L. Epidemiology of Cyclospora cayetanensis: A review focusing in endemic areas. Acta Trop. 2010, 115, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Burstein Alva, S. Ciclosporosis: Una parasitosis emergente. (II) Diagnóstico microbiológico mediante una nueva técnica de coloración. Rev. Gastroenterol. Perú 2005, 25, 336–340. [Google Scholar] [PubMed]
- Fletcher, S.M.; Stark, D.; Ellis, J. Prevalence of gastrointestinal pathogens in Sub-Saharan Africa: Systematic review and meta-analysis. J. Public Health Afr. 2011, 2, e30. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Gasser, R.B. Human cyclosporiasis. Lancet Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, G.E.; Martínez-Loya, O.A.; Ortega, Y.R. Cyclospora cayetanensis in a pediatric hospital in Morelia, México. Am. J. Trop. Med. Hyg. 2014, 91, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, D.; Acosta, M.E.; Acosta, M.E.; Morales, P. Estudio clínico-epidemiológico de coccidiosis intestinales en una población rural de región semiárida del estado Falcón, Venezuela. Investig. Clin. 2012, 53, 273–288. [Google Scholar]
- Thima, K.; Mori, H.; Praevanit, R.; Mongkhonmu, S.; Waikagul, J.; Watthanakulpanich, D. Recovery of Cyclospora cayetanensis among asymptomatic rural Thai schoolchildren. Asian Pac. J. Trop. Med. 2014, 7, 119–123. [Google Scholar] [CrossRef]
- Dhanabal, J.; Selvadoss, P.P.; Muthuswamy, K. Comparative study of the prevalence of intestinal parasites in low socioeconomic areas from South Chennai, India. J. Parasitol. Res. 2014, 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Tandukar, S.; Ansari, S.; Adhikari, N.; Shrestha, A.; Gautam, J.; Sharma, B.; Rajbhandari, D.; Gautam, S.; Nepal, H.P.; Sherchand, J.B. Intestinal parasitosis in school children of Lalitpur district of Nepal. BMC Res. Notes 2013, 6, 449. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, D.; Tandukar, S.; Parajuli, H.; Thapa, P.; Chaudhary, P.; Shrestha, D.; Shah, P.K.; Sherchan, J.B.; Sherchand, J.B. Cyclospora infection among school children in Kathmandu, Nepal: Prevalence and associated risk factors. Trop. Med. Health 2015, 43, 211–216. [Google Scholar] [CrossRef] [PubMed]
- De la Luz Galván-Ramírez, M.; Madriz-Elisondo, A.L.; Ramírez, C.G.T.; de Jesús Romero Rameño, J.; de la O Carrasco, D.A.; Lopez, M.A.C. Enteroparasitism and risk factors associated with clinical manifestations in children and adults of Jalisco State in Western Mexico. Osong Public Health Res. Perspect 2019, 10, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Nace, E.K.; Freeman, A.R.; Streit, T.G.; da Silva, A.J.; Lammie, P.J. Cyclospora cayetanensis infections in Haiti: A common occurrence in the absence of watery diarrhea. Am. J. Trop. Med. Hyg. 1999, 60, 584–586. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Hernandez, B.; Lopez, M.B.; Arrowood, M.J.; de Mejia, M.A.; de Merida, A.M.; Hightower, A.W.; Venczel, L.; Herwaldt, B.L.; Klein, R.E. Epidemiologic studies of Cyclospora cayetanensis in Guatemala. Emerg. Infect. Dis. 1999, 5, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Arrowood, M.J.; Eberhard, M.; Maguire, J.H. Cyclospora in Guatemala: Further considerations. J. Clin. Microbiol. 2002, 40, 731–732. [Google Scholar] [CrossRef]
- Hall, R.L.; Jones, J.L.; Hurd, S.; Smith, G.; Mahon, B.E.; Herwaldt, B.L. Population-based active surveillance for Cyclospora infection-United States, Foodborne Diseases Active Surveillance Network (FoodNet), 1997–2009. Clin. Infect. Dis. 2012, 54 (Suppl. 5), S411–S417. [Google Scholar] [CrossRef]
- Kaminsky, R.G.; Lagos, J.; Raudales Santos, G.; Urrutia, S. Marked seasonality of Cyclospora cayetanensis infections: Ten-year observation of hospital cases, Honduras. BMC Infect. Dis. 2016, 16, 66. [Google Scholar] [CrossRef]
- Jiang, Y.; Yuan, Z.; Zang, G.; Li, D.; Wang, Y.; Zhang, Y.; Liu, H.; Cao, J.; Shen, Y. Cyclospora cayetanensis infections among diarrheal outpatients in Shanghai: A retrospective case study. Front. Med. 2018, 12, 98–103. [Google Scholar] [CrossRef]
- Ortega, Y.R.; Robertson, L.J. Cyclospora Cayetanensis as A Foodborne Pathogen (Springer Briefs in Food, Health, and Nutrition), 1st ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–65. [Google Scholar]
- Madico, G.; Gilman, R.H.; Miranda, E.; Cabrera, L.; Sterling, C.R. Treatment of Cyclospora infections with co-trimoxazole. Lancet 1993, 342, 122–123. [Google Scholar] [CrossRef]
- Madico, G.; McDonald, J.; Gilman, R.H.; Cabrera, L.; Sterling, C.R. Epidemiology and treatment of Cyclospora cayetanensis infection in Peruvian children. Clin. Infect. Dis. 1997, 24, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Nimri, L.F. Cyclospora cayetanensis and other intestinal parasites associated with diarrhea in a rural area of Jordan. Int. Microbiol. 2003, 6, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Turgay, N.; Yolasigmaz, A.; Erdogan, D.D.; Zeyrek, F.Y.; Uner, A. Incidence of cyclosporosis in patients with gastrointestinal symptoms in western Turkey. Med. Sci. Monit. 2007, 13, CR34–CR39. [Google Scholar] [PubMed]
- Tuli, L.; Gulati, A.K.; Sundar, S.; Mohapatra, T.M. Correlation between CD4 counts of HIV patients and enteric protozoan in different seasons—An experience of a tertiary care hospital in Varanasi (India). BMC Gastroenterol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Tram, N.T.; Hoang, L.M.N.; Cam, P.D.; Chung, P.T.; Fyfe, M.W.; Isaac-Renton, J.L.; Ong, C.S.L. Cyclospora spp. in herbs and water samples collected from markets and farms in Hanoi, Vietnam. Trop. Med. Int. Health 2008, 13, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.L.; Jones, J.L.; Herwaldt, B.L. Surveillance for laboratory-confirmed sporadic cases of cyclosporiasis—United States, 1997–2008. MMWR Surveill. Summ. 2011, 60, 1–11. [Google Scholar] [PubMed]
- Sherchand, J.B.; Cross, J.H.; Jimba, M.; Sherchand, S.; Shrestha, M.P. Study of Cyclospora cayetanensis in health care facilities, sewage water and green leafy vegetables in Nepal. Southeast Asian J. Trop. Med. Public Health 1999, 30, 58–63. [Google Scholar]
- El-Karamany, E.M.N.; Zaher, T.I.; El-Bahnasawy, M.M. Role of water in the transmission of cyclosporiasis in Sharkia Governorate, Egypt. J. Egypt. Soc. Parasitol. 2005, 35, 953–962. [Google Scholar]
- Lopez, A.S.; Bendik, J.M.; Alliance, J.Y.; Roberts, J.M.; da Silva, A.J.; Moura, I.N.S.; Arrowood, M.J.; Eberhard, M.L.; Herwaldt, B.L. Epidemiology of Cyclospora cayetanensis and other intestinal parasites in a community in Haiti. J. Clin. Microbiol. 2003, 41, 2047–2054. [Google Scholar] [CrossRef]
- Gajadhar, A.A.; Lalonde, L.F.; Al-Adhami, B.; Singh, B.B.; Lobanov, V. Foodborne apicomplexan protozoa: Coccidia. In Foodborne Parasites in the Food Supply Web: Occurrence and Control; Gajadhar, A.A., Ed.; Woodhead Publishing: Sawston, UK, 2015. [Google Scholar]
- Hoge, C.W.; Shlim, D.R.; Ghimire, M.; Rabold, J.G.; Pandey, P.; Walch, A.; Rajah, R.; Gaudio, P.; Echevarria, P. Placebo-controlled trial of co-trimoxazole for Cyclospora infections among travellers and foreign residents in Nepal. Lancet 1995, 345, 691–693. [Google Scholar] [CrossRef]
- Sherchan, J.B.; Sherpa, K.; Tandukar, S.; Cross, J.H.; Gajadhar, A.; Sherchand, J.B. Infection of Cyclospora cayetanensis in diarrhoeal children of Nepal. J. Nepal Paed. Soc. 2010, 30, 23–30. [Google Scholar] [CrossRef]
- El Fatni, C.; Olmo, F.; El Fatni, H.; Romero, D.; Rosales, M.J. First genotyping of Giardia duodenalis and prevalence of enteroparasites in children from Tetouan (Morocco). Parasite 2014, 21, 48. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hafeez, E.H.; Ahmad, A.K.; Ali, B.A.; Moslam, F.A. Opportunistic parasites among immunosuppressed children in Minia District, Egypt. Korean J. Parasitol. 2012, 50, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Mbae, C.K.; Nokes, D.J.; Mulinge, E.; Nyambura, J.; Waruru, A.; Kariuki, S. Intestinal parasitic infections in children presenting with diarrhoea in outpatient and inpatient settings in an informal settlement of Nairobi, Kenya. BMC Infect. Dis. 2013, 13, 243. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Vats, O.; Kumar, D.; Singh, S. Coccidian intestinal parasites among immunocompetent children presenting with diarrhea: Are we missing them? Trop. Parasitol. 2017, 7, 37–40. [Google Scholar] [PubMed]
- Birdal Akis, F.; Beyhan, Y.E. Distribution of intestinal parasites in patients hospitalized in child intensive care unit. Turk. Parazitol. Derg. 2018, 42, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Frickmann, H.; Schwarz, N.G.; Rakotozandrindrainy, R.; May, J.; Hagen, R.M. PCR for enteric pathogens in high-prevalence settings. What does a positive signal tell us? Infect. Dis. 2015, 47, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Rizk, H.; Soliman, M. Coccidiosis among malnourished children in Mansoura, Dakahlia Governorate, Egypt. J. Egypt. Soc. Parasitol. 2001, 31, 877–886. [Google Scholar] [PubMed]
- Pratdesaba, R.A.; González, M.; Piedrasanta, E.; Mérida, C.; Contreras, K.; Vela, C.; Culajay, F.; Flores, L.; Torres, O. Cyclospora cayetanensis in three populations at risk in Guatemala. J. Clin. Microbiol. 2001, 39, 2951–2953. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hoge, C.W.; Echeverria, P.; Rajah, R.; Jacobs, J.; Malthouse, S.; Chapman, E.; Jimenez, L.M.; Shlim, D.R. Prevalence of Cyclospora species and other enteric pathogens among children less than 5 years of age in Nepal. J. Clin. Microbiol. 1995, 33, 3058–3060. [Google Scholar] [PubMed]
- Chacín-Bonilla, L.; Estévez, J.; Monsalve, F.; Quijada, L. Cyclospora cayetanensis infections among diarrheal patients from Venezuela. Am. J. Trop. Med. Hyg. 2001, 65, 351–354. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chacin-Bonilla, L.; Mejia de Young, M.; Estevez, J. Prevalence and pathogenic role of Cyclospora cayetanensis in a Venezuelan community. Am. J. Trop. Med. Hyg. 2003, 68, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Chacín-Bonilla, L.; Barrios, F.; Sanchez, Y. Epidemiology of Cyclospora cayetanensis infection in San Carlos Island, Venezuela: Strong association between socio-economic status and infection. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Mundaca, C.C.; Torres-Slimming, P.A.; Araujo-Castillo, R.V.; Morán, M.; Bacon, D.J.; Ortega, Y.; Gilman, R.H.; Blazes, D.L. Use of PCR to improve diagnostic yield in an outbreak of cyclosporiasis in Lima, Peru. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Torres-Slimming, P.A.; Mundaca, C.C.; Moran, M.; Quispe, J.; Colina, O.; Bacon, D.J.; Lescano, A.G.; Gilman, R.H.; Blazes, D.L. Outbreak of cyclosporiasis at a naval base in Lima, Peru. Am. J. Trop. Med. Hyg. 2006, 75, 546–548. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lv, B.; Wang, Q.; Wang, R.; Jian, F.; Zhang, L.; Ning, C.; Fu, K.; Wang, Y.; Qi, M.; et al. Prevalence and molecular characterization of Cyclospora cayetanensis, Henan, China. Emerg. Infect. Dis. 2011, 17, 1887–1890. [Google Scholar] [CrossRef] [PubMed]
- Bhattachan, B.; Sherchand, J.B.; Tandukar, S.; Dhoubhadel, B.G.; Gauchan, L.; Rai, G. Detection of Cryptosporidium parvum and Cyclospora cayetanensis infections among people living in a slum area in Kathmandu valley, Nepal. BMC Res. Notes 2017, 10, 464. [Google Scholar] [CrossRef]
- Wang, K.X.; Li, C.P.; Wang, J.; Tian, Y. Cyclospore cayetanensis in Anhui, China. World J. Gastroenterol. 2002, 8, 1144–1148. [Google Scholar] [CrossRef]
- Döller, P.C.; Dietrich, K.; Filipp, N.; Brockmann, S.; Dreweck, C.; Vonthein, R.; Wagner-Wiening, C.; Wiedenmann, A. Cyclosporiasis outbreak in Germany associated with the consumption of salad. Emerg. Infect. Dis. 2002, 8, 992–994. [Google Scholar] [CrossRef]
- Gómez Martínez, E.; Figuera, L.; Guilarte, D.V.; Simoni, Z.; Díaz, M.T.; Berrizbeitia, M.; Cerrada, S. Primer reporte de Cyclospora cayetanensis en una comunidad indígena Kariña del municipio Sucre, estado Sucre, Venezuela. Boletín de Malariología y Salud Ambiental 2016, 56, 24–38. [Google Scholar]
- Soave, R. Cyclospora: An overview. Clin. Infect. Dis. 1996, 23, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Gumbo, T.; Sarbah, S.; Gangaidzo, I.T.; Ortega, Y.; Sterling, C.R.; Carville, A.; Tzipori, S.; Wiest, P.W. Intestinal parasites in patients with diarrhea and human immunodeficiency virus infection in Zimbabwe. AIDS 1999, 13, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Verdier, R.I.; Fitzgerald, D.W.; Johnson, W.D., Jr.; Pape, J.W. Trimethoprim-sulfamethoxazole compared with ciprofloxacin for treatment and prophylaxis of Isospora belli and Cyclospora cayetanensis infection in HIV-infected patients: A randomized, controlled trial. Ann. Intern. Med. 2000, 132, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L. Cyclospora cayetanensis: A review, focusing on the outbreaks of cyclosporiasis in the 1990s. Clin. Infect. Dis. 2000, 31, 1040–1057. [Google Scholar] [CrossRef] [PubMed]
- Asma, I.; Johari, S.; Sim, B.L.H.; Lim, Y.A.L. How common is intestinal parasitism in HIV-infected patients in Malaysia? Trop. Biomed. 2011, 28, 400–410. [Google Scholar] [PubMed]
- Sarfati, C.; Bourgeois, A.; Menotti, J.; Liegeois, F.; Moyou-Somo, R.; Delaporte, E.; Derouin, F.; Ngole, E.M.; Molina, J.M. Prevalence of intestinal parasites including microsporidia in human immunodeficiency virus-infected adults in Cameroon: A cross-sectional study. Am. J. Trop. Med. Hyg. 2006, 74, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Raccurt, C.P.; Fouché, B.; Agnamey, P.; Menotti, J.; Chouaki, T.; Totet, A.; Pape, J.W. Short report: Presence of Enterocytozoon bieneusi associated with intestinal coccidia in patients with chronic diarrhea visiting an HIV Center in Haiti. Am. J. Trop. Med. Hyg. 2008, 79, 579–580. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Rodríguez, Z.; Hernández, A.; Bracho, Á.; Salazar, S.; Villalobos, R. Prevalencia de microsporidiosis intestinales y otros entero enteroparásitos en pacientes con VIH positivo de Maracaibo, Venezuela. Biomédica 2013, 33, 538–545. [Google Scholar] [CrossRef][Green Version]
- Dash, M.; Padhi, S.; Panda, P.; Parida, B. Intestinal protozoans in adults with diarrhea. N. Am. J. Med. Sci. 2013, 5, 707–712. [Google Scholar] [CrossRef]
- Mathur, M.K.; Verma, A.K.; Makwana, G.E.; Sinha, M. Study of opportunistic intestinal parasitic infections in human immunodeficiency virus/acquired immunodeficiency syndrome patients. J. Glob. Infect. Dis. 2013, 5, 164–167. [Google Scholar] [CrossRef]
- Nsagha, D.S.; Njunda, A.L.; Assob, N.J.C.; Ayima, C.W.; Tanue, E.A.; Kibu, O.D.; Kwenti, T.E. Intestinal parasitic infections in relation to CD4+ T cell counts and diarrhea in HIV/AIDS patients with or without antiretroviral therapy in Cameroon. BMC Infect. Dis. 2016, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Helmy, M.M.F.; Rashed, L.A.; Abdel-Fattah, H.S. Co-infection with Cryptosporidium parvum and Cyclospora cayetanensis in immunocompromised patients. J. Egypt. Soc. Parasitol. 2006, 36, 613–627. [Google Scholar] [PubMed]
- Uysal, H.K.; Adas, G.T.; Atalik, K.; Altiparmak, S.; Akgul, O.; Saribas, S.; Gurcan, M.; Yuksel, P.; Yildirmak, T.; Kocazeybek, B.; et al. The Prevalence of Cyclospora cayetanensis and Cryptosporidium spp. in Turkish patients infected with HIV-1. Acta Parasitol. 2017, 62, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, A.; Karyadi, T.; Dwintasari, S.W.; Sari, I.P.; Yunihastuti, E.; Djauzi, S.; Smith, H.V. Intestinal parasitic infections in HIV/AIDS patients presenting with diarrhoea in Jakarta, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Sanad, M.M.; Thagfan, F.A.; Al Olayan, E.M.; Almogren, A.; Al Hammaad, A.; Al-Mawash, A.; Mohamed, A.A. Opportunistic coccidian parasites among Saudi cancer patients presenting with diarrhea: Prevalence and immune status. Res. J. Parasitol. 2014, 9, 55–63. [Google Scholar] [CrossRef]
- Thomas, M.K.; Murray, R.; Flockhart, L.; Pintar, K.; Pollari, F.; Fazil, A.; Nesbitt, A.; Marshall, B. Estimates of the burden of foodborne illness in Canada for 30 specified pathogens and unspecified agents, circa 2006. Foodborne Pathog. Dis. 2013, 10, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Gascón, J.; Álvarez, M.; Valls, M.E.; Bordas, J.M.; de Anta, M.T.J.; Corachán, M. Ciclosporiasis: Estudio clinicoepidemiológico en viajeros con Cyclospora cayetanensis importada. Med. Clin. 2001, 116, 461–464. [Google Scholar] [CrossRef]
- Puente, S.; Morente, A.; García-Benayas, T.; Subirats, M.; Gascón, J.; González-Lahoz, J.M. Cyclosporiasis: A point source outbreak acquired in Guatemala. J. Travel Med. 2006, 13, 334–337. [Google Scholar] [CrossRef]
- Ramírez-Olivencia, G.; Herrero, M.D.; Subirats, M.; Rivas González, P.; Puente, S. Brote de Cyclospora cayetanensis en viajeros a Cuba. Enferm. Infecc. Microbiol. Clin. 2008, 26, 558–560. [Google Scholar] [CrossRef]
- Nichols, G.L.; Freedman, J.; Pollock, K.G.; Rumble, C.; Chalmers, R.M.; Chiodini, P.; Hawkins, G.; Alexander, C.L.; Godbole, G.; Williams, C.; et al. Cyclospora infection linked to travel to Mexico, June to September 2015. Euro Surveill. 2015, 20, 30048. [Google Scholar] [CrossRef]
- Marques, D.F.P.; Alexander, C.L.; Chalmers, R.M.; Elson, R.; Freedman, J.; Hawkins, G.; Lo, J.; Robinson, G.; Russell, K.; Smith-Palmer, A.; et al. Cyclosporiasis in travellers returning to the United Kingdom from Mexico in summer 2017: Lessons from the recent past to inform the future. Euro Surveill. 2017, 22, 30592. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thapa, S.S.; Basnyat, B. Chronic diarrhea in a traveler: Cyclosporiasis. Am. J. Med. 2017, 130, e536. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, M.; Bajer, A.; Welc-Faleciak, R.; Pawelas, A. Cyclospora cayetanensis infection in transplant traveller: A case report of outbreak. Parasit. Vectors 2015, 8, 411. [Google Scholar] [CrossRef] [PubMed]
- Paschke, C.; Apelt, N.; Fleischmann, E.; Perona, P.; Walentiny, C.; Loscher, T.; Herbinger, K.H. Controlled study on enteropathogens in travellers returning from the tropics with and without diarrhoea. Clin. Microbiol. Infect. 2011, 17, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Jelinkova, A.; Kasickova, D.; Valkoun, A. Cyclospora cayetanensis—The rare causal agent of diarrhoeal diseases. Klin. Mikrobiol. Infekc. Lek. 2011, 17, 86–88. [Google Scholar] [PubMed]
- Blans, M.C.A.; Ridwan, B.U.; Verweij, J.J.; Rozenberg-Arska, M.; Verhoef, J. Cyclosporiasis outbreak, Indonesia. Emerg. Infect. Dis. 2005, 11, 1453–1455. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef]
- Scallan, E.; Griffin, P.M.; Angulo, F.J.; Tauxe, R.V.; Hoekstra, R.M. Foodborne illness acquired in the United States-unspecified agents. Emerg. Infect. Dis. 2011, 17, 16–22. [Google Scholar] [CrossRef]
- Butler, A.J.; Thomas, M.K.; Pintar, K.D.M. Expert elicitation as a means to attribute 28 enteric pathogens to foodborne, waterborne, animal contact, and person-to-person transmission routes in Canada. Foodborne Pathog. Dis. 2015, 12, 335–344. [Google Scholar] [CrossRef]
- Dawson, D. Foodborne protozoan parasites. Int. J. Food Microbiol. 2005, 103, 207–227. [Google Scholar] [CrossRef]
- Anh, V.T.; Tram, N.T.; Klank, L.T.; Cam, P.D.; Dalsgaard, A. Faecal and protozoan parasite contamination of water spinach (Ipomoea aquatica) cultivated in urban wastewater in Phnom Penh, Cambodia. Trop. Med. Int. Health 2007, 12, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.; Carazo, M.; Arias, M.L.; Chaves, C.; Monge, R.; Chinchilla, M. Prevalencia de Cyclospora sp., Cryptosporidium sp., microsporidos y determinación de coliformes fecales en frutas y vegetales frescos de consumo crudo en Costa Rica. Arch. Latinoam. Nutr. 2004, 54, 428–432. [Google Scholar] [PubMed]
- Dixon, B.; Parrington, L.; Cook, A.; Pollari, F.; Farber, J. Detection of Cyclospora, Cryptosporidium, and Giardia in ready-to-eat packaged leafy greens in Ontario, Canada. J. Food Prot. 2013, 76, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.F.; Gajadhar, A.A. Detection of Cyclospora cayetanensis, Cryptosporidium spp., and Toxoplasma gondii on imported leafy green vegetables in Canadian survey. Food Waterborne Parasitol. 2016, 2, 8–14. [Google Scholar] [CrossRef]
- Abou el Naga, I.F. Studies on a newly emerging protozoal pathogen: Cyclospora cayetanensis. J. Egypt. Soc. Parasitol. 1999, 29, 575–586. [Google Scholar]
- El Said Said, D. Detection of parasites in commonly consumed raw vegetables. Alex. J. Med. 2012, 48, 345–352. [Google Scholar] [CrossRef]
- Mossallam, S.F. Detection of some intestinal protozoa in commercial fresh juices. J. Egypt. Soc. Parasitol. 2010, 40, 135–149. [Google Scholar]
- Bekele, F.; Tefera, T.; Biresaw, G.; Yohannes, T. Parasitic contamination of raw vegetables and fruits collected from selected local markets in Arba Minch town, Southern Ethiopia. Infect. Dis. Poverty 2017, 6, 19. [Google Scholar] [CrossRef]
- Duedu, K.O.; Yarnie, E.A.; Tetteh-Quarcoo, P.B.; Attah, S.K.; Donkor, E.S.; Ayeh-Kumi, P.F. A comparative survey of the prevalence of human parasites found in fresh vegetables sold in supermarkets and open-aired markets in Accra, Ghana. BMC Res. Notes 2014, 7, 836. [Google Scholar] [CrossRef]
- Giangaspero, A.; Marangi, M.; Koehler, A.V.; Papini, R.; Normanno, G.; Lacasella, V.; Lonigro, A.; Gasser, R.B. Molecular detection of Cyclospora in water, soil, vegetables and humans in southern Italy signals a need for improved monitoring by health authorities. Int. J. Food Microbiol. 2015, 211, 95–100. [Google Scholar] [CrossRef]
- Caradonna, T.; Marangi, M.; Del Chierico, F.; Ferrari, N.; Reddel, S.; Bracaglia, G.; Normanno, G.; Putignani, L.; Giangaspero, A. Detection and prevalence of protozoan parasites in ready-to-eat packaged salads on sale in Italy. Food Microbiol. 2017, 67, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Won, J.; Kim, J.W.; Kim, K.; Park, W.Y.; Yu, J.R. Simultaneous molecular detection of Cryptosporidium and Cyclospora from raw vegetables in Korea. Korean J. Parasitol. 2017, 55, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Roxas, C.R.; Gilman, R.H.; Miller, N.J.; Cabrera, L.; Taquiri, C.; Sterling, C.R. Isolation of Cryptosporidium parvum and Cyclospora cayetanensis from vegetables collected in markets of an endemic region in Peru. Am. J. Trop. Med. Hyg. 1997, 57, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Devera, R.; Blanco, Y.; González, H.; García, L. Parásitos intestinales en lechugas comercializadas en mercados populares y supermercados de Ciudad Bolívar, Estado Bolívar, Venezuela. Revista de la Sociedad Venezolana de Microbiología 2006, 26, 100–107. [Google Scholar]
- Almeria, S.; da Silva, A.J.; Blessington, T.; Cloyd, T.C.; Cinar, H.N.; Durigan, M.; Murphy, H.R. Evaluation of the U.S. Food and Drug Administration validated method for detection of Cyclospora cayetanensis in high-risk fresh produce matrices and a method modification for a prepared dish. Food Microbiol. 2018, 76, 497–503. [Google Scholar] [CrossRef]
- Ho, A.Y.; Lopez, A.S.; Eberhart, M.G.; Levenson, R.; Finkel, B.S.; da Silva, A.J.; Roberts, J.M.; Orlandi, P.A.; Johnson, C.C.; Herwaldt, B.L. Outbreak of cyclosporiasis associated with imported raspberries, Philadelphia, Pennsylvania, 2000. Emerg. Infect. Dis. 2002, 8, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Hoang, L.M.N.; Fyfe, M.; Ong, C.; Harb, J.; Champagne, S.; Dixon, B.; Isaac-Renton, J. Outbreak of cyclosporiasis in British Columbia associated with imported Thai basil. Epidemiol. Infect. 2005, 133, 23–27. [Google Scholar] [CrossRef]
- Insulander, M.; Svenungsson, B.; Lebbad, M.; Karlsson, L.; de Jong, B. A foodborne outbreak of Cyclospora infection in Stockholm, Sweden. Foodborne Pathog. Dis. 2010, 7, 1585–1587. [Google Scholar] [CrossRef]
- Gibbs, R.A.; Nanyonjo, R.; Pingault, N.M.; Combs, B.G.; Mazzucchelli, T.; Armstrong, P.; Tarling, G.; Dowse, G.K. An outbreak of Cyclospora infection on a cruise ship. Epidemiol. Infect. 2013, 141, 508–516. [Google Scholar] [CrossRef]
- Dixon, B.R. Parasitic illnesses associated with the consumption of fresh produce—An emerging issue in developed countries. Curr. Opin. Food Sci. 2016, 8, 104–109. [Google Scholar] [CrossRef]
- Buss, B.F.; Joshi, M.V.; O’Keefe, A.L.; Allensworth, C.D.; Garvey, A.; Obbink, K.; Mandernach, S.; Safranek, T.J. Regional investigation of a cyclosporiasis outbreak linked to imported romaine lettuce—Nebraska and Iowa, June-August 2013. Epidemiol. Infect. 2016, 144, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Buss, B.F.; Joshi, M.V.; Dement, J.L.; Cantu, V.; Safranek, T.J. Multistate porduct traceforward investigation to link imported romaine lettuce to a US cyclosporoiasis outbreak—Nebraska, Texas, and Florida, June–August 2013. Epidemiol. Infect. 2016, 144, 2709–2718. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Whitfield, Y.; Johnson, K.; Hanson, H.; Huneault, D. 2015 outbreak of cyclosporiasis linked to the consumption of imported sugar snap peas in Ontario, Canada. J. Food Prot. 2017, 80, 1666–1669. [Google Scholar] [CrossRef] [PubMed]
- Zini, R.M.; Santos, C.C.M.; Almeida, I.A.Z.C.; Peresi, J.T.M.; Marques, C.C.A. Public health laboratory action for elucidation of diarrhea outbreak by Cyclospora cayetanensis in General Salgado Municipality, SP. Rev. Inst. Adolfo Lutz 2004, 63, 116–121. [Google Scholar]
- Dixon, B.R.; Fayer, R.; Santín, M.; Hill, D.E.; Dubey, J.P. Protozoan parasites: Cryptosporidium, Giardia, Cyclospora, and Toxoplasma. In Rapid Detection, Characterization, and Enumeration of Food-Borne Pathogens; Hoorfar, J., Ed.; ASM Press: Washington, DC, USA, 2011. [Google Scholar]
- Herwaldt, B.L.; Ackers, M.L.; The Cyclospora Working Group. An outbreak in 1996 of cyclosporiasis associated with imported raspberries. N. Engl. J. Med. 1997, 336, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Manuel, D.G.; Neamatullah, S.; Shahin, R.; Reymond, D.; Keystone, J.; Carlson, J.; Le Ber, C.; Herwaldt, B.L.; Werker, D.H. An outbreak of cyclosporiasis in 1996 associated with consumption of fresh berries—Ontario. Can. J. Infect. Dis. 2000, 11, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Beach, M.J.; The Cyclospora Working Group. The return of Cyclospora in 1997: Another outbreak of cyclosporiasis in North America associated with imported raspberries. Ann. Intern. Med. 1999, 130, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Kozak, G.K.; MacDonald, D.; Landry, L.; Farber, J.M. Foodborne outbreaks in Canada linked to produce: 2001 through 2009. J. Food Prot. 2013, 76, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Milord, F.; Lampron-Goulet, E.; St-Amour, M.; Levac, E.; Ramsay, D. Cyclospora cayetanensis: A description of clinical aspects of an outbreak in Quebec, Canada. Epidemiol. Infect. 2012, 140, 626–632. [Google Scholar] [CrossRef]
- Shah, L.; MacDougall, L.; Ellis, A.; Ong, C.; Shyng, S.; LeBlanc, L. The British Columbia Cyclospora Investingation Team Challenges of investigating community outbreaks of cyclosporiasis, British Columbia, Canada. Emerg. Infect. Dis. 2009, 15, 1286–1288. [Google Scholar] [CrossRef]
- Dixon, B.; Mihajlovic, B.; Couture, H.; Farber, J.M. Qualitative risk assessment: Cyclospora cayetanensis on fresh raspberries and blackberries imported into Canada. Food Prot. Trends 2016, 36, 18–32. [Google Scholar]
- CFIA-PHAC Public Health Notice—Outbreak of Cyclospora Infections Under Investigation October 11 2017—Final Update. Available online: https://www.canada.ca/en/public-health/services/public-health-notices/2017/public-health-notice-outbreak-cyclospora-infections-under-investigation.html (accessed on 28 June 2019).
- Botero-Garces, J.; Montoya-Palacio, M.N.; Barguil, J.I.; Castaño-González, A. Brote epidémico por Cyclospora cayetanensis en Medellín, Colombia. Rev. Salud Pública 2006, 8, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Shlim, D.R.; Cohen, M.T.; Eaton, M.; Rajah, R.; Long, E.G.; Ungar, B.L.P. An alga-like organism associated with an outbreak of prolonged diarrhea among foreigners in Nepal. Am. J. Trop. Med. Hyg. 1991, 45, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Rabold, J.G.; Hoge, C.W.; Shlim, D.R.; Kefford, C.; Rajah, R.; Echeverria, P. Cyclospora outbreak associated with chlorinated drinking water. Lancet 1994, 344, 1360–1361. [Google Scholar] [CrossRef]
- Ayala-Gaytán, J.J.; Díaz-Olachea, C.; Riojas-Montalvo, P.; Palacios-Martínez, C. Ciclosporidiosis: Características clínicas y diagnósticas de un brote epidémico. Rev. Gastroenterol. Mex. 2004, 69, 226–229. [Google Scholar] [PubMed]
- Aksoy, U.; Akisu, C.; Sahin, S.; Usluca, S.; Yalcin, G.; Kuralay, F.; Oral, A.M. First reported waterborne outbreak of cryptosporidiosis with Cyclospora co-infection in Turkey. Eurosurveillance 2007, 12, E070215. [Google Scholar] [CrossRef]
- Ozdamar, M.; Hakko, E.; Turkoglu, S. High occurrence of cyclosporiasis in Istanbul, Turkey, during a dry and warm summer. Parasit. Vectors 2010, 3, 39. [Google Scholar] [CrossRef]
- Huang, P.; Weber, J.T.; Sosin, D.M.; Griffin, P.M.; Long, E.G.; Murphy, J.J.; Kocka, F.; Peters, C.; Kallick, C. The first reported outbreak of diarrheal illness associated with Cyclospora in the United States. Ann. Intern. Med. 1995, 123, 409–414. [Google Scholar] [CrossRef]
- Koumans, E.H.A.; Katz, D.J.; Malecki, J.M.; Kumar, S.; Wahlquist, S.P.; Arrowood, M.J.; Hightower, A.W.; Herwaldt, B.L. An outbreak of cyclosporiasis in Florida in 1995: A harbinger of multistate outbreaks in 1996 and 1997. Am. J. Trop. Med. Hyg. 1998, 59, 235–242. [Google Scholar] [CrossRef]
- Fleming, C.A.; Caron, D.; Gunn, J.E.; Barry, M.A. A foodborne outbreak of Cyclospora cayetanensis at a wedding: Clinical features and risk factors for illness. Arch. Intern. Med. 1998, 158, 1121–1125. [Google Scholar] [CrossRef]
- Katz, D.; Kumar, S.; Malecki, J.; Lowdermilk, M.; Koumans, E.H.A.; Hopkins, R. Cyclosporiasis associated with imported raspberries, Florida, 1996. Public Health Rep. 1999, 114, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, V.M.; Ball, R.T.; Somerfeldt, S.A.; Mackey, L.; Nichols, S.E.; MacKenzie, W.R.; Herwaldt, B.L. A foodborne outbreak of cyclosporiasis caused by imported raspberries. J. Fam. Pract. 1998, 47, 231–234. [Google Scholar] [PubMed]
- Lopez, A.S.; Dodson, D.R.; Arrowood, M.J.; Orlandi, P.A., Jr.; da Silva, A.J.; Bier, J.W.; Hanauer, S.D.; Kuster, R.L.; Oltman, S.; Baldwin, M.S.; et al. Outbreak of cyclosporiasis associated with basil in Missouri in 1999. Clin. Infect. Dis. 2001, 32, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. U.S. Foodborne Outbreaks of Cyclosporiasis—2000–2017. Cyclosporiasis. Available online: http://www.cdc.gov/parasites/cyclosporiasis/outbreaks/foodborneoutbreaks.html (accessed on 27 June 2019).
- CDC Centers for Disease Control and Prevention. Outbreak of cyclosporiasis associated with snow peas—Pennsylvania, 2004. MMWR 2004, 53, 876–878. Available online: https://cdc.gov/mmwr/preview/mmwrhtml/mm5337a6.htm (accessed on 20 June 2019).
- Abanyie, F.; Harvey, R.R.; Harris, J.R.; Wiegand, R.E.; Gaul, L.; Desvignes-Kendrick, M.; Irvin, K.; Williams, I.; Hall, R.L.; Herwaldt, B.; et al. 2013 multistate outbreaks of Cyclospora cayetanensis infections associated with fresh produce: Focus on the Texas investigations. Epidemiol. Infect. 2015, 143, 3451–3458. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.C. Restaurant-associated outbreak of cyclosporiasis in Austin, Texas 2016. In Proceedings of the 2017 CSTE Annual Conference, Austin, TX, USA, 4–8 June 2017. [Google Scholar]
- CDC Centers for Disease Control and Prevention. Domestically Acquired Cases of Cyclosproriasis—United States, May–August 2018. Available online: http://www.cdc.gov/parasites/cyclosporiasis/outbreaks/2018/c-082318/index.html (accessed on 27 June 2019).
- Casillas, S.M.; Bennett, C.; Straily, A. Multiple cyclosporiasis outbreaks—United States, 2018. MMWR 2018, 67, 1101–1111. [Google Scholar]
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Cyclosporiasis Linked to Fresh Expdress Salad Mix Sold at McDonald’s Restaurants—Nited States, 2018: Final Update. Available online: http://www.cdc.gov/parasites/cyclosporiasis/outbreaks/2018/b-071318/index.html (accessed on 24 June 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Cyclosporiasis Linked to Del Monte Fresh Produce Vegetable Trays—United States, 2018: Final Update. Available online: http://cdc.gov/parasites/cyclosporiasis/outbreaks/2018/a-062018/index.html (accessed on 24 June 2019).
- Murphy, H.R.; Lee, S.; da Silva, A.J. Evaluation of an improved U.S. Food and Drug Administration method for the detection of Cyclospora cayetanensis in produce using real-time PCR. J. Food Prot. 2017, 80, 1133–1144. [Google Scholar] [CrossRef]
- Tefera, T.; Tysnes, K.R.; Utaaker, K.S.; Robertson, L.J. Parasite contamination of berries: Risk, occurrence, and approaches for mitigation. Food Waterborne Parasitol. 2018, 10, 23–38. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention. Outbreaks of cyclosporiasis—United States, June–August 2013. MMWR 2013, 62, 862. [Google Scholar]
- Sturbaum, G.D.; Ortega, Y.R.; Gilman, R.H.; Sterling, C.R.; Cabrera, L.; Klein, D.A. Detection of Cyclospora cayetanensis in wastewater. Appl. Environ. Microbiol. 1998, 64, 2284–2286. [Google Scholar]
- Khalifa, A.M.; El Temsahy, M.M.; bou El Naga, I.F. Effect of ozone on the viability of some protozoa in drinking water. J. Egypt. Soc. Parasitol. 2001, 31, 603–616. [Google Scholar] [PubMed]
- Elshazly, A.M.; Elsheikha, H.M.; Soltan, D.M.; Mohammad, K.A.; Morsy, T.A. Protozoal pollution of surface water sources in Dakahlia Governorate, Egypt. J. Egypt. Soc. Parasitol. 2007, 37, 51–64. [Google Scholar] [PubMed]
- Khalifa, R.M.A.; Ahmad, A.K.; Abdel-Hafeez, E.H.; Mosllem, F.A. Present status of protozoan pathogens causing water-borne disease in northern part of El-Minia Governorate, Egypt. J. Egypt. Soc. Parasitol. 2014, 44, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Kwakye-Nuako, G.; Borketey, P.B.; Mensah-Attipoe, I.; Asmah, R.H.; Ayeh-Kumi, P.F. Sachet drinking water in Accra: The potential threats of transmission of enteric pathogenic protozoan organisms. Ghana Med. J. 2007, 41, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Dowd, S.E.; John, D.; Eliopolus, J.; Gerba, C.P.; Naranjo, J.; Klein, R.; López, B.; de Mejía, M.; Mendoza, C.E.; Pepper, I.L. Confirmed detection of Cyclospora cayetanesis, Encephalitozoon intestinalis and Cryptosporidium parvum in water used for drinking. J. Water Health 2003, 1, 117–123. [Google Scholar] [CrossRef]
- Giangaspero, A.; Marangi, M.; Arace, E. Cyclospora cayetanensis travels in tap water on Italian trains. J. Water Health 2015, 13, 210–216. [Google Scholar] [CrossRef]
- Bilung, L.M.; Tahar, A.S.; Yunos, N.E.; Apun, K.; Lim, Y.A.L.; Nillian, E.; Hashim, H.F. Detection of Cryptosporidium and Cyclospora oocysts from environmental water for drinking and recreational activities in Sarawak, Malaysia. BioMed Res. Int. 2017, 2017, 4636420. [Google Scholar] [CrossRef]
- Galván, A.L.; Magnet, A.; Izquierdo, F.; Fenoy, S.; Rueda, C.; Vadillo, C.F.; Henriques-Gil, N.; del Aguila, C. Molecular characterization of human-pathogenic microsporidia and Cyclospora cayetanensis isolated from various water sources in Spain: A year-long longitudinal study. Appl. Environ. Microbiol. 2013, 79, 449–459. [Google Scholar] [CrossRef]
- Ben Ayed, L.; Yang, W.; Widmer, G.; Cama, V.; Ortega, Y.; Xiao, L. Survey and genetic characterization of wastewater in Tunisia for Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi, Cyclospora cayetanensis and Eimeria spp. J. Water Health 2012, 10, 431–444. [Google Scholar] [CrossRef]
- Karaman, U.; Koloren, Z.; Seferoglu, O.; Ayaz, E.; Demirel, E. Presence of parasites in environmental waters in Samsun and its districts (in Turkish). Turk. Parazitol. Derg. 2017, 41, 19–21. [Google Scholar] [CrossRef]
- Kitajima, M.; Haramoto, E.; Iker, B.C.; Gerba, C.P. Occurrence of Cryptosporidium, Giardia, and Cyclospora in influent and effluent water at wastewater treatment plants in Arizona. Sci. Total Environ. 2014, 484, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Miegeville, M.; Koubi, V.; Dan, L.C.; Barbier, J.P.; Cam, P.D. Cyclospora cayetanensis et sa présence en milieu hydrique à Hanoï (Vietnam). Étude dans l’environnement (eaux de forage, lacs et rivières). Bull. Soc. Pathol. Exot. 2003, 96, 149–152. [Google Scholar] [PubMed]
- Marshall, M.M.; Naumovitz, D.; Ortega, Y.; Sterling, C.R. Waterborne protozoan pathogens. Clin. Microbiol. Rev. 1997, 10, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Baldursson, S.; Karanis, P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks-an update 2004–2010. Water Res. 2011, 45, 6603–6614. [Google Scholar] [CrossRef]
- Mansfield, L.S.; Gajadhar, A.A. Cyclospora cayetanensis, a food- and waterborne coccidian parasite. Vet. Parasitol. 2004, 126, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Soave, R.; Herwaldt, B.L.; Relman, D.A. Cyclospora. Infect. Dis. Clin. 1998, 12, 1–12. [Google Scholar] [CrossRef]
- Mota, P.; Rauch, C.A.; Edberg, S.C. Microsporidia and Cyclospora: Epidemiology and assessment of risk from the environment. Crit. Rev. Microbiol. 2000, 26, 69–90. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Ortega, Y.R.; Conn, D.B. Recovery of waterborne oocysts of Cyclospora cayetanensis by Asian freshwater clams (Corbicula fluminea). Am. J. Trop. Med. Hyg. 1998, 59, 928–932. [Google Scholar] [CrossRef][Green Version]
- Negm, A.Y. Human pathogenic protozoa in bivalves collected from local markets in Alexandria. J. Egypt. Soc. Parasitol. 2003, 33, 991–998. [Google Scholar]
- Aksoy, U.; Marangi, M.; Papini, R.; Ozkoc, S.; Bayram Delibas, S.; Giangaspero, A. Detection of Toxoplasma gondii and Cyclospora cayetanensis in Mytilus galloprovincialis from Izmir Province coast (Turkey) by Real Time PCR/High-Resolution Melting analysis (HRM). Food Microbiol. 2014, 44, 128–135. [Google Scholar] [CrossRef]
- Ghozzi, K.; Marangi, M.; Papini, R.; Lahmar, I.; Challouf, R.; Houas, N.; Ben Dhiab, R.; Normanno, G.; Babba, H.; Giangaspero, A. First report of Tunisian coastal water contamination by protozoan parasites using mollusk bibalves as biological indicators. Mar. Pollut. Bull. 2017, 117, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. sites, 2015–2018. MMWR 2019, 68, 369–373. [Google Scholar] [PubMed]
- Bendall, R.P.; Lucas, S.; Moody, A.; Tovey, G.; Chiodini, P.L. Diarrhoea associated with cyanobacterium-like bodies: A new coccidian enteritis of man. Lancet 1993, 341, 590–592. [Google Scholar] [CrossRef]
- Behera, B.; Mirdha, B.R.; Makharia, G.K.; Bhatnagar, S.; Dattagupta, S.; Samantaray, J.C. Parasites in patients with malabsorption syndrome: A clinical study in children and adults. Dig. Dis. Sci. 2008, 53, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Hussein, E.M.; El-Moamly, A.A.; Dawoud, H.A.; Fahmy, H.; El-Shal, H.E.; Sabek, N.A. Real-time PCR and flow cytometry in detection of Cyclospora oocysts in fecal samples of symptomatic and asymptomatic pediatrics patients. J. Egypt. Soc. Parasitol. 2007, 37, 151–170. [Google Scholar] [PubMed]
- Schubach, T.M.; Neves, E.S.; Leite, A.C.; Araújo, A.Q.C.; de Moura, H. Cyclospora cayetanensis in an asymptomatic patient infected with HIV and HTLV-1. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 175. [Google Scholar] [CrossRef]
- Sancak, B.; Akyon, Y.; Ergüven, S. Cyclospora infection in five immunocompetent patients in a Turkish university hospital. J. Med. Microbiol. 2006, 55, 459–462. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Richardson, R.F., Jr.; Remler, B.F.; Katirji, B.; Murad, M.H. Guillain-Barré syndrome after Cyclospora infection. Muscle Nerve 1998, 21, 669–671. [Google Scholar] [CrossRef]
- Connor, B.A.; Johnson, E.; Soave, R. Reiter syndrome following protracted symptoms of Cyclospora infection. Emerg. Infect. Dis. 2001, 7, 453–454. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.S.; Arrowood, M.; Kokoskin, E.; Paltridge, G.P.; Pillai, D.R.; Procop, G.W.; Ryan, N.; Shimizu, R.Y.; Visvesvara, G. Laboratory diagnosis of parasites from the gastrointestinal tract. Clin. Microbiol. Rev. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Negm, A.Y. Identification of Cyclospora cayetanensis in stool using different stains. J. Egypt. Soc. Parasitol. 1998, 28, 429–436. [Google Scholar] [PubMed]
- Harrington, B.J. Microscopy of 4 pathogenic enteric protozoan parasites: A review. Labmedicine 2008, 39, 231–238. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Moura, H.; Kovacs-Nace, E.; Wallace, S.; Eberhard, M.L. Uniform staining of Cyclospora oocysts in fecal smears by a modified safranin technique with microwave heating. J. Clin. Microbiol. 1997, 35, 730–733. [Google Scholar] [PubMed]
- Maratim, A.C.; Kamar, K.K.; Ngindu, A.; Akoru, C.N.; Diero, L.; Sidle, J. Safranin staining of Cyclospora cayetanensis oocysts not requiring microwave heating. Br. J. Biomed. Sci. 2002, 59, 114–115. [Google Scholar] [CrossRef] [PubMed]
- Galvan-Díaz, A.L.; Herrera-Jaramillo, V.; Santos-Rodriguez, Z.M.; Delgado-Naranjo, M. Coloraciones Ziehl-Neelsen y Safraninia modificadas para el diagnóstico de Cyclospora cayetanensis. Rev. Salud Pública 2008, 10, 488–493. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bialek, R.; Binder, N.; Dietz, K.; Knobloch, J.; Zelck, U.E. Comparison of autofluorescence and iodine staining for detection of Isospora belli in feces. Am. J. Trop. Med. Hyg. 2002, 67, 304–305. [Google Scholar] [CrossRef]
- Long, E.G.; White, E.H.; Carmichael, W.W.; Quinlisk, P.M.; Raja, R.; Swisher, B.L.; Daugharty, H.; Cohen, M.T. Morphologic and staining characteristics of a Cyanobacterium-like organism associated with diarrhea. J. Infect. Dis. 1991, 164, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Varea, M.; Clavel, A.; Doiz, O.; Castillo, F.J.; Rubio, M.C.; Gómez-Lus, R. Fuchsin fluorescence and autofluorescence in Cryptosporidum, Isospora and Cyclospora oocysts. Int. J. Parasitol. 1998, 28, 1881–1883. [Google Scholar] [CrossRef]
- Ortega, Y.R.; Sterling, C.R. Cyclospora cayetanensis: Epidemiology and diagnosis. Clin. Microbiol. Newsl. 1996, 18, 169–172. [Google Scholar] [CrossRef]
- Relman, D.A.; Schmidt, T.M.; Gajadhar, A.; Sogin, M.; Cross, J.; Yoder, K.; Sethabutr, O.; Echeverria, P. Molecular phylogenetic analysis of Cyclospora, the human intestinal pathogen, suggests that it is closely related to Eimeria species. J. Infect. Dis. 1996, 173, 440–445. [Google Scholar]
- Olivier, C.; van de Pas, S.; Lepp, P.W.; Yoder, K.; Relman, D.A. Sequence variability in the first internal transcribed spacer region within and among Cyclospora species is consistent with polyparasitism. Int. J. Parasitol. 2001, 31, 1475–1487. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Torres, P.; Simpson, S.; Kerdahi, K.; Ortega, Y. Sequence characterization of heat shock protein gene of Cyclospora cayetanensis isolates from Nepal, Mexico, and Peru. J. Parasitol. 2013, 99, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Varma, M.; Hester, J.D.; Schaefer, F.W., III; Ware, M.W.; Lindquist, H.D.A. Detection of Cyclospora cayetanensis using a quantitative real-time PCR assay. J. Microbiol. Methods 2003, 53, 27–36. [Google Scholar] [CrossRef]
- Verweij, J.J.; Laeijendecker, D.; Brienen, E.A.T.; van Lieshout, L.; Polderman, A.M. Detection of Cyclospora cayetanensis in travellers returning from the tropics and subtropics using microscopy and real-time PCR. Int. J. Med. Microbiol. 2003, 293, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, M.; Verweij, J.J.; Sethabutr, O.; Bodhidatta, L.; Garcia, L.; Maro, A.; Kumburu, H.; Gratz, J.; Kibiki, G.; Houpt, E.R. Multiplex polymerase chain reaction method to detect Cyclospora, Cystoisospora, and Microsporidia in stool samples. Diagn. Microbiol. Infect. Dis. 2011, 71, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Lee, S.E.; Kim, T.S.; Ma, D.W.; Chai, J.Y.; Shin, E.H. Multiplex-Touchdown PCR to simultaneously detect Cryptosporidium parvum, Giardia lamblia, and Cyclospora cayetanensis, the major causes of traveler’s diarrhea. Korean J. Parasitol. 2016, 54, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Benedict, T.; Marcet, P.L.; Wiegand, R.E.; Herwaldt, B.L.; da Silva, A.J. Molecular detection of Cyclospora cayetanensis in human stool specimens using UNEX-based DNA extraction and real-time PCR. Parasitology 2018, 145, 865–870. [Google Scholar] [CrossRef]
- Murphy, H.R.; Cinar, H.N.; Gopinath, G.; Noe, K.E.; Chatman, L.D.; Miranda, N.E.; Wetherington, J.H.; Neal-McKinney, J.; Pires, G.S.; Sachs, E.; et al. Interlaboratory validation of an improved method for detection of Cyclospora cayetanensis in produce using a real-time PCR assay. Food Microbiol. 2018, 69, 170–178. [Google Scholar] [CrossRef]
- Shin, J.H.; Lee, S.E.; Kim, T.S.; Ma, D.W.; Cho, S.H.; Chai, J.Y.; Shin, E.H. Development of molecular diagnosis using Multiplex Real-Time PCR and T4 phage internal control to simultaneously detect Cryptosporidium parvum, Giardia lamblia, and Cyclospora cayetanensis from human stool samples. Korean J. Parasitol. 2018, 56, 419–427. [Google Scholar] [CrossRef]
- Buss, S.N.; Leber, A.; Chapin, K.; Fey, P.D.; Bankowski, M.J.; Jones, M.K.; Rogatcheva, M.; Kanack, K.J.; Bourzac, K.M. Multicenter evaluation of the BioFire FilmArray Gastrointestinal Panel for etiologic diagnosis of infectious gastroenteritis. J. Clin. Microbiol. 2015, 53, 915–925. [Google Scholar] [CrossRef]
- Hitchcock, M.M.; Hogan, C.A.; Budvytiene, I.; Banaei, N. Reproducibility of positive results for rare pathogens on the FilmArray GI Panel. Diagn. Microbiol. Infect. Dis. 2019, 95, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Wei-Pridgeon, Y.; Li, W.; Nascimento, F.S.; Bishop, H.S.; Herwaldt, B.L.; Moss, D.M.; Nayak, V.; Srinivasamoorthy, G.; Sheth, M.; et al. Draft genome sequences from Cyclospora cayetanensis oocysts purified from a human stool sample. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.S.; Wei-Pridgeon, Y.; Arrowood, M.J.; Moss, D.; da Silva, A.J.; Talundzic, E.; Qvarnstrom, Y. Evaluation of library preparation methods for Illumina next generation sequencing of small amounts of DNA from foodborne parasites. J. Microbiol. Methods 2016, 130, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, L.; Zheng, H.; Xu, Z.; Roellig, D.M.; Li, N.; Frace, M.A.; Tang, K.; Arrowood, M.J.; Moss, D.M.; et al. Comparative genomics reveals Cyclospora cayetanensis possesses coccidia-like metabolism and invasion components but unique surface antigens. BMC Genom. 2016, 17, 316. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Guo, Y.; Zhang, L.; Rowe, L.A.; Roellig, D.M.; Frace, M.A.; Li, N.; Liu, S.; Feng, Y.; Xiao, L. Genetic similarities between Cyclospora cayetanensis and cecum-infecting avian Eimeria spp. in apicoplast and mitochondrial genomes. Parasit. Vectors 2015, 8, 358. [Google Scholar] [CrossRef]
- Ogedengbe, M.E.; Qvarnstrom, Y.; da Silva, A.J.; Arrowood, M.J.; Barta, J.R. A linear mitochondrial genome of Cyclospora cayetanensis (Eimeriidae, Eucoccidiorida, Coccidiasina, Apicomplexa) suggests the ancestral start position within mitochondrial genomes of eimeriid coccidia. Int. J. Parasitol. 2015, 45, 361–365. [Google Scholar] [CrossRef]
- Cinar, H.N.; Gopinath, G.; Jarvis, K.; Murphy, H.R. The complete mitochondrial genome of the foodborne parasitic pathogen Cyclospora cayetanensis. PLoS ONE 2015, 10, e0128645. [Google Scholar] [CrossRef]
- Cinar, H.N.; Qvarnstrom, Y.; Wei-Pridgeon, Y.; Li, W.; Nascimento, F.S.; Arrowood, M.J.; Murphy, H.R.; Jang, A.; Kim, E.; Kim, R.; et al. Comparative sequence analysis of Cyclospora cayetanensis apicoplast genomes originating from diverse geographical regions. Parasit. Vectors 2016, 9, 611. [Google Scholar] [CrossRef]
- Gopinath, G.R.; Cinar, H.N.; Murphy, H.R.; Durigan, M.; Almeria, M.; Tall, B.D.; DaSilva, A.J. A hybrid reference-guided de novo assembly approach for generating Cyclospora mitochondrion genomes. Gut Pathog. 2018, 10, 15. [Google Scholar] [CrossRef]
- Nascimento, F.S.; Barta, J.R.; Wale, J.; Hofstetter, J.N.; Casillas, S.; Barratt, J.; Talundzic, E.; Arrowood, M.J.; Qvarnstrom, Y. Mitochondrial junction region as genotyping marker for Cyclospora cayetanensis. Emerg. Infect. Dis. 2019, 25, 1307–1312. [Google Scholar] [CrossRef]
- Barratt, J.L.N.; Park, S.; Nascimento, F.S.; Hofstetter, J.; Plucinski, M.; Casillas, S.; Bradbury, R.S.; Arrowood, M.J.; Qvarnstrom, Y.; Talundzic, E. Genotyping genetically heterogeneous Cyclospora cayetanensis infections to complement epidemiological case linkage. Parasitology 2019, 146, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Roellig, D.M.; Li, N.; Tang, K.; Frace, M.; Ortega, Y.; Arrowood, M.J.; Feng, Y.; Qvarnstrom, Y.; Wang, L.; et al. Multilocus sequence typing tool for Cyclospora cayetanensis. Emerg. Infect. Dis. 2016, 22, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, N.; Ortega, Y.R.; Zhang, L.; Roellig, D.M.; Feng, Y.; Xiao, L. Population genetic characterization of Cyclospora cayetanensis from discrete geographical regions. Exp. Parasitol. 2018, 184, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, Y.; Wang, X.; Zhang, L.; Ortega, Y.; Feng, Y. Mitochondrial genome sequence variation as a useful marker for assessing genetic heterogeneity among Cyclospora cayetanensis isolates and source-tracking. Parasit. Vectors 2019, 12, 47. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, J.N.; Nascimento, F.S.; Park, S.; Casillas, S.; Herwaldt, B.L.; Arrowood, M.J.; Qvarnstrom, Y. Evaluation of multilocus sequence typing of Cyclospora cayetanensis based on microsatellite markers. Parasite 2019, 26, 3. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Lee, M.M.; Murphy, H.R. Use of a common laboratory glassware detergent improves recovery of Cryptosporidium parvum and Cyclospora cayetanensis from lettuce, herbs and raspberries. Int. J. Food Microbiol. 2012, 153, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Chandra, V.; Torres, M.; Ortega, Y.R. Efficacy of wash solutions in recovering Cyclospora cayetanensis, Cryptosporidium parvum, and Toxoplasma gondii from basil. J. Food Prot. 2014, 77, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.F.; Gajadhar, A.A. Optimization and validation of methods for isolation and real-time PCR identification of protozoan oocysts on leafy green vegetables and berry fruits. Food Waterborne Parasitol. 2016, 2, 1–7. [Google Scholar] [CrossRef]
- Robertson, L.J.; Gjerde, B.; Campbell, A.T. Isolation of Cyclospora oocysts from fruits and vegetables using lectin-coated paramagnetic beads. J. Food Prot. 2000, 63, 1410–1414. [Google Scholar] [CrossRef]
- Steele, M.; Unger, S.; Odumeru, J. Sensitivity of PCR detection of Cyclospora cayetanensis in raspberries, basil, and mesclun lettuce. J. Microbiol. Methods 2003, 54, 277–280. [Google Scholar] [CrossRef]
- Lalonde, L.F.; Gajadhar, A.A. Detection and differentiation of coccidian oocysts by real-time PCR and melting curve analysis. J. Parasitol. 2011, 97, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.R.; Almeria, S.; da Silva, A.J. BAM 19b: Molecular Detection of Cyclospora Cayetanensis in Fresh Produce Using Real-Time PCR. Available online: https://www. fda. gov/food/laboratory-methods-food-safety/bam-19b-molecular-detection-Cyclospora-cayetanensis-fresh-produce-using-real-time-pcr (accessed on 14 August 2018).
- Medina-De la Garza, C.E.; García-López, H.L.; Salinas-Carmona, M.C.; González-Spencer, D.J. Use of discontinuous Percoll gradients to isolate Cyclospora oocysts. Ann. Trop. Med. Parasitol. 1997, 91, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Riner, D.K.; Mullin, A.S.; Lucas, S.Y.; Cross, J.H.; Lindquist, H.D.A. Enhanced concentration and isolation of Cyclospora cayetanensis oocysts from human fecal samples. J. Microbiol. Methods 2007, 71, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Wei-Pridgeon, Y.; Van Roey, E.; Park, S.; Srinivasamoorthy, G.; Nascimento, F.S.; Moss, D.M.; Talundzic, E.; Arrowood, M.J. Purification of Cyclospora cayetanensis oocysts obtained from human stool specimens for whole genome sequencing. Gut Pathog. 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Dalton, C.; Goater, A.D.; Pethig, R.; Smith, H.V. Viability of Giardia intestinalis cysts and viability and sporulation state of Cyclospora cayetanensis oocysts determined by electrorotation. Appl. Environ. Microbiol. 2001, 67, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Guerrant, R.L.; Van Gilder, T.; Steiner, T.S.; Thielman, N.M.; Slutsker, L.; Tauxe, R.V.; Hennessy, T.; Griffin, P.M.; DuPont, H.; Bradley Sack, R.; et al. Practice guidelines for the management of infectious diarrhea. Clin. Infect. Dis. 2001, 32, 331–350. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, S.M.; Schuetz, A.N.; Franco-Paredes, C. Efficacy of nitazoxanide for cyclosporiasis in patients with sulfa allergy. Clin. Infect. Dis. 2007, 44, 466–467. [Google Scholar] [CrossRef] [PubMed]
- Diaz, E.; Mondragon, J.; Ramirez, E.; Bernal, R. Epidemiology and control of intestinal parasites with nitazoxanide in children in Mexico. Am. J. Trop. Med. Hyg. 2003, 68, 384–385. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, M.B.; Lee, E.H. Coccidial contamination of raspberries: Mock contamination with Eimeria acervulina as a model for decontamination treatment studies. J. Food Prot. 2001, 64, 1854–1857. [Google Scholar] [CrossRef]
- Kniel, K.E.; Shearer, A.E.H.; Cascarino, J.L.; Wilkins, G.C.; Jenkins, M.C. High hydrostatic pressure and UV light treatment of produce contaminated with Eimeria acervulina as a Cyclospora cayetanensis surrogate. J. Food Prot. 2007, 70, 2837–2842. [Google Scholar] [CrossRef]
- El Zawawy, L.A.; El-Said, D.; Ali, S.M.; Fathy, F.M. Disinfection efficacy of sodium dichloroisocyanurate (NADCC) against common food-borne intestinal protozoa. J. Egypt. Soc. Parasitol. 2010, 40, 165–185. [Google Scholar] [PubMed]
- Hussein, E.M.; Ahmed, S.A.; Mokhtar, A.B.; Elzagawy, S.M.; Yahi, S.H.; Hussein, A.M.; El-Tantawey, F. Antiprotozoal activity of magnesium oxide (MgO) nanoparticles against Cyclospora cayetanensis oocysts. Parasitol. Int. 2018, 67, 666–674. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Location | % (No. Positive/Total Samples Analyzed) | Food Type | References |
|---|---|---|---|
| Cambodia | 8.3 (3/36) | Water spinach | [126] |
| Costa Rica | 8.0 (2/25) | Lettuce | [127] |
| Canada | 1.7 (9/544) | Precut salads and leafy greens | [128] |
| Canada | 0.5 (6/1171) | Arugula/baby arugula, baby spinach and spring mix | [129] |
| Egypt | 16 (4/25) | Lettuce heads | [130] |
| Egypt | 21.3 (64/300) | Rocket, lettuce, parsley, leek, green onion | [131] |
| Egypt | 25.7 (9/35) | Fresh strawberry juice | [132] |
| Ethiopia | 6.9 (25/360) | Vegetables and raw fruits (avocado, lettuce, cabbage, carrot, tomato, banana and mango) * | [133] |
| Ghana | 5.1 (20/395) | Cabbage, green pepper, onion, tomato, lettuce | [134] |
| Italy | 12.2 (6/49) | Fennel, cucumber, celery, tomato | [135] |
| Italy | 1.3 (8/648) | Ready to eat-packaged salads | [136] |
| Korea | 1.2 (5/404) | Winter grown cabbage, sprouts, blueberries, and cherry tomatoes | [137] |
| Nepal | Number not indicated | Cabbage, lettuce, mustard leaves | [71] |
| Nepal | Number not indicated | Lettuce, spinach, mustard and basil leaves | [76] |
| Peru | Two surveys: 1.8 (2/110) and 1.6 (1/62) | Lettuce, mint, black mint | [138] |
| Venezuela | 5.9% (6/102) | Lettuce | [139] |
| Vietnam | 8.4 (24/287) | Herbs (basil, coriander sativum and coriander, Vietnamese mint, marjoram, persicaria) and lettuce | [69] |
| Country | Year | Disease Cases (If Laboratory Confirmed: Number of Cases) | Source of Exposure Suspected or Known | Origin/Notes | References |
|---|---|---|---|---|---|
| Australia (cruise) | 2010 | 266 (73) | Lettuce (suspected) | Two consecutive voyages | [144] |
| Brazil | 1999–2002 | 132 (16) | Not indicated | Not available | [149] |
| Canada (2 provinces) and USA (20 states) | 1996 | 1465 (978) | Raspberries | Guatemala | [150,151,152] |
| Canada (Ontario) and USA (14 states) | 1997 | 1012 (422) | Raspberries | Guatemala | [150,153] |
| Canada (Ontario) | 1998 | 315 | Raspberries | Guatemala | [99,150] |
| Canada (Ontario) | 1999 | 104 | Dessert berries (blackberries suspected) | Blackberries (Guatemala), strawberries (USA), frozen raspberries (Chile) | [99,150] |
| Canada (BC &) | 1999 | 15 | Undetermined | Undetermined | [150] |
| Canada (BC) | 2001 | 30 (17) | Thai basil | Imported from Thailand (supplier in USA). Not confirmed | [142] |
| Canada (BC) | 2003 | 11 | Cilantro | Undetermined, community | [154] |
| Canada (BC) | 2004 | 17 (9) | Mango or basil (suspected) | Not confirmed, community | [154] |
| Canada (BC) | 2004 | 8 | Cilantro suspected | Undetermined, community | [154] |
| Canada (Quebec) | 2005 | 250 (142) | Fresh basil | Mexico, workers who ate in a restaurant | [155] |
| Canada (Ontario) | 2005 | 44 (16) | Basil (suspected) | Retreat. Pasta salad with basil, origin not confirmed (shipment from Peru and Costa Rica) | [154] |
| Canada (BC) | 2006 | 28 | Basil or garlic | Undetermined | [154] |
| Canada (BC) | 2007 | 29 (14) | Organic basil | Mexico | [156] |
| Canada (Sarnia) | 2010 | 210 | Basil, pesto | Fundraiser event | [19] |
| Canada | 2013 | 25 | Leafy greens suspected | Undetermined | [157] |
| Canada (multiple provinces) | 2015 | 97 | Undetermined, multistate (BC: 5, Alberta: 1, Ontario: 84, Quebec: 7) | Travelers from Mexico | [115,158] |
| Canada (multiple provinces) | 2016 | 87 | Undetermined | BC:2, Alberta:2, Ontario 75, Quebec: 8 | [158] |
| Canada (multiple provinces) | 2017 | 164 | Undetermined | BC (17), Ontario (143), Quebec (3) and Nova Scotia (1) | [158] |
| Colombia | 2002 | 56 (31) | Salad, juices | University employees | [159] |
| Germany | 2000 | 34 | Salad, green leafy herbs | France, Italy, Germany | [94] |
| Nepal | 1989 | 535 (55) | Drinking water suspected | Foreigners, Travelers from UK | [160] |
| Nepal | 1989 | 14 (12) | Drinking water suspected | Foreigners | [161] |
| Nepal | 1992 | 964 (108) | Drinking water suspected | Foreigners | [40] |
| Mexico | 2001 | 97 (55/70 fecal samples analyzed) | Watercress (salad berros) | Party attendees at wedding and christening | [162] |
| Peru | 2004 | 127/274 people with diarrhea, 24/77 positive by microscopy | Undetermined (salsa sauces suspected) | Recruits | [90] |
| Peru | 2005 | 52 recruits (37 positive, 15 control) 20/35 positive by PCR | Undetermined (food, water) | Recruits | [89] |
| Poland | 2013 | 3 | Drinking water suspected | Travelers from Indonesia | [118] |
| Spain | 2003 | 13 (7) | Raspberry juice suspected | Travelers from Guatemala | [113] |
| Sweden | 2009 | 18 (12) | Snap peas | Guatemala | [143] |
| The Netherlands | 2001 | 29 (14) | Could not investigate potential food sources | Dutch participants at a scientific meeting in Bogor, Indonesia | [121] |
| Turkey | 2005 | 35 | Drinking water suspected | Undetermined | [163] |
| Turkey | 2007 | 505 stools (14/17 positive by PCR) | Undetermined, suspected to be related to lack of rain and use of contaminated water | Unwashed green salad suspected | [164] |
| United Kingdom | 2015 | 79 | Undetermined | Travelers from Mexico | [115] |
| USA (Illinois) | 1990 | 21 | Tap water or food | Not completely clarified | [165] |
| USA (Florida) | 1995 | 38 | Raspberries suspected, risk factor soil contact | (Guatemala as possible source) | [166] |
| USA (New York) | 1995 | 32 | Fruit suspected | Undetermined | [99] |
| USA (Massachusetts) | 1996 | 57 (12) | Berries | Wedding, strawberries (California), blueberries (Florida), blackberries (Guatemala), raspberries (Guatemala/Chile) | [167] |
| USA (Florida) | 1996 | 86 | Raspberries (suspected) | Guatemala | [168] |
| USA (South Carolina) | 1996 | 38 | Raspberries | Guatemala | [169] |
| USA (20 states) and Canada (2 provinces) | 1996 | 1465 (978) | Raspberries | Guatemala | [70,150,151,152,154] |
| USA (Florida), | 1997 | 220 (including people in cruise ship that departed from Florida) | Mesclun | Peru or USA (If related to 12 cases from Florida above, then mesclun from Peru most probable source) | [99] |
| USA (14 states) and Canada (Ontario) | 1997 | 1012 (422) | Raspberries | Guatemala | [150,153] |
| USA (Northern Virginia, Washington DC-Baltimore metropolitan area) | 1997 | 341 (48) | Basil | Multiple possible sources, may be local contamination | [99] |
| USA (Virginia) | 1997 | 21 | Fruit plate | Undetermined | [99] |
| USA (Georgia) | 1998 | 17 | Probable fruit salad | Undetermined | [99] |
| USA (Florida) | 1999 | 94 | Berries likely | Undetermined | [99] |
| USA (Missouri) | 1999 | 62 | Basil in chicken pasta and tomato basil salad | Mexico or USA, two events | [170] |
| USA (Georgia) | 2000 | 19 | Raspberries and/or blackberries (suspected) | Suspected to be from Guatemala | [171] |
| USA (Pennsylvania) | 2000 | 54 | Raspberry, wedding cake | Guatemala | [141] |
| USA (Florida) | 2001 | 39 | Undetermined | Undetermined | [171] |
| USA (New York City) | 2001 | 3 | Undetermined | Undetermined | [171] |
| USA (Vermont) | Dec 2001-Jan 2002 | 22 | Raspberry (likely) | Suspected to be from Chile | [171] |
| USA (Massachusetts) | 2002 | 8 | Undetermined | Undetermined | [171] |
| USA (New York) | 2002 | 14 | Undetermined | Undetermined | [171] |
| USA (Texas, Illinois) | 2004 | 95 (38 in Texas, 57 in Illinois) | Undetermined | Undetermined, basil likely | [3,171] |
| USA (Tennessee) | 2004 | 12 | Undetermined | Undetermined | [171] |
| USA (Pennsylvania) | 2004 | 96 | Snow peas | Guatemala | [172] |
| USA (Florida) | 2005 | 582 | Basil, restaurants | Peru | [19,171] |
| USA (South Carolina) | 2005 | 6 | Undetermined | Undetermined | [171] |
| USA (Massachusetts) | 2005 | 74 | Two different outbreaks (58 and 16 cases) | Undetermined | [171] |
| USA (Connecticut) | 2005 | 30 | Basil suspected | Undetermined | [171] |
| USA (Minnesota) | 2006 | 14 | Undetermined | Undetermined | [171] |
| USA (New York) | 2006 | 20 | Undetermined | Undetermined | [171] |
| USA (Georgia) | 2006 | 3 | Undetermined | Undetermined | [171] |
| USA (Wisconsin) | 2008 | 4 | Sugar snap peas (likely) | Guatemala not confirmed | [171] |
| USA (California) | 2008 | 45 | Raspberries and/or blackberries (likely) | Undetermined | [171] |
| USA (District of Columbia) | 2009 | 34 | Undetermined | Undetermined | [171] |
| USA (Connecticut) | 2009 | 8 | Blackberries and raspberries | Undetermined | [171] |
| USA (Florida) | 2011 | 12 | Undetermined | Undetermined | [171] |
| USA (Georgia) | 2011 | 88* | Undetermined | Undetermined | [171] |
| USA (Texas) | 2012 | 16 | Undetermined | Undetermined | [171] |
| USA (Iowa, Nebraska and neighboring states) | 2013 | 162 | Bagged salad mix | Mexico | [171] |
| USA (Texas) | 2013 | 270 (38) | Cilantro | Mexico, multistate | [173] |
| USA (Wisconsin) | 2013 | 8 | Berry salad (suspected) | Undetermined | [171] |
| USA (Michigan) | 2014 | 14 | Undetermined | Undetermined | [171] |
| USA (Iowa, Nebraska) | 2014 | 227 (161) | Lettuce (imported romaine lettuce) | Mexico | [146,147] |
| USA (Texas) | 2014 | 304 (26) | Cilantro | Mexico | [171] |
| USA (South Carolina) | 2014 | 13 | Undetermined | Undetermined | [171] |
| USA (31 states) | 2015 | 546 (90 cases in Georgia, Texas and Wisconsin) | Cilantro (suspected) | Mexico | [171] |
| USA (Texas) | 2016 | 6 ** | Carrots or green cabbage in coleslaw (suspected) | Undetermined | [171,174] |
| USA (40 states) | 2017 (summary) | 1065 | Undetermined | Undetermined | [171,174] |
| USA (Florida) | 2017 | 6 | Berries (suspected) | Undetermined | [171] |
| USA (Texas) | 2017 | 38 *** | Scallions (i.e., green onions) | Undetermined | [171] |
| USA (Michigan) | 2017 | 29 | Undetermined | Undetermined | [171] |
| USA (Tennessee) | 2017 | 4 ††† | Undetermined | Undetermined | [171] |
| USA (Connecticut) | 2017 | 3 | Undetermined | Undetermined | [171] |
| USA (Florida) | 2017 | 3 ‡‡‡ | Undetermined | Undetermined | [171] |
| USA (33 states) | 2018 (summary) | 2299 total cases (partial data indicated in a, b, c and d) | Many cases not directly linked, rest indicated in a, b, c and d cases | [175,176] | |
| USA (4 states) a | 2018 | 250 (250) | Vegetable trays with broccoli, cauliflower, carrots and dill dip | Bought in supermarkets. It was not possible to determine if an individual component of the vegetable trays was the likely vehicle of infection | [177] |
| USA (15 states and New York) b | 2018 | 511 (511) | Romaine lettuce and carrot mix | Salads purchased from a fast restaurant chain | [178] |
| USA (2 states) c | 2018 | 8 (8) | Basil | Undetermined | [175] |
| USA (Midwest- 3 clusters) d | 2018 | 53 (53) | Cilantro | Mexican-style restaurants | [175] |
| USA (30 states) & | 2019 | 1,696 (1,696) | Basil (205) | Mexico | [175] |
| Country | Type of Water | Percentage (Positive/Total Analyzed) | Reference |
|---|---|---|---|
| Cambodia | Water spinach | 8.3 (3/36) | [126] |
| Egypt | Household water tanks in Alexandria | 9.0 (9/100) | [183] |
| Egypt | Finished piped water, irrigation canals, shallow underground water and drain water | positive detection from 5 residential areas. Number not indicated | [72] |
| Egypt | Treated potable water from tanks | 0.2 (2/840) | [184] |
| Egypt | River Nile, water works, water pumps, water tank, pond and canals | 5.9 (20/336) | [185] |
| Ghana | Sachet drinking water | 59.3 (16/27) | [186] |
| Guatemala | Rivers | 6.7 (2/30) | [58] |
| Guatemala | Drinking water sources | 41.7 (5/12) * | [187] |
| Italy | Tap water | 30.0 (3/10) * | [135] |
| Italy | Treated wastewater | 21.3 (20/94) * | [188] |
| Italy | Well water | 6.2 (1/16) * | [188] |
| Malaysia | (1) Drinking water treatment plants and (2) Recreational water (man-made lake) | (1) 8.3 (2/24) and (2) 16.7 (2/12) | [189] |
| Nepal | Chlorinated water | presence of oocysts | [161] |
| Nepal | Sewage contamination | 22.2 (4/18); 41.7 (5/12) | [15,71] |
| Nepal | Irrigation water, pond water, tap water and tube wells water | 2 of 8 irrigation canals, 1 of 12 pond water samples (none in tap water or tube wells) | [76] |
| Peru | Wastewater | 72.7 (8/11) | [182] |
| Spain | DWTP a, WWTP b, rivers | 9.0 (20/223) | [190] |
| Tunisia | Wastewater | 0.4 (1/232) * | [191] |
| Turkey | (1) Streams and (2) drinking water | Total: 24.6 (56/228) (1) 31.1 (56/180), (2) 0.0 (0/48) | [192] |
| USA | WWTP b (1) influent and (2) effluent | (1) 25.0 (6/24) (2) 12.5(3/24) * | [193] |
| Vietnam | Lakes and rivers | 63.6 (84/132) * | [194] |
| Vietnam | Water samples from markets and farms | 12.6 (12 /95) | [69] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeria, S.; Cinar, H.N.; Dubey, J.P. Cyclospora cayetanensis and Cyclosporiasis: An Update. Microorganisms 2019, 7, 317. https://doi.org/10.3390/microorganisms7090317
Almeria S, Cinar HN, Dubey JP. Cyclospora cayetanensis and Cyclosporiasis: An Update. Microorganisms. 2019; 7(9):317. https://doi.org/10.3390/microorganisms7090317
Chicago/Turabian StyleAlmeria, Sonia, Hediye N. Cinar, and Jitender P. Dubey. 2019. "Cyclospora cayetanensis and Cyclosporiasis: An Update" Microorganisms 7, no. 9: 317. https://doi.org/10.3390/microorganisms7090317
APA StyleAlmeria, S., Cinar, H. N., & Dubey, J. P. (2019). Cyclospora cayetanensis and Cyclosporiasis: An Update. Microorganisms, 7(9), 317. https://doi.org/10.3390/microorganisms7090317