Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and CAR Genes Transcript Levels to Counteract the Pro-Oxidant Effect of Hydrogen Peroxide


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Media
2.2. Yeast Cultivation
2.3. Carotenoid Extraction
2.4. Biochemical Methods
2.5. RT-PCR
2.6. Data Analysis
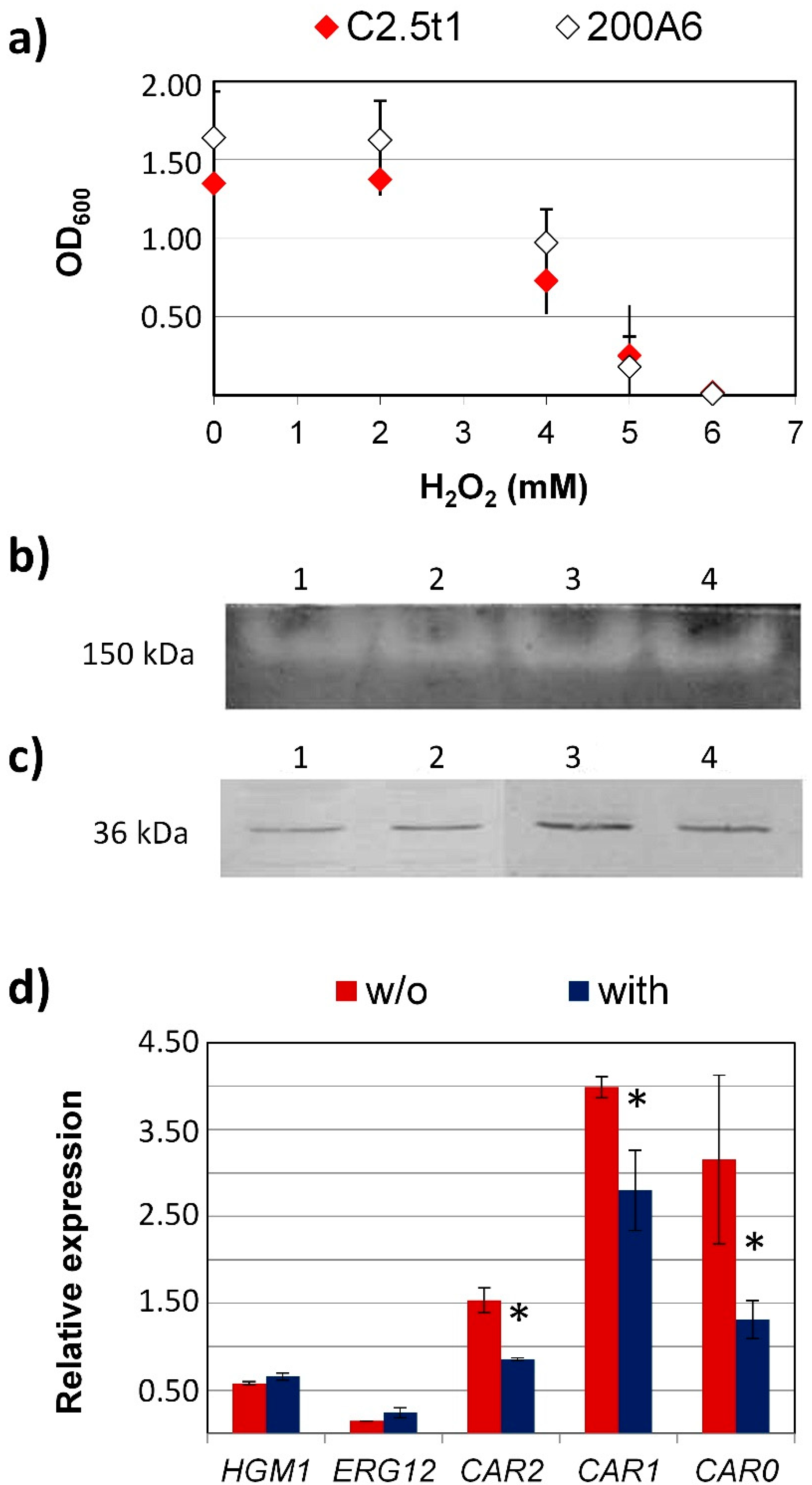
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, E.A.; Schroeder, W.A. Microbial carotenoids. Adv. Biochem. Eng. Biotechnol. 1996, 53, 119–178. [Google Scholar] [PubMed]
- Vershinin, A. Biological functions of carotenoids-diversity and evolution. Biofactors 1999, 10, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Emodi, A. Carotenoids. Properties and applications. Food Technol. 1978, 32, 38–40. [Google Scholar]
- Moliné, M.; Libkind, D.; del Carmen Diéguez, M.; van Broock, M. Photoprotective role of carotenoids in yeasts: Response to UV-B of pigmented and naturally-occurring albino strains. J. Photochem. Photobiol. B Biol. 2009, 95, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Hennekens, C.H. β-Carotene supplementation and cancer prevention. Nutrition 1997, 13, 697–699. [Google Scholar] [CrossRef]
- Moore, M.M.; Breedveld, M.W.; Autor, A.P. The role of carotenoids in preventing oxidative damage in the pigmented yeast Rhodotorula mucilaginosa. Arch. Biochem. Biophys. 1989, 270, 419–431. [Google Scholar] [CrossRef]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as antioxidants: A review. J. Photochem. Photobiol. B 1997, 41, 189–200. [Google Scholar] [CrossRef]
- Mendez-Alvarez, S.; Rüfenacht, K.; Eggen, R.I.L. The oxidative stress-sensitive yap1 null strain of Saccharomyces cerevisiae becomes resistant due to increased carotenoid levels upon the introduction of the Chlamydomonas reinhardtii cDNA, coding for the 60S ribosomal protein L10a. Biochem. Biophys. Res. Commun. 2000, 267, 953–959. [Google Scholar] [CrossRef]
- Yan, G.L.; Liang, H.Y.; Wang, Z.Q.; Yang, X.F.; Liu, D.; Liu, J.F.; Duan, C.Q. Important role of catalase in the production of β-carotene by recombinant Saccharomyces cerevisiae under H2O2 stress. Curr. Microbiol. 2011, 62, 1056–1061. [Google Scholar] [CrossRef]
- Nanou, K.; Roukas, T. Oxidative stress response and morphological changes of Blakeslea trispora induced by butylated hydroxytoluene during carotene production. Appl. Biochem. Biotech. 2010, 160, 2415–2423. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Koci, R.; Breierova, E. Production of carotenoid/ergosterol supplemeted biomass by red yeast Rhodotorula glutinis grown under external stress. Food Technol. Biotech. 2010, 48, 56–61. [Google Scholar]
- Irazusta, V.; Nieto-Penalver, C.G.; Cabral, M.E.; Amoroso, M.J.; de Figueroa, L.I.C. Relationship among carotenoid production, copper bioremediation and oxidative stress in Rhodotorula mucilaginosa RC-11. Proc. Biochem. 2013, 48, 803–809. [Google Scholar] [CrossRef]
- Lee, J.; Koo, N.; Min, D.B. Reactive Oxygen Species, aging, and antioxidative nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2004, 3, 21–33. [Google Scholar] [CrossRef]
- Ukibe, K.; Katsuragi, T.; Tani, Y.; Takagi, H. Efficient screening for astaxanthin overproducing mutants of the yeast Xanthophyllomyces dendrorhous by flow cytometry. FEMS. Microbiol. Lett. 2008, 286, 241–248. [Google Scholar] [CrossRef]
- Amengual, J.; Lobo, G.P.; Golczak, M.; Li, H.N.; Klimova, T.; Hoppel, C.L.; Wyss, A.; Palczewski, K.; von Lintig, J. A mitochondrial enzyme degrades carotenoids and protects against oxidative stress. Faseb J. 2011, 25, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Gruszecki, W.I.; Strzalka, K. Carotenoids as modulators of lipid membrane physical properties. Biochim. Biophys. Acta 2005, 1740, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verwaal, R.; Jiang, Y.; Wang, J.; Daran, J.M.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. Heterologous carotenoid production in Saccharomyces cerevisiae induces the pleiotropic drug resistance stress response. Yeast 2010, 27, 983–998. [Google Scholar] [CrossRef]
- Aksu, Z.; Eren, A.T. Carotenoids production by the yeast Rhodotorula mucilaginosa: use of agricultural wastes as carbon source. Process Biochem. 2005, 40, 2985–2991. [Google Scholar] [CrossRef]
- Cutzu, R.; Coi, A.; Rosso, F.; Bardi, L.; Ciani, M.; Budroni, M.; Zara, G.; Zara, S.; Mannazzu, I. From crude glycerol to carotenoids by using a Rhodotorula glutinis mutant. World J. Microbiol. Biotechnol. 2013, 29, 1009–1017. [Google Scholar] [CrossRef]
- Singh, P.; Tsuji, M.; Singh, S.M.; Roy, U.; Hoshino, T. Taxonomic characterization, adaptation strategies and biotechnological potential of cryophilic yeasts from ice cores of Midre Lovénbreen glacier, Svalbard, Arctic. Cryobiology 2013, 66, 167–175. [Google Scholar] [CrossRef]
- Kawahara, H.; Hirai, A.; Minabe, T.; Obata, H. Stabilization of astaxanthin by a novel biosurfactant produced by Rhodotorula mucilaginosa KUGPP-1. Biocontrol Sci. 2013, 18, 21–28. [Google Scholar] [CrossRef]
- Li, R.; Zhang, H.; Liu, W.; Zheng, X. Biocontrol of postharvest gray and blue mold decay of apples with Rhodotorula mucilaginosa and possible mechanisms of action. Int. J. Food Microbiol. 2011, 146, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ge, L.; Chen, K.; Zhao, L.; Zhang, X. Enhanced biocontrol activity of Rhodotorula mucilaginosa cultured in media containing chitosan against postharvest diseases in strawberries: possible mechanisms underlying the effect. J. Agric. Food Chem. 2014, 62, 4214–4224. [Google Scholar] [CrossRef]
- Deligios, M.; Fraumene, C.; Abbondio, M.; Mannazzu, I.; Tanca, A.; Addis, M.F.; Uzzau, S. Draft genome sequence of Rhodotorula mucilaginosa, an emergent opportunistic pathogen. Genome Announc. 2015, 3. [Google Scholar] [CrossRef]
- Addis, M.F.; Tanca, A.; Abbondio, M.; Cutzu, R.; Biosa, G.; Pagnozzi, D.; Uzzau, S.; Mannazzu, I. Proteomic analysis of Rhodotorula mucilaginosa: Dealing with the issues of a non-conventional yeast. Yeast 2016, 33, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Landolfo, S.; Ianiri, G.; Camiolo, S.; Porceddu, A.; Mulas, G.; Chessa, R.; Zara, G.; Mannazzu, I. CAR gene cluster and transcript levels of carotenogenic genes in Rhodotorula mucilaginosa. Microbiology 2018, 164, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bocanegra, A.R.; Torres Muñoz, J.A. Astaxanthin hyperproduction by Phaffia rhodozyma (now Xanthophyllomyces dendrorhous) with crude coconut milk as sole source of energy. Appl. Microbiol. Biotechnol. 2004, 66, 249–252. [Google Scholar] [CrossRef]
- Mannazzu, I.; Guerra, E.; Strabbioli, R.; Pediconi, D.; Fatichenti, F. The vanadate tolerant yeast Hansenula polymorpha undergoes cellular reorganization during growth in, and recovery from, the presence of vanadate. Microbiology 1998, 144, 2589–2597. [Google Scholar] [CrossRef]
- Landolfo, S.; Politi, H.; Angelozzi, D.; Mannazzu, I. ROS accumulation and oxidative damage to cell structures in Saccharomyces cerevisiae wine strains during fermentation of high-sugar-containing medium. Biochim. Biophys. Acta 2008, 1780, 892–898. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Luck, H. Catalase. In Methods in Enzymatic Analysis; Bergmeyer, H.-U., Ed.; Academic Press: New York, NY, USA, 1963; pp. 885–888. [Google Scholar]
- Gamero-Sandemetrio, E.; Gómez-Pastor, R.; Matallana, E. Zymogram profiling of superoxide dismutase and catalase activities allows Saccharomyces and non-Saccharomyces species differentiation and correlates to their fermentation performance. Appl. Microbiol. Biotechnol. 2013, 97, 4563–4576. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Kim, D.R. pcr: An R package for quality assessment, analysis and testing of qPCR data. PeerJ 2018. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, G.W.; Reodica, M.; Davies, M.J.; Heeren, G.; Jarolim, S.; Pillay, B.; Breitenbach, M.; Higgins, V.J.; Dawes, I.W. Superoxide radicals have a protective role during H2O2 stress. Mol. Biol. Cell 2013, 24, 2876–2884. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide-dismutase: improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Jeong, J.C.; Lee, I.Y.; Kim, S.W.; Park, Y.H. Stimulation of β-carotene synthesis by hydrogen peroxide in Blakeslea trispora. Biotech. Lett. 1999, 21, 683–686. [Google Scholar] [CrossRef]
- Iigusa, H.; Yoshida, Y.; Hasunuma, K. Oxygen and hydrogen peroxide enhance light-induced carotenoid synthesis in Neurospora crassa. FEBS Lett. 2005, 579, 4012–4016. [Google Scholar] [CrossRef]
- Liu, Y.S.; Wu, J.Y. Hydrogen peroxide-induced astaxanthin biosynthesis and catalase activity in Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 2006, 73, 663–668. [Google Scholar] [CrossRef]
- Wang, G.Y.; Keasling, J.D. Amplification of HMG-CoA reductase production enhances carotenoid accumulation in Neurospora crassa. Metab. Eng. 2002, 4, 193–201. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, L.; Shang, F.; Yan, G. Enhanced production of β-carotene in recombinant Saccharomyces cerevisiae by inverse metabolic engineering with supplementation of unsaturated fatty acids. Process Biochem. 2016, 51, 568–577. [Google Scholar] [CrossRef]
- Harrison, P.J.; Bugg, T.D. Enzymology of the carotenoid cleavage dioxygenases: Reaction mechanisms, inhibition and biochemical roles. Arch. Biochem. Biophys. 2014, 544, 105–111. [Google Scholar] [CrossRef]
- Reyes, L.H.; Gomez, J.M.; Kao, K.K. Improving carotenoids production in yeast via adaptive laboratory evolution. Metab. Eng. 2014, 21, 26–33. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Molecular Target | Primer Name | Sequence |
|---|---|---|
| HMG1 | HMG1F | 5′-TCACGCTCCACTCGCTCAAC-3′ |
| HMG1R | 5′-CGAGGACAAGATGGGGTTGG-3′ | |
| ERG12 | ERG12F | 5′-CAGTCGGCGCAGGCGTTCTT-3′ |
| ERG12R | 5′-GGACGCCGTGCGAGTAGAGC-3′ | |
| CAR2 | CAR2F | 5′-CCTTCCTCGCCAACGCCTCT-3′ |
| CAR2R | 5′-CGTTGTTGGCGTACAGGAGG-3′ | |
| CAR1 | CAR1F | 5′-CGGTCCCTCGCTCTACCTCA-3′ |
| CAR1R | 5′-CCTTGTCCGGGAAGACGATG-3′ | |
| CAR0 | CAR0F | 5′-CCGTCGGGTACTACAGTCTC-3′ |
| CAR0R | 5′-TAGCTCAGATATGGCGGCAA-3′ | |
| ACT1 | ACT1AF | 5′-CGTTCAGATCCAGGCCGTCT-3′ |
| ACT1AR | 5′-CGGCAATGCAAACCCTTCAT-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landolfo, S.; Chessa, R.; Zara, G.; Zara, S.; Budroni, M.; Mannazzu, I. Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and CAR Genes Transcript Levels to Counteract the Pro-Oxidant Effect of Hydrogen Peroxide. Microorganisms 2019, 7, 316. https://doi.org/10.3390/microorganisms7090316
Landolfo S, Chessa R, Zara G, Zara S, Budroni M, Mannazzu I. Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and CAR Genes Transcript Levels to Counteract the Pro-Oxidant Effect of Hydrogen Peroxide. Microorganisms. 2019; 7(9):316. https://doi.org/10.3390/microorganisms7090316
Chicago/Turabian StyleLandolfo, Sara, Rossella Chessa, Giacomo Zara, Severino Zara, Marilena Budroni, and Ilaria Mannazzu. 2019. "Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and CAR Genes Transcript Levels to Counteract the Pro-Oxidant Effect of Hydrogen Peroxide" Microorganisms 7, no. 9: 316. https://doi.org/10.3390/microorganisms7090316