The Algal Symbiont Modifies the Transcriptome of the Scleractinian Coral Euphyllia paradivisa during Heat Stress

 ,
,  , and
, and 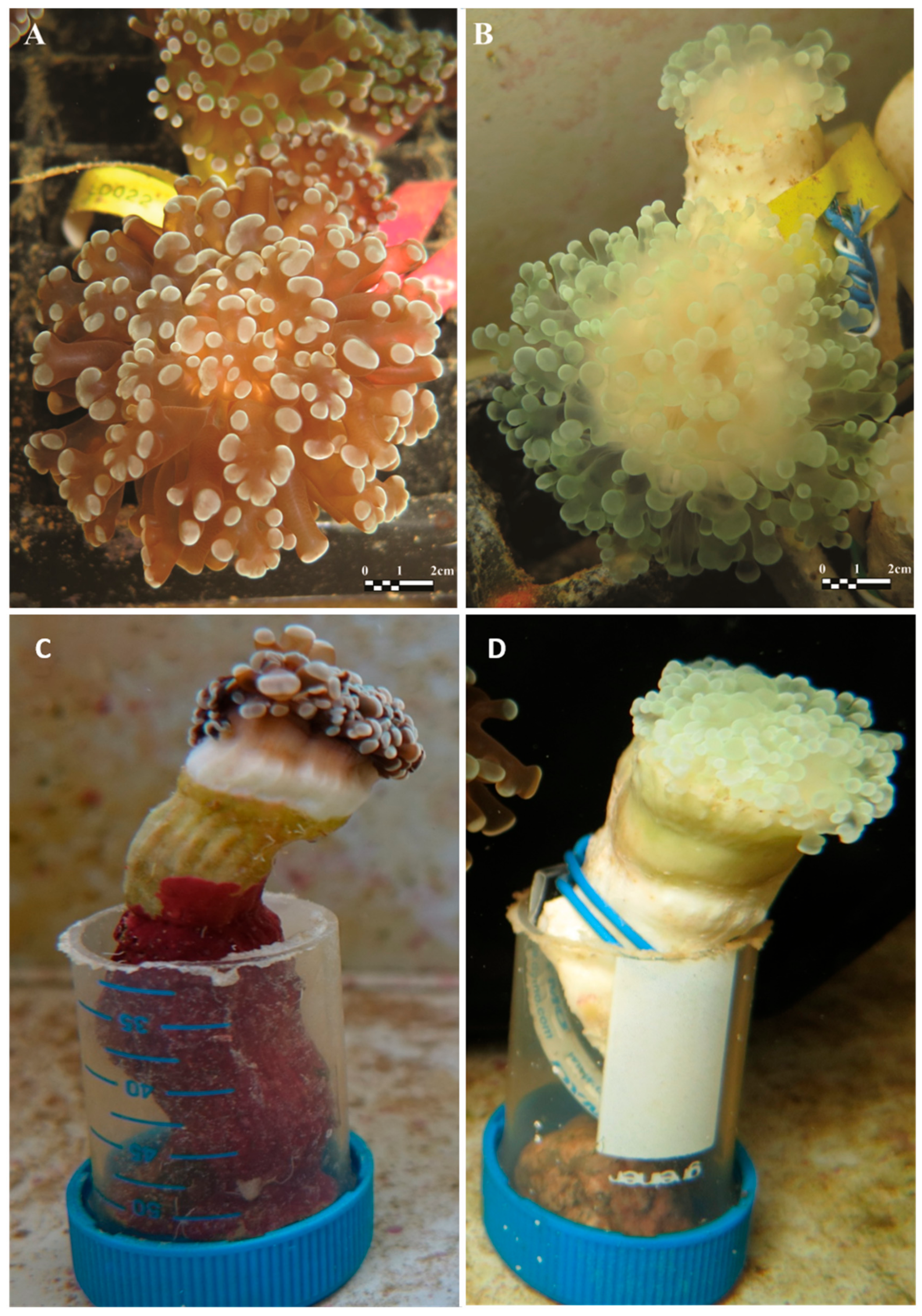{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Materials and Methods
2.1. Coral Sampling and Experimental Design
2.2. Physiological Measurements
2.3. RNA Extraction and NGS Analyses
2.4. Bioinformatics Analysis
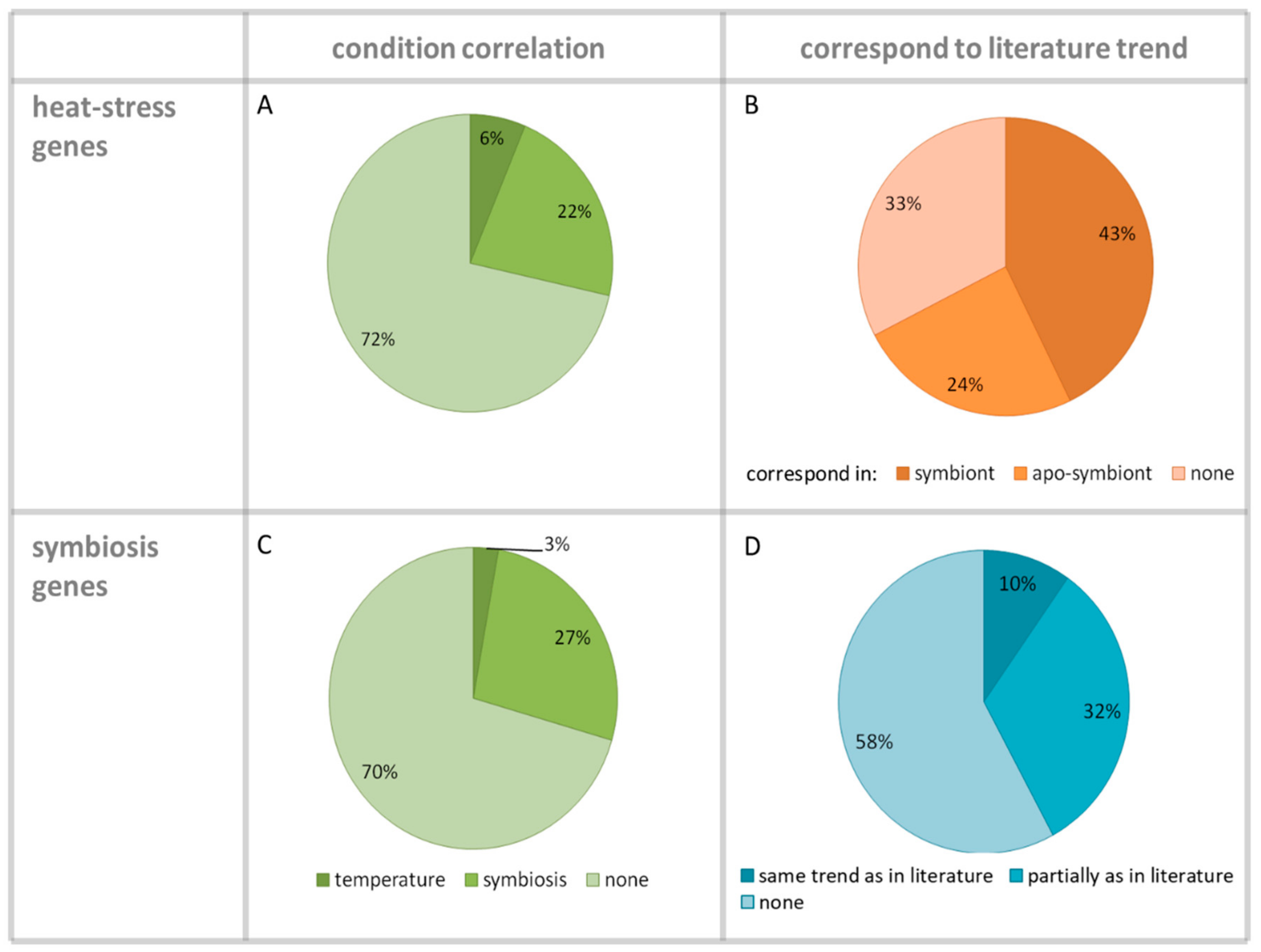
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farber, S.C.; Costanza, R.; Wilson, M.A. Economic and ecological concepts for valuing ecosystem services. Ecol. Econ. 2002, 41, 375–392. [Google Scholar] [CrossRef]
- Sebens, K. Biodiversity of Coral Reefs: What Are We Losing and Why? Am. Zool. 1994, 34, 115–133. [Google Scholar] [CrossRef]
- NOAA. Implementation of the National Coral Reef Action Strategy: Report on US Coral Reef Task Force Agency Activities from 2004 to 2006. Available online: https://www. coris. noaa. gov/activities/reportcongress2008/full_report. pdf. (accessed on 15 June 2006).
- The International Panel on Climate Change; 2014. Available online: https://www.ipcc.ch/site/assets/uploads/2018/05/SYR_AR5_FINAL_full_wcover.pdf (accessed on 1 November 2014).
- Stambler, N. Zooxanthellae: The yellow symbionts inside animals. In Coral Reefs: An Ecosystem in Transition; Stambler, N., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 87–106. ISBN 9789400701137. [Google Scholar]
- Muscatine, L.; Goiran, C.; Land, L.; Jaubert, J.; Cuif, J.-P.; Allemand, D. Stable isotopes (delta13C and delta15N) of organic matrix from coral skeleton. Proc. Natl. Acad. Sci. USA 2005, 102, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Muscatine, L.; Porter, J.W. Reef corals: Mutualistic symbioses adapted to nutrient-poor environments. Bioscience 1977, 27.7, 454–460. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Levitus, S.; Antonov, J.I.; Boyer, T.P.; Locarnini, R.A.; Garcia, H.E.; Mishonov, A.V. Global ocean heat content 1955–2008 in light of recently revealed instrumentation problems. Geophys. Res. Lett. 2009, 36, L07608. [Google Scholar] [CrossRef]
- Berkelmans, R.; van Oppen, M.J.H. The role of zooxanthellae in the thermal tolerance of corals: A “nugget of hope” for coral reefs in an era of climate change. Proc. Biol. Sci. 2006, 273, 2305–2312. [Google Scholar] [CrossRef]
- Done, T.; Whetton, P.; Jones, R.; Berkelmans, R.; Lough, J.; Skirving, W.; Wooldridge, S. Global Climate Change and Coral Bleaching on the Great Barrier Reef. In Final Report to the State of Queensland Greenhouse Taskforce through the Department of Natural Resources and Mines 49; Queensland Department of Natural Resources and Mines: Brisbane, Australia, 2003; Available online: http://www.academia.edu/download/44286335/Global_Climate_Change_and_Coral_Bleachin20160401-21979-nmu3ni.pdf (accessed on 24 June 2019).
- Maor-Landaw, K.; Levy, O. Survey of cnidarian gene expression profiles in response to environmental stressors: Summarizing 20 years of research, what are we heading for? In The Cnidaria, Past, Present and Future. The World of Medusa and Her Sisters; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 523–543. [Google Scholar]
- Lesser, M.P. Oxidative stress in marine environments: biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Lesser, M.P. Coral Bleaching: Causes and mechanisms. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer Science & Business Media: New York, NY, USA, 2011; pp. 405–419. [Google Scholar]
- DeSalvo, M.K.; Voolstra, C.R.; Sunagawa, S.; Schwarz, J.A.; Stillman, J.H.; Coffroth, M.A.; Szmant, A.M.; Medina, M. Differential gene expression during thermal stress and bleaching in the Caribbean coral Montastraea faveolata. Mol. Ecol. 2008, 17, 3952–3971. [Google Scholar] [CrossRef]
- Maor-Landaw, K.; Karako-Lampert, S.; Ben-Asher, H.W.; Goffredo, S.; Falini, G.; Dubinsky, Z.; Levy, O. Gene expression profiles during short-term heat stress in the Red Sea coral Stylophora pistillata. Glob. Chang. Biol. 2014, 20, 3026–3035. [Google Scholar] [CrossRef]
- Hahn, J.; Hu, Z.; Thiele, D.J.; Iyer, V.R. Genome-Wide Analysis of the Biology of Stress Responses through Heat Shock Transcription Factor. Mol. Cell. Biol. 2004, 24, 5249–5256. [Google Scholar] [CrossRef] [Green Version]
- Kültz, D. Evolution of the cellular stress proteome: from monophyletic origin to ubiquitous function. J. Exp. Biol. 2003, 206, 3119–3124. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef]
- Walter, P.; Ron, D. The unfolded protein response: from stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef]
- Brading, P.; Warner, M.E.; Davey, P.; Smith, D.J.; Achterberg, E.P.; Suggett, D.J. Differential effects of ocean acidification on growth and photosynthesis among phylotypes of Symbiodinium (Dinophyceae). Limnol. Oceanogr. 2011, 56, 927–938. [Google Scholar] [CrossRef]
- Kriegenburg, F.; Ellgaard, L.; Hartmann-Petersen, R. Molecular chaperones in targeting misfolded proteins for ubiquitin-dependent degradation. FEBS J. 2012, 279, 532–542. [Google Scholar] [CrossRef]
- Eyal, G.; Eyal-Shaham, L.; Cohen, I.; Tamir, R.; Ben-Zvi, O.; Sinniger, F.; Loya, Y. Euphyllia paradivisa, a successful mesophotic coral in the northern Gulf of Eilat/Aqaba, Red Sea. Coral Reefs 2016, 35, 91–102. [Google Scholar] [CrossRef]
- Oakley, C.A.; Ameismeier, M.F.; Peng, L.; Weis, V.M.; Grossman, A.R.; Davy, S.K. Symbiosis induces widespread changes in the proteome of the model cnidarian Aiptasia. Cell. Microbiol. 2016, 18, 1009–1023. [Google Scholar] [CrossRef]
- Lehnert, E.M.; Mouchka, M.E.; Burriesci, M.S.; Gallo, N.D.; Schwarz, J.A.; Pringle, J.R. Extensive differences in gene expression between symbiotic and aposymbiotic cnidarians. G3 (Bethesda) 2014, 4, 277–295. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Jokiel, P.; Maragos, J.; Franzisket, L. Coral growth: Buoyant weight technique. In Coral Reef Research Methods; Stoddart, D.R., Johannes, R.E., Eds.; United Nations Educational, Scientific and Cultural Organization: Paris, France, 1978; pp. 529–542. [Google Scholar]
- Levy, O.; Appelbaum, L.; Leggat, W.; Gothlif, Y.; Hayward, D.C.; Miller, D.J.; Hoegh-Guldberg, O. Light-Responsive Cryptochromes from a Simple Multicellular Animal, the Coral Acropora millepora. Science 2007, 318, 467–470. [Google Scholar] [CrossRef]
- Levy, O.; Kaniewska, P.; Alon, S.; Eisenberg, E.; Karako-Lampert, S.; Bay, L.K.; Reef, R.; Rodriguez-Lanetty, M.; Miller, D.J.; Hoegh-Guldberg, O. Complex diel cycles of gene expression in coral-algal symbiosis. Science 2011, 331, 175. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2013, 29, 644–652. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Wessa, P. Pearson Correlation (v1.0.10) in Free Statistics Software (v1.1.23-r7), Office for Research Development and Education. 2015. Available online: http://www.wessa.net/rwasp_correlation.wasp/ ( accessed on 25 June 2019).
- Meyer, E.; Weis, V.M. Study of cnidarian-algal symbiosis in the “omics” age. Biol. Bull. 2012, 223, 44–65. [Google Scholar] [CrossRef]
- Wolfowicz, I.; Baumgarten, S.; Voss, P.A.; Hambleton, E.A.; Voolstra, C.R.; Hatta, M.; Guse, A. Aiptasia sp. larvae as a model to reveal mechanisms of symbiont selection in cnidarians. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Mohamed, A.R.; Cumbo, V.; Harii, S.; Shinzato, C.; Chan, C.X.; Ragan, M.A.; Bourne, D.G.; Willis, B.L.; Ball, E.E.; Satoh, N.; et al. The transcriptomic response of the coral Acropora digitifera to a competent Symbiodinium strain: The symbiosome as an arrested early phagosome. Mol. Ecol. 2016, 25, 3127–3141. [Google Scholar] [CrossRef]
- Seneca, F.O.; Palumbi, S.R. The role of transcriptome resilience in resistance of corals to bleaching. Mol. Ecol. 2015, 24, 1467–1484. [Google Scholar] [CrossRef]
- Eyal-Shaham, L.; Eyal, G.; Tamir, R.; Loya, Y. Reproduction, abundance and survivorship of two Alveopora spp. in the mesophotic reefs of Eilat, Red Sea. Sci. Rep. 2016, 6, 20964. [Google Scholar] [CrossRef]
- Downs, C.A.; Fauth, J.E.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Radic. Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- Downs, C.A.; Kramarsky-winter, E.; Martinez, J.; Kushmaro, A.; Woodley, C.M.; Loya, Y. Symbiophagy as a cellular mechanism for coral bleaching. Autopjagy 2009, 5, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Maor-Landaw, K.; Levy, O. Gene expression profiles during short-term heat stress; branching vs. massive Scleractinian corals of the Red Sea. PeerJ. 2016, 4. eCollection 2016. [Google Scholar] [CrossRef]
- Grottoli, A.G.; Rodrigues, L.J.; Palardy, J.E. Heterotrophic plasticity and resilience in bleached corals. Nature 2006, 440, 1186–1189. [Google Scholar] [CrossRef]
- Caroselli, E.; Mattioli, G.; Levy, O.; Falini, G.; Dubinsky, Z.; Goffredo, S. Inferred calcification rate of a Mediterranean azooxanthellate coral is uncoupled with sea surface temperature along an 8° latitudinal gradient. Front. Zool. 2012, 9, 32. [Google Scholar] [CrossRef]
- Caroselli, E.; Nanni, V.; Levy, O.; Falini, G.; Dubinsky, Z.; Goffredo, S. Latitudinal variations in biometry and population density of a mediterranean solitary coral. Limnol. Oceanogr. 2015, 60, 1356–1370. [Google Scholar] [CrossRef]
- Levy, O.; Karako-Lampert, S.; Waldman Ben-Asher, H.; Zoccola, D.; Pagès, G.; Ferrier-Pagès, C. Molecular assessment of the effect of light and heterotrophy in the scleractinian coral Stylophora pistillata. Proc. Biol. Sci. 2016, 283, 20153025. [Google Scholar] [CrossRef]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell Biology of Cnidarian-Dinoflagellate Symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 1–34. [Google Scholar] [CrossRef]
- Hagedorn, M.; Carter, V.L.; Lager, C.; Camperio Ciani, J.F.; Dygert, A.N.; Schleiger, R.D.; Henley, E.M. Potential bleaching effects on coral reproduction. Reprod. Fertil. Dev. 2016, 28, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.; Harrison, P.; Hoegh-guldberg, O. Coral Bleaching Reduces Reproduction of Scleractinian Corals and Increases Susceptibility to Future Stress. In Proceedings of the 9th International Coral Reef Symposium, Bali, Indonesia, 23–27 October 2000. [Google Scholar]
- Szmant, A.M.; Gassman, N.J. The effects of prolonged “bleaching” on the tissue biomass and reproduction of the reef coral Montastrea annularis. Coral Reefs 1990, 8, 217–224. [Google Scholar] [CrossRef]
- Bongaerts, P.; Ridgway, T.; Sampayo, E.M.; Hoegh-Guldberg, O. Assessing the “deep reef refugia” hypothesis: Focus on Caribbean reefs. Coral Reefs 2010, 29, 1–19. [Google Scholar] [CrossRef]
- Glynn, P.W. Coral reef bleaching: Facts, hypotheses and implications. Glob. Chang. Biol. 1996, 2, 495–509. [Google Scholar] [CrossRef]
- Inoue, M.; Shinmen, K.; Kawahata, H.; Nakamura, T.; Tanaka, Y.; Kato, A.; Shinzato, C.; Iguchi, A.; Kan, H.; Suzuki, A.; et al. Estimate of calcification responses to thermal and freshening stresses based on culture experiments with symbiotic and aposymbiotic primary polyps of a coral, Acropora digitifera. Glob. Planet. Chang. 2012, 92–93, 1–7. [Google Scholar] [CrossRef]
- Ohki, S.; Irie, T.; Inoue, M.; Shinmen, K.; Kawahata, H.; Nakamura, T.; Kato, A.; Nojiri, Y.; Suzuki, A.; Sakai, K.; et al. Calcification responses of symbiotic and aposymbiotic corals to near-future levels of ocean acidification. Biogeosciences 2013, 10, 6807–6814. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meron, D.; Maor-Landaw, K.; Weizman, E.; Waldman Ben-Asher, H.; Eyal, G.; Banin, E.; Loya, Y.; Levy, O. The Algal Symbiont Modifies the Transcriptome of the Scleractinian Coral Euphyllia paradivisa during Heat Stress. Microorganisms 2019, 7, 256. https://doi.org/10.3390/microorganisms7080256
Meron D, Maor-Landaw K, Weizman E, Waldman Ben-Asher H, Eyal G, Banin E, Loya Y, Levy O. The Algal Symbiont Modifies the Transcriptome of the Scleractinian Coral Euphyllia paradivisa during Heat Stress. Microorganisms. 2019; 7(8):256. https://doi.org/10.3390/microorganisms7080256
Chicago/Turabian StyleMeron, Dalit, Keren Maor-Landaw, Eviatar Weizman, Hiba Waldman Ben-Asher, Gal Eyal, Ehud Banin, Yossi Loya, and Oren Levy. 2019. "The Algal Symbiont Modifies the Transcriptome of the Scleractinian Coral Euphyllia paradivisa during Heat Stress" Microorganisms 7, no. 8: 256. https://doi.org/10.3390/microorganisms7080256