Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Cheese Samples and LAB Isolation
2.2. LAB Identification
2.3. Reference Strains and Culture Conditions
2.4. Safety Assessment of LAB Isolates
2.4.1. DNAse, Gelatinase, Hemolytic and Mucin Degradation Activities
2.4.2. Biogenic Amine Production
2.4.3. Antibiotic Susceptibility and MIC Determination
2.5. Lysozyme, Acidic and Bile Salts Tolerances
2.6. Survival during Simulated Gastrointestinal Transit
2.7. Antimicrobial Activity
2.8. Hydrophobicity, Auto-Aggregation, and Co-Aggregation Abilities
2.9. Adhesion to Caco-2 Cells
2.10. Antioxidant Activity
2.11. Anti-Inflammatory Activity
2.12. RNA Extraction and qRT-PCR
2.13. Preliminary Identification of Metabolites Responsible for Antagonistic Activity against Pathogens
2.14. Detection of Genes for Virulence and Antibiotic Resistance
2.15. Statistical Analysis
3. Results
3.1. LAB Isolation and Identification
3.2. Safety Properties
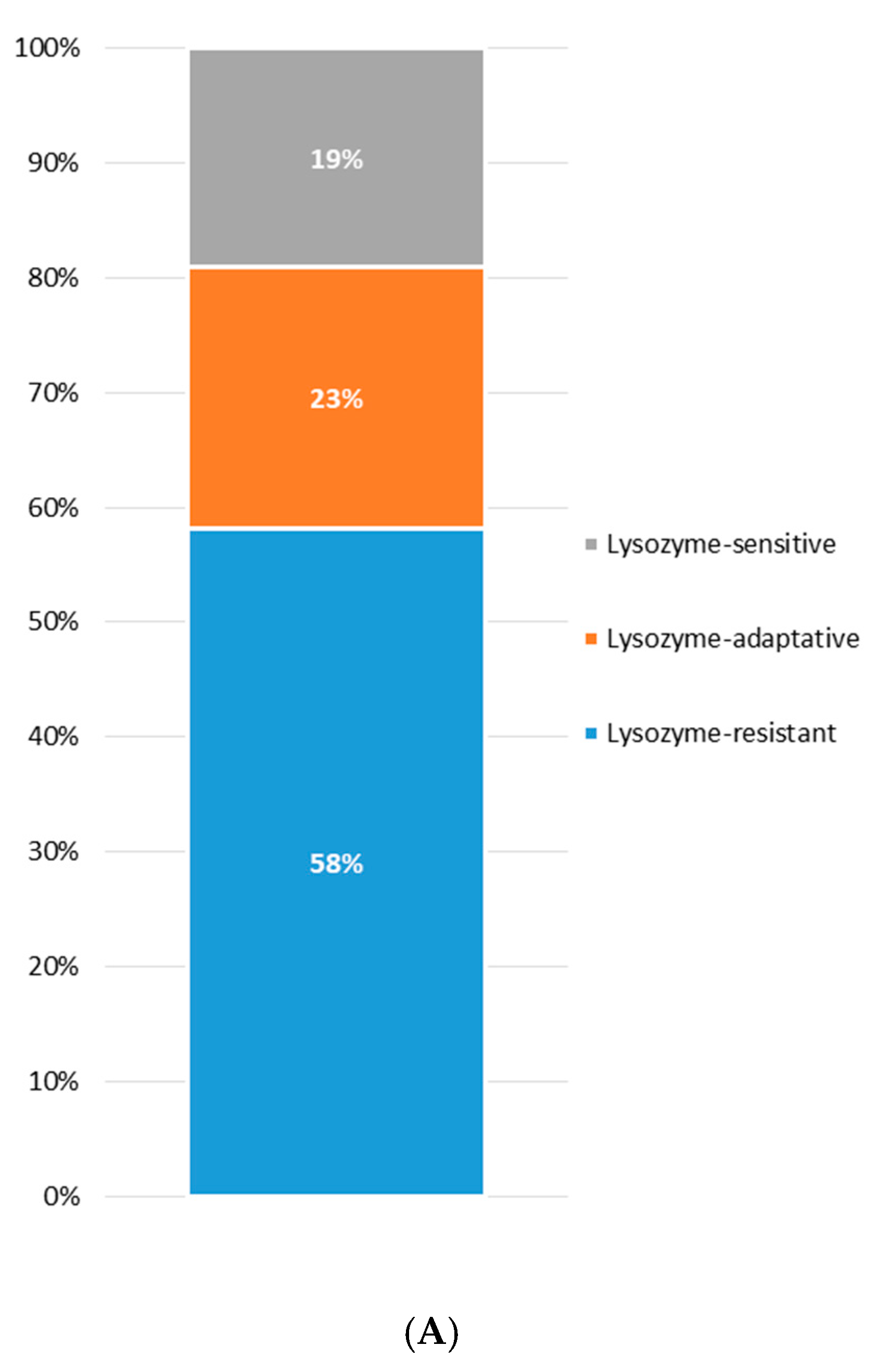
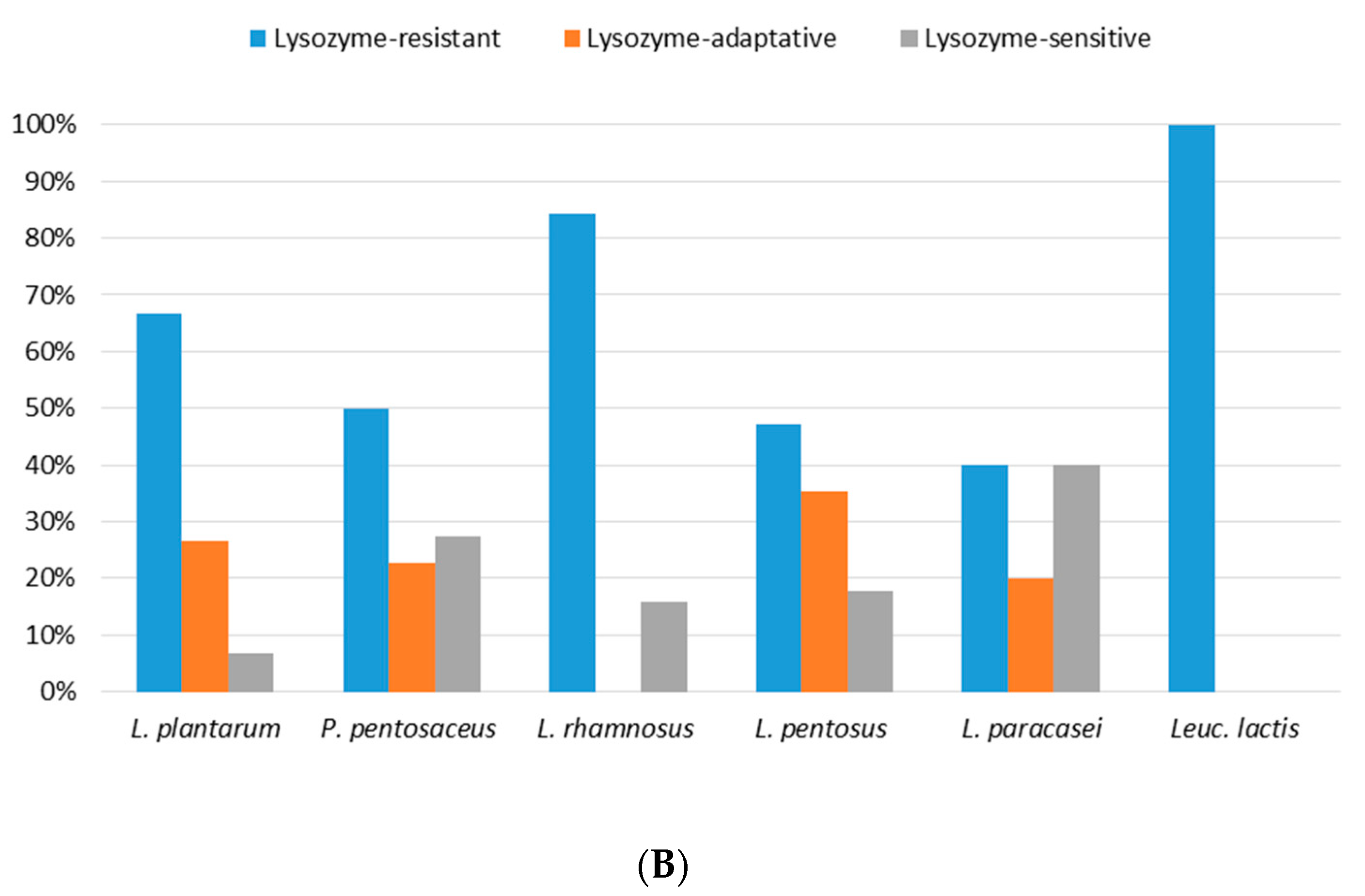
3.3. Lysozyme Tolerance
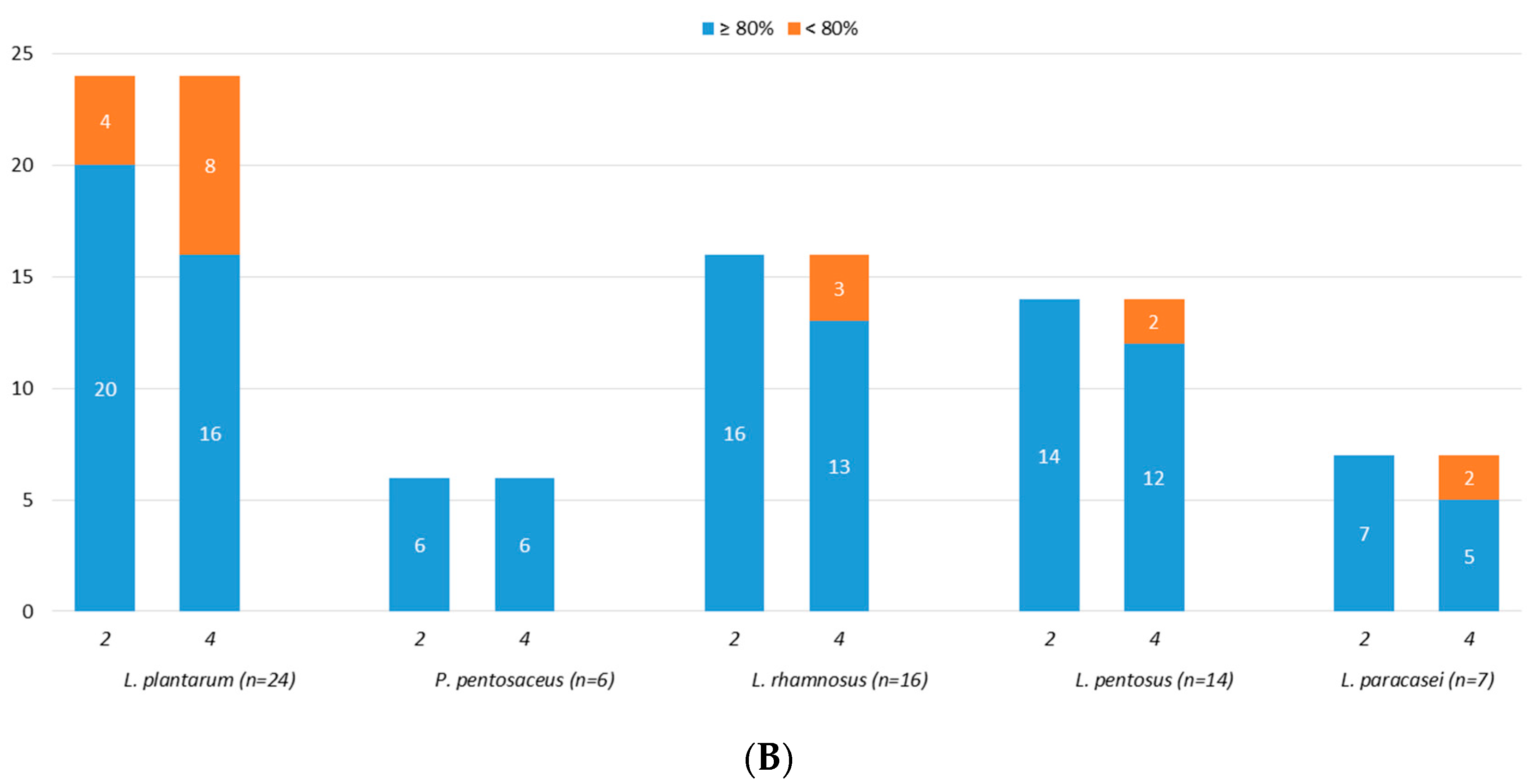
3.4. Acidic Tolerance
3.5. Bile Salts Tolerance
3.6. Survival During In Vitro GI Transit
3.7. Antagonistic Activity against Pathogens
3.8. Hydrophobicity, Auto-Aggregation, and Co-Aggregation Abilities
3.9. Adhesion to Caco-2 Cells
3.10. In Vitro Antioxidant Activity
3.11. In Vitro Anti-Inflammatory Activity
3.12. Preliminary Identification of Metabolites Responsible for Antagonistic Activity against Pathogens
3.13. Virulence Factors and Antibiotic Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaur, N.; Singh, D.P. Deciphering the consumer behaviour facets of functional foods: A literature review. Appetite 2017, 112, 167–187. [Google Scholar] [CrossRef] [PubMed]
- Abuajah, C.I.; Ogbonna, A.C.; Osuji, C.M. Functional components and medicinal properties of food: A review. J. Food Sci. Tech. 2015, 52, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrinets 2015, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Bella, F.; Godos, J.; Sciacca, S.; Del Rio, D.; Ray, S.; Galvano, F.; Giovannucci, E.L. Possible role of diet in cancer: Systematic review and multiple meta-analyses of dietary patterns, lifestyle factors, and cancer risk. Nutr. Rev. 2015, 75, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean Diet, its Components, and Cardiovascular Disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.J. Functional Foods and Active molecules with relevance to Health and Chronic disease: Editorial. Funct. Food Health Dis. 2017, 10, 849–852. [Google Scholar] [CrossRef]
- Martirosyan, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Foods Health Dis. 2015, 5, 209–223. [Google Scholar]
- Granato, D.K.; Branco, G.F.; Cruz, A.G.; de Faria, J.A.F.; Shah, N.P. Probiotic dairy products as functional foods. Compr. Rev. Food Sci. F. 2010, 9, 455–470. [Google Scholar] [CrossRef]
- Lenssen, K.G.M.; Bast, A.; Boer, A. Clarifying the health claim assessment procedure of EFSA will benefit functional food innovation. J. Funct. Foods. 2018, 47, 386–396. [Google Scholar] [CrossRef]
- FAO/WHO. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO/WHO: London, ON, Canada, 2002. [Google Scholar]
- Binns, N. Probiotics, prebiotics and the gut microbiota. ILSI Eur. Concise Monogr. 2013, 1, 1–32. [Google Scholar]
- Reid, G.; Jass, J.; Sebulsky, M.T.; McCormick, J.K. Potential uses of probiotics in clinical practice. Clin. Microbiol. Rev. 2003, 16, 658–672. [Google Scholar] [CrossRef] [PubMed]
- Ezendam, J.; van Loveren, H. Probiotics: Immunomodulation and evaluation of safety and efficacy. Nutr. Rev. 2006, 64, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Santosa, S.; Farnworth, E.; Jones, P.J.H. Probiotics and their potential health claims. Nutr. Rev. 2006, 64, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Capellini, B.; Villarreal, F.; Suárez, V.; Quiberoni, A.; Reinheimer, J. Usefulness of a set of simple in vitro tests for the screening and identification of probiotic candidate strains for dairy use. LWT Food Sci. Technol. 2008, 41, 1678–1688. [Google Scholar] [CrossRef]
- Aureli, P.; Capurso, L.; Castellazzi, A.M.; Clerici, M.; Giovannini, M.; Morelli, L.; Poli, A.; Pregliasco, F.; Salvini, F.; Zuccotti, G.V. Probiotics and health: An evidence-based review. Pharmacol. Res. 2011, 63, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.; Grimmer, S.; Naterstad, K.; Axelsson, L. In vitro testing of commercial and potential probiotic lactic acid bacteria. Int. J. Food Microbiol. 2012, 153, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef]
- Zielińska, D.; Długosz, E.; Zawistowska-Deniziak, E. Functional Properties of Food Origin Lactobacillus in the Gastrointestinal Ecosystem—In Vitro Study. Probiotics Antimicrob. Proteins 2018. [Google Scholar] [CrossRef]
- Puniya, M.; Kumar, R.; Panwar, H.; Kumar, N.; Ramneek, A.K.P. Screening of Lactic Acid Bacteria of Different Origin for their Probiotic Potential. J. Food Process. Technol. 2016, 7, 1. [Google Scholar]
- Castro, J.M.; Tornadijo, M.E.; Fresno, J.M.; Sandoval, H. Biocheese: A Food Probiotic Carrier. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Drosinos, E.H. Probiotic Dairy Products: Inventions Toward Ultramodern Production. In Innovations in Technologies for Fermented Food and Beverage Industries; Panda, S., Shetty, P., Eds.; Food Microbiology and Food Safety, Springer: Cham, Switzerland, 2018; pp. 143–157. [Google Scholar]
- Plessas, S.; Bosnea, L.; Alexopoulos, A.; Bezirtzoglou, E. Potential effects of probiotics in cheese and yogurt production: A review. Eng. Life Sci. 2012, 12, 433–440. [Google Scholar] [CrossRef]
- Caggia, C.; De Angelis, M.; Pitino, I.; Pino, A.; Randazzo, C.L. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses. Food Microb. 2015, 50, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microb. 2017, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, P.; Kelly, A.L.; Sheehan, J.J. Symposium review: Structure-function relationships in cheese. J. Dairy Sci. 2018, 101, 2692–2709. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.B.A.; Deliza, R.; dos Santos, K.M.O.; Walter, E.H.M.; Martins, J.M.; Rosenthal, A. Viability of Probiotics in Goat Cheese During Storage and Under Simulated Gastrointestinal Conditions. Food Bioprocess Technol. 2018, 11, 853–863. [Google Scholar] [CrossRef]
- Horne, J.; Carpino, S.; Tuminello, L.; Rapisarda, T.; Corallo, L.; Licitra, G. Volatiles, chemical, microbial and sensory characterization of artisanal and industrial Piacentinu Ennese cheese. Int. Dairy J. 2005, 15, 605–617. [Google Scholar] [CrossRef]
- Fallico, V.; Tuminello, L.; Pediliggieri, C.; Horne, J.; Carpino, S.; Licitra, G. Proteolysis and microstructure of Piacentinu Ennese cheese made using different farm technologies. J. Dairy Sci. 2006, 89, 37–48. [Google Scholar] [CrossRef]
- Pino, A.; Liotta, L.; Randazzo, C.L.; Todaro, C.; Mazzaglia, A.; De Nardo, F.; Chiofalo, V.; Caggia, C. Polyphasic approach to study physico-chemical, microbiological and sensorial characteristics of artisanal Nicastrese goat’s cheese. Food Microb. 2018, 70, 143–154. [Google Scholar] [CrossRef]
- Pino, A.; Van Hoorde, K.; Pitino, I.; Russo, N.; Carpino, S.; Caggia, C.; Randazzo, C.L. Survival of potential probiotic lactobacilli used as adjunct cultures on Pecorino Siciliano cheese ripening and passage through the gastrointestinal tract of healthy volunteers. Int. J. Food Microbiol. 2017, 252, 42–52. [Google Scholar] [CrossRef]
- Russo, N.; Caggia, N.; Pino, A.; Coque, T.M.; Arioli, S.; Randazzo, C.L. Enterococcus spp. in Ragusano PDO and Pecorino Siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef]
- Lavilla-Lerma, L.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Characterization of functional, safety, and gut survival related characteristics of Lactobacillus strains isolated from farmhouse goat’s milk cheeses. Int. J. Food Microbiol. 2013, 163, 136–145. [Google Scholar] [CrossRef]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; del Campo, R.; Hernández, P.E. Antimicrobial activity, antibiotic susceptibility and virulence factors of lactic acid bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- ISO: International Organization for Standardization. Milk and Milk Products—Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB) ISO 10932/IDF 233 Standard 2010; ISO: International Organization for Standardization: Geneva, Switzerland, 2010. [Google Scholar]
- Pithva, S.; Shekh, S.; Dave, J.; Vyas, B.R. Probiotic attributes of autochthonous Lactobacillus rhamnosus strains of human origin. Appl. Biochem. Biotechnol. 2014, 173, 259–277. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Russo, N.; Pino, A.; Mazzaglia, A.; Ferrante, M.; Oliveri Conti, G.; Caggia, C. Effects of selected bacterial cultures on safety and sensory traits of Nocellara Etnea olives produced at large factory scale. Food Chem. Toxicol. 2018, 115, 491–498. [Google Scholar] [CrossRef]
- Solieri, L.; Bianchi, A.; Mottolese, G.; Lemmetti, F.; Giudici, P. Tailoring the probiotic potential of non-starter Lactobacillus strains from ripened Parmigiano Reggiano cheese by in vitro screening and principal component analysis. Food Microb. 2014, 38, 240–249. [Google Scholar] [CrossRef]
- Dhanani, A.S.; Gaudana, S.B.; Bagchi, T. The ability of Lactobacillus adhesin EF-Tu to interfere with pathogen adhesion. Eur. Food Res. Technol. 2011, 232, 777–785. [Google Scholar] [CrossRef]
- Ferrara, G.; Giancaspro, A.; Mazzeo, A.; Giove, S.L.; Matarrese, A.M.S.; Pacucci, C.; Punzi, R.; Trani, A.; Gambacorta, G.; Blanco, A.; et al. Characterization of pomegranate (Punica granatum L.) genotypes collected in Puglia region, Southeastern Italy. Sci. Hortic. 2014, 178, 70–78. [Google Scholar] [CrossRef]
- Osawa, T.; Namiki, M. Natural antioxidant isolated from Eucalyptus leaf waxes. J. Agric. Food Chem. 1985, 33, 777–780. [Google Scholar] [CrossRef]
- Campanella, D.; Rizzello, C.G.; Fasciano, C.; Gambacorta, G.; Pinto, D.; Marzani, B.; Scarano, N.; De Angelis, M.; Gobbetti, M. Exploitation of grape marc as functional substrate for lactic acid bacteria and bifidobacteria growth and enhanced antioxidant activity. Food Microb. 2017, 65, 25–35. [Google Scholar] [CrossRef]
- Mitsuta, H.; Yasumoto, K.; Iwami, K. Antioxidative action of indole compounds during the autoxidation of linoleic acid. Nihon Koshu Eisei Zasshi 1996, 19, 210–214. [Google Scholar]
- Xu, T.; Pan, L.; Ge, Y.; Li, P.; Meng, X.; Huang, C.; Li, J. Tmem88 Mediates Inflammatory Cytokines Secretion by Regulating Jnk/P38 and Canonical Wnt/Beta-Catenin Signaling Pathway in Lx-2 Cells. Inflammopharmacology 2018, 26, 1339–1348. [Google Scholar] [CrossRef]
- Zheng, K.; Chen, Z.; Sun, W.; Liu, B.; Fan, D.; Guo, O.; Luo, H.; Shen, J.; Li, L.; He, X.; et al. Hei-Gu-Teng Zhuifenghuoluo Granule Modulates IL-12 Signal Pathway to Inhibit the Inflammatory Response in Rheumatoid Arthritis. J. Immunol. Res. 2018. [Google Scholar] [CrossRef]
- dos Santos, K.M.O.; Vieira, A.D.S.; Buriti, F.C.A.; do Nascimento, J.C.F.; de Melo, M.E.S.; Bruno, L.M.; de Fátima Borges, M.; Rocha, C.R.C.; de Souza Lopes, A.C.; de Melo Franco, B.D.G.; et al. Artisanal Coalho cheeses as source of beneficial Lactobacillus plantarum and Lactobacillus rhamnosus strains. Dairy Sci. Technol. 2015, 95, 209–230. [Google Scholar] [CrossRef]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and Its Bacteriocin. Probiotics Antimicrob. Proteins 2017, 9, 334–344. [Google Scholar] [CrossRef]
- Zoumpopoulou, G.; Tzouvanou, A.; Mavrogonatou, E.; Alexandraki, V.; Georgalaki, M.; Anastasiou, R.; Papadelli, M.; Manolopoulou, E.; Kazou, M.; Kletsas, D.; et al. Probiotic features of lactic acid bacteria isolated from a diverse pool of traditional Greek dairy products regarding specific strain-host interactions. Probiotics Antimicrob. Proteins 2018, 10, 313–322. [Google Scholar] [CrossRef]
- Kerrya, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented foods, the gut and mental health: A mechanistic overview with implications for depression and anxiety. Nutr. Neurosci. 2018. [Google Scholar] [CrossRef]
- Gille, D.; Schmid, A.; Walther, B.; Vergères, G. Fermented Food and Non-Communicable Chronic Diseases: A Review. Nutrients 2018, 10, 448. [Google Scholar] [CrossRef]
- Mota de Carvalho, N.; Costa, E.M.; Silva, S.; Pimentel, L.; Fernandes, T.H.; Pintado, M.E. Fermented Foods and Beverages in Human Diet and Their Influence on Gut Microbiota and Health. Fermentation 2018, 4, 90. [Google Scholar] [CrossRef]
- Carpino, S.; Randazzo, C.L.; Pino, A.; Russo, N.; Rapisarda, T.; Belvedere, G.; Caggia, C. Influence of PDO Ragusano cheese biofilm microbiota on flavour compounds formation. Food Microb. 2017, 61, 126–135. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Torriani, S.; Akkermans, A.D.L.; de Vos, W.M.; Vaughan, E.E. Diversity, Dynamics, and Activity of Bacterial Communities during Production of an Artisanal Sicilian Cheese as Evaluated by 16S rRNA Analysis. Appl. Environ. Microbiol. 2002, 68, 1882–1892. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Vaughan, E.E.; Caggia, C. Artisanal and experimental Pecorino Siciliano cheese: Microbial dynamics during manufacture assessed by culturing and PCR-DGGE analyses. Int. J. Food Microbiol. 2006, 109, 1–8. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Pitino, I.; Ribbera, A.; Caggia, C. Pecorino Crotonese cheese: Study of bacterial population and flavour compounds. Food Microb. 2010, 27, 363–374. [Google Scholar] [CrossRef]
- Guarcello, R.; Carpino, S.; Gaglio, R.; Pino, A.; Rapisarda, T.; Caggia, C.; Marino, G.; Randazzo, C.L.; Settanni, L.; Todaro, M. large factory-scale application of selected autochthonous lactic acid bacteria for PDO Pecorino Siciliano cheese production. Food Microb. 2016, 59, 66–75. [Google Scholar] [CrossRef]
- Manolopoulou, E.; Sarantinopoulos, P.; Zoidou, E.; Aktypis, A.; Moschopoulou, E.; Kandarakis, I.G.; Anifantakis, E.M. Evolution of microbial populations during traditional Feta cheese manufacture and ripening. Int. J. Food Microbiol. 2003, 82, 153–161. [Google Scholar] [CrossRef]
- Nieto-Arribas, P.; Sesena, S.; Poveda, J.M.; Palop, L.; Cabezas, L. Genotypic and technological characterization of Leuconostoc isolates to be used as adjunct starters in Manchego cheese manufacture. Food Microbiol. 2010, 27, 85–93. [Google Scholar] [CrossRef]
- Terzic-Vidojevic, A.; Tolinacki, M.; Nikolic, M.; Lozo, J.; Begovic, J.; Gulahmadov, S.G.O.; Kuliev, A.A.; Dalgalarrondo, M.; Chobert, J.-M.; Haertle, T.; et al. Phenotypic and genotypic characterization of lactic acid bacteria isolated from Azerbaijani traditional dairy products. Afr. J. Biotechnol. 2009, 8, 2576–2588. [Google Scholar]
- Reid, G. The importance of guidelines in the development and application of probiotics. Curr. Pharm. Des. 2005, 11, 11–16. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Devirgiliis, C.; Barile, S.; Perozzi, G. Antibiotic resistance determinants in the interplay between food and gut microbiota. Genes Nutr. 2011, 6, 275–284. [Google Scholar] [CrossRef]
- Ahn, C.; Collins-Thompson, D.; Duncan, C.; Stiles, M.E. Mobilization and location of the genetic determinant of chloramphenicol resistance from Lactobacillus plantarum caTC2R. Plasmid 1992, 27, 169–176. [Google Scholar] [CrossRef]
- Lin, C.F.; Fung, Z.F.; Wu, C.L.; Chung, T.C. Molecular characterization of a plasmid-borne (pTC82) chloramphenicol resistance determinant (cat-TC) from Lactobacillus reuteri G4. Plasmid 1996, 36, 116–124. [Google Scholar] [CrossRef]
- Klein, G.; Hallmann, C.; Casas, I.A.; Abad, J.; Louwers, J.; Reuter, G. Exclusion of vanA, vanB and vanC type glycopeptide resistance in strains of Lactobacillus reuteri and Lactobacillus rhamnosus used as probiotics by polymerase chain reaction and hybridization methods. J. Appl. Microbiol. 2000, 89, 815–824. [Google Scholar] [CrossRef]
- Danielsen, M.; Wind, A. Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 2003, 82, 1–11. [Google Scholar] [CrossRef]
- Cataloluk, O.; Gogebakan, B. Presence of drug resistance in intestinal lactobacilli of dairy and human origin in Turkey. FEMS Microbiol. Lett. 2004, 236, 7–12. [Google Scholar] [CrossRef]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef]
- Huys, G.; D’Haene, K.; Danielsen, M.; Mättö, J.; Egervärn, M.; Vandamme, P. Phenotypic and molecular assessment of antimicrobial resistance in Lactobacillus paracasei strains of food origin. J. Food Prot. 2008, 71, 339–344. [Google Scholar] [CrossRef]
- Korhonen, J.M.; Van Hoek, A.H.A.M.; Saarela, M.; Huys, G.; Tosi, L.; Mayrhofer, S.; Von Wright, A. Antimicrobial susceptibility of Lactobacillus rhamnosus. Benef. Microbes 2010, 1, 75–80. [Google Scholar] [CrossRef]
- Comunian, R.; Daga, E.; Dupré, I.; Paba, A.; Devirgiliis, C.; Piccioni, V.; Perozzi, G.; Zonenschain, D.; Rebecchi, A.; Morelli, L.; et al. Susceptibility to tetracycline and erythromycin of Lactobacillus paracasei strains isolated from traditional Italian fermented foods. Int. J. Food Microbiol. 2010, 31, 151–156. [Google Scholar] [CrossRef]
- Hummel, A.S.; Hertel, C.; Holzapfel, W.H.; Franz, C.M. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Appl. Environ. Microbiol. 2007, 73, 730–739. [Google Scholar] [CrossRef]
- Adimpong, D.B.; Nielsen, D.S.; Sørensen, K.I.; Derkx, P.M.; Jespersen, L. Genotypic characterization and safety assessment of lactic acid bacteria from indigenous African fermented food products. BMC Microbiol. 2012, 12, 75. [Google Scholar] [CrossRef]
- Alegría, Á.; Delgado, S.; Flórez, A.B.; Mayo, B. Identification, typing, and functional characterization of Leuconostoc spp. strains from traditional, starter-free cheeses. Dairy Sci. Technol. 2013, 93, 657–673. [Google Scholar] [CrossRef]
- Flórez, A.B.; Campedelli, I.; Delgado, S.; Alegría, Á.; Salvetti, E.; Felis, G.E.; Mayo, B.; Torriani, S. Antibiotic Susceptibility Profiles of Dairy Leuconostoc, Analysis of the Genetic Basis of Atypical Resistances and Transfer of Genes In Vitro and in a Food Matrix. PLoS ONE 2016, 11, e0145203. [Google Scholar] [CrossRef]
- Picón, A.; Garde, S.; Ávila, M.; Núñez, M. Microbiota dynamics and lactic acid bacteria biodiversity in raw goat milk cheeses. Int. Dairy J. 2016, 58, 14–22. [Google Scholar] [CrossRef]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef]
- Bibiloni, R.; Pérez, P.F.; Garrote, L.G.; Disalvo, E.A.; De Antoni, G.L. Surface characterization and adhesive properties of Bifidobacteria. Methods Enzym. 2001, 336, 411–427. [Google Scholar]
- Kos, B.; Suskovic, J.; Vukovic, S.; Simpraga, M.; Frece, J.; Matosic, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Shahidi, F.; Mortazavi, S.A.; Milani, E.; Eshaghi, Z. Potentially Probiotic Lactobacillus Strains from Traditional Kurdish Cheese. Probiotics Antimicrob. Proteins 2014, 6, 22–31. [Google Scholar] [CrossRef]
- Oelschlaeger, T.A. Mechanisms of probiotic actions—A review. Int. J. Med. Microbiol. 2010, 300, 57–62. [Google Scholar] [CrossRef]
- Pino, A.; Bartolo, E.; Caggia, C.; Cianci, A.; Randazzo, C.L. Detection of vaginal lactobacilli as probiotic candidates. Sci. Rep. 2019, 9, 3355. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Ayyanna, R.; Ankaiah, D.; Arul, V. Anti-inflammatory and Antioxidant Properties of Probiotic Bacterium Lactobacillus mucosae AN1 and Lactobacillus fermentum SNR1 in Wistar Albino Rats. Front Microbiol. 2018, 9, 3063. [Google Scholar] [CrossRef]
- Tuberoso, C.I.G.; Rosa, A.; Bifulco, E.; Melis, M.P.; Atzeri, A.; Pirisi, F.M.; Dessi, M.A. Chemical composition and antioxidant activities of Myrtus communis L. berries extracts. Food Chem. 2010, 123, 1242–1251. [Google Scholar] [CrossRef]
- Williams, C.S.; Mann, M.; DuBois, R.N. The Role of Cyclooxygenases in Inflammation, Cancer, and Development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef]
- Grosch, S.; Niederberger, E.; Geisslinger, G. Investigational Drugs Targeting the Prostaglandin E2 Signaling Pathway for the Treatment of Inflammatory Pain. Expert Opin. Investig. Drugs 2017, 26, 51–61. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, C.; Chen, H.; Wang, C.; Liu, X.; Qiu, M.; Tang, H.; Zhang, F.; Zhou, X.; Yang, J. Inhibition of Cox-2/Mpges-1 and 5-Lox in Macrophages by Leonurine Ameliorates Monosodium Urate Crystal-Induced Inflammation. Toxicol. Appl. Pharmacol. 2018, 351, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | CI | SGJ | SIF |
|---|---|---|---|---|
| L. plantarum (n = 16) | PE09 | 9.03 ± 0.11 | 5.30 ± 0.02 | 5.11 ± 0.02 |
| PE11 | 9.12 ± 0.03 | 8.96 ± 0.06 | 8.84 ± 0.02 | |
| PE27 | 9.10 ± 0.07 | 8.02 ± 0.03 | 8.06 ± 0.06 | |
| PE30 | 9.01 ± 0.14 | 8.11 ± 0.02 | 8.01 ± 0.02 | |
| PE41 | 9.16 ± 0.02 | 4.98 ± 0.05 | 4.16 ± 0.03 | |
| PE42 | 9.28 ± 0.02 | 5.26 ± 0.08 | 5.15 ± 0.06 | |
| PE58 | 9.21 ± 0.05 | 5.02 ± 0.02 | 4.00 ± 0.03 | |
| PE66 | 9.22 ± 0.03 | 8.14 ± 0.06 | 8.03 ± 0.05 | |
| PE77 | 9.31 ± 0.06 | 5.31 ± 0.02 | 5.03 ± 0.08 | |
| PE92 | 9.24 ± 0.08 | 8.16 ± 0.02 | 8.11 ± 0.03 | |
| PE99 | 9.18 ± 0.05 | 9.01 ± 0.03 | 8.86 ± 0.06 | |
| PE101 | 9.20 ± 0.11 | 8.28 ± 0.04 | 8.14 ± 0.02 | |
| PE107 | 9.30 ± 0.13 | 4.05 ± 0.02 | 3.83 ± 0.02 | |
| PE115 | 9.12 ± 0.02 | 6.00 ± 0.06 | 5.84 ± 0.03 | |
| PE139 | 9.06 ± 0.12 | 4.21 ± 0.08 | 4.07 ± 0.10 | |
| PE141 | 9.19 ± 0.05 | 5.11 ± 0.02 | 5.03 ± 0.02 | |
| L. rhamnosus (n = 13) | PE07 | 9.13 ± 0.04 | 8.12 ± 0.07 | 8.06 ± 0.08 |
| PE08 | 9.11 ± 0.03 | 8.13 ± 0.04 | 8.02 ± 0.03 | |
| PE13 | 9.06 ± 0.07 | 8.01 ± 0.04 | 8.00 ± 0.02 | |
| PE25 | 9.14 ± 0.05 | 8.20 ± 0.06 | 8.15 ± 0.02 | |
| PE44 | 9.19 ± 0.07 | 8.01 ± 0.08 | 7.93 ± 0.06 | |
| PE61 | 9.10 ± 0.04 | 8.00 ± 0.05 | 7.96 ± 0.04 | |
| PE64 | 9.26 ± 0.06 | 8.87 ± 0.02 | 8.51 ± 0.04 | |
| PE81 | 9.23 ± 0.08 | 8.68 ± 0.02 | 8.05 ± 0.06 | |
| PE113 | 9.36 ± 0.04 | 4.88 ± 0.06 | 3.12 ± 0.04 | |
| PE137 | 9.07 ± 0.03 | 5.03 ± 0.06 | 4.91 ± 0.03 | |
| PE138 | 9.03 ± 0.11 | 5.21 ± 0.12 | 5.10 ± 0.07 | |
| PE151 | 9.23 ± 0.05 | 5.23 ± 0.11 | 5.06 ± 0.10 | |
| PE152 | 9.22 ± 0.07 | 9.04 ± 0.08 | 8.93 ± 0.06 | |
| L. pentosus (n = 12) | PE14 | 9.15 ± 0.04 | 8.11 ± 0.03 | 8.04 ± 0.03 |
| PE15 | 9.11 ± 0.03 | 8.02 ± 0.04 | 7.96 ± 0.06 | |
| PE16 | 9.23 ± 0.11 | 4.98 ± 0.07 | 4.33 ± 0.07 | |
| PE21 | 9.17 ± 0.06 | 9.03 ± 0.04 | 8.96 ± 0.07 | |
| PE38 | 9.11 ± 0.03 | 5.12 ± 0.08 | 4.91 ± 0.07 | |
| PE72 | 9.07 ± 0.08 | 8.03 ± 0.07 | 7.94 ± 0.08 | |
| PE73 | 9.09 ± 0.07 | 8.21 ± 0.06 | 8.07 ± 0.07 | |
| PE122 | 9.11 ± 0.06 | 8.18 ± 0.07 | 8.09 ± 0.04 | |
| PE123 | 9.16 ± 0.03 | 5.54 ± 0.08 | 5.31 ± 0.03 | |
| PE124 | 9.09 ± 0.06 | 8.86 ± 0.12 | 8.03 ± 0.06 | |
| PE159 | 9.21 ± 0.11 | 5.18 ± 0.14 | 5.03 ± 0.04 | |
| PE160 | 9.18 ± 0.04 | 8.07 ± 0.04 | 7.91 ± 0.07 | |
| P. pentosaceus (n = 6) | PE96 | 9.16 ± 0.08 | 8.25 ± 0.02 | 8.13 ± 0.02 |
| PE104 | 9.07 ± 0.02 | 8.15 ± 0.04 | 7.95 ± 0.03 | |
| PE120 | 9.30 ± 0.10 | 8.21 ± 0.03 | 8.02 ± 0.02 | |
| PE133 | 9.28 ± 0.02 | 5.24 ± 0.02 | 5.21 ± 0.04 | |
| PE135 | 9.18 ± 0.04 | 5.02 ± 0.02 | 4.56 ± 0.06 | |
| PE136 | 9.15 ± 0.06 | 4.34 ± 0.02 | 4.46 ± 0.02 | |
| L. paracasei (n = 5) | PE04 | 9.16 ± 0.06 | 9.07 ± 0.06 | 8.94 ± 0.06 |
| PE24 | 9.09 ± 0.07 | 8.24 ± 0.07 | 8.10 ± 0.03 | |
| PE85 | 9.13 ± 0.11 | 8.16 ± 0.14 | 7.98 ± 0.07 | |
| PE86 | 9.20 ± 0.07 | 8.83 ± 0.07 | 8.03 ± 0.14 | |
| PE102 | 9.14 ± 0.03 | 8.96 ± 0.07 | 8.84 ± 0.04 |
| Species | Strains | E. coli ATCC 25922 | S. aureus ATCC 6538 | L. monocytogenes DSM 12464 | S. enterica s. typhimurium ATCC 14028 |
|---|---|---|---|---|---|
| L. rhamnosus (n = 9) | PE44, PE61, PE25, PE13 | +++ | ++ | +++ | +++ |
| PE64, PE08, PE81 | ++ | - | +++ | ++ | |
| PE07 | - | ++ | - | + | |
| PE152 | ++ | ++ | - | - | |
| L. pentosus (n = 8) | PE14, PE15, PE21 | +++ | ++ | ++ | +++ |
| PE72, PE73 | +++ | +++ | - | + | |
| PE124, PE160, PE122 | - | - | - | - | |
| L. plantarum (n = 7) | PE30 | + | - | - | - |
| PE92, PE101, PE11 | - | - | - | - | |
| PE66 | - | + | - | + | |
| PE09, PE27 | + | + | - | - | |
| L. paracasei (n = 5) | PE24, PE85, PE86 | ++ | +++ | ++ | ++ |
| PE04, PE102 | - | - | - | ||
| P. pentosaceus (n = 3) | PE96, PE104 | - | - | - | - |
| PE120 | - | - | - | - |
| Species | Strains | H% | Auto-A% | CoA% | ||
|---|---|---|---|---|---|---|
| E. coli 555 | L. monocytogenes | S. enterica S. typhimurium | ||||
| L. plantarum | PE27 | 28.13 ± 0.54 a | 12.30 ± 0.19 a | 14.20 ± 0.20 c | 23.22 ± 0.21 e | 24.31 ± 0.26 e |
| PE30 | 31.09 ± 0.19 b | 53.13 ± 0.15 e | 21.47 ± 0.27 d | 32.54 ± 0.20 h | 12.25 ± 0.15 b | |
| PE92 | 41.21 ± 0.38 d | 56.30 ± 0.23 g | 46.18 ± 0.12 h | 14.38 ± 0.22 b | 37.17 ± 0.10 h | |
| PE101 | 36.30 ± 0.16 c | 58.41 ± 0.38 h | 31.48 ± 0.27 f | 23.20 ± 0.18 e | 28.63 ± 0.18 g | |
| L. rhamnosus | PE07 | 43.12 ± 0.30 e | 74.11 ± 0.14 pq | 59.23 ± 0.07 no | 58.29 ± 0.24 p | 67.33 ± 0.18 r |
| PE13 | 71.36 ± 0.42 m | 77.41 ± 0.23 s | 51.29 ± 0.16 i | 60.33 ± 0.18 rr | 52.28 ± 0.13 i | |
| PE25 | 78.14 ± 0.13 q | 71.37 ± 0.28 n | 68.27 ± 0.16 r | 72.35 ± 0.19 v | 58.26 ± 0.15 m | |
| PE64 | 56.39 ± 0.33 l | 65.20 ± 0.20 l | 54.26 ± 0.13 l | 67.34 ± 0.27 s | 52.42 ± 0.14 i | |
| PE08 | 46.24 ± 0.31 f | 69.03 ± 0.23 m | 51.17 ± 0.12 i | 52.37 ± 0.15 m | 60.22 ± 0.19 o | |
| PE81 | 41.14 ± 0.37 d | 59.99 ± 0.27 i | 58.59 ± 0.20 n | 53.34 ± 0.24 n | 60.30 ± 0.20 o | |
| PE152 | 48.35 ± 0.35 g | 72.29 ± 0.09 o | 63.24 ± 0.18 p | 59.37 ± 0.28 q | 56.37 ± 0.22 l | |
| PE44 | 72.23 ± 0.20 mn | 71.47 ± 0.35 no | 72.35 ± 0.23 t | 51.41 ± 0.17 l | 58.64 ± 0.15 mn | |
| PE61 | 74.29 ± 0.26 no | 76.04 ± 0.34 r | 57.27 ± 0.16 m | 42.93 ± 0.32 i | 59.27 ± 0.23 n | |
| L. pentosus | PE21 | 52.11 ± 0.10 i | 19.24 ± 0.17 c | 32.27 ± 0.16 g | 28.18 ± 0.12 f | 24.16 ± 0.12 e |
| PE72 | 56.33 ± 0.26 l | 15.16 ± 0.19 b | 14.20 ± 0.12 c | 29.34 ± 0.21 g | 21.73 ± 0.23 d | |
| PE73 | 48.22 ± 0.15 g | 23.38 ± 0.24 d | 6.35 ± 0.20 a | 12.18 ± 0.12 a | 18.71 ± 0.12 c | |
| PE124 | 46.35 ± 0.45 f | 54.61 ± 0.28 f | 23.40 ± 0.28 e | 18.21 ± 0.18 d | 26.74 ±0.22 f | |
| PE160 | 50.01 ± 0.53 h | 57.17 ± 0.13 g | 11.26 ± 0.16 b | 15.29 ± 0.24 c | 9.37 ± 0.20 a | |
| L. paracasei | PE24 | 76.37 ± 0.35 p | 76.37 ± 0.24 r | 71.48 ± 0.21 s | 71.29 ± 0.18 u | 61.33 ± 0.11 p |
| PE85 | 73.25 ± 0.24 no | 73.23 ± 0.19 p | 66.34 ± 0.22 q | 69.27 ± 0.17 t | 71.19 ± 0.13 s | |
| PE86 | 74.33 ± 0.34 no | 74.36 ± 0.22 q | 59.60 ± 0.20 o | 56.39 ± 0.26 o | 63.28 ± 0.13 q | |
| COX-1 | COX-2 | IL-8 | IL-10 | |
|---|---|---|---|---|
| LPS | 2.07 ± 0.10 | 15.22 ± 0.76 | 0.64 ± 0.03 | 128.50 ± 6.42 |
| PE13 | 34.31 ± 1.71 * | 3.24 ± 17.81 * | 1.42 ± 0.07 * | 103192 ± 5159 * |
| PE24 | 24.05 ± 1.20 * | 0.09 ± 0.004 * | 0.60 ± 0.03 | 615.96 ± 30.79 * |
| PE25 | 1.18 ± 0.21 | 2.28 ± 0.06 * | 0.003 ± 0.02 * | 9153 ± 12.1 * |
| PE44 | 0.42 ± 0.02 * | 2.41 ± 0.12 * | 0.0018 ± 9.27 × 10−5 * | 488 ± 0.24 * |
| PE61 | 0.002 ± 0.0001 * | 0.69 ± 0.03 * | 0.00053 ± 2.68 × 10−5 * | 797.55 ± 39.87 * |
| PE85 | 3.65 ± 0.18 * | 36.51 ± 1.82 * | 0.38 ± 0.019 | 3360.34 ± 168.01 * |
| PE86 | 2.28 ± 0.11 | 0.28 ± 0.01 * | 0.0018 ± 9.01 × 10−5 * | 1617.93 ± 80.89 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms 2019, 7, 254. https://doi.org/10.3390/microorganisms7080254
Pino A, Russo N, Van Hoorde K, De Angelis M, Sferrazzo G, Randazzo CL, Caggia C. Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms. 2019; 7(8):254. https://doi.org/10.3390/microorganisms7080254
Chicago/Turabian StylePino, Alessandra, Nunziatina Russo, Koenraad Van Hoorde, Maria De Angelis, Giuseppe Sferrazzo, Cinzia Lucia Randazzo, and Cinzia Caggia. 2019. "Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria" Microorganisms 7, no. 8: 254. https://doi.org/10.3390/microorganisms7080254
APA StylePino, A., Russo, N., Van Hoorde, K., De Angelis, M., Sferrazzo, G., Randazzo, C. L., & Caggia, C. (2019). Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms, 7(8), 254. https://doi.org/10.3390/microorganisms7080254