Biocontrol of Aspergillus flavus in Ensiled Sorghum by Water Kefir Microorganisms

 ,
,
Abstract
:1. Introduction
2. Results
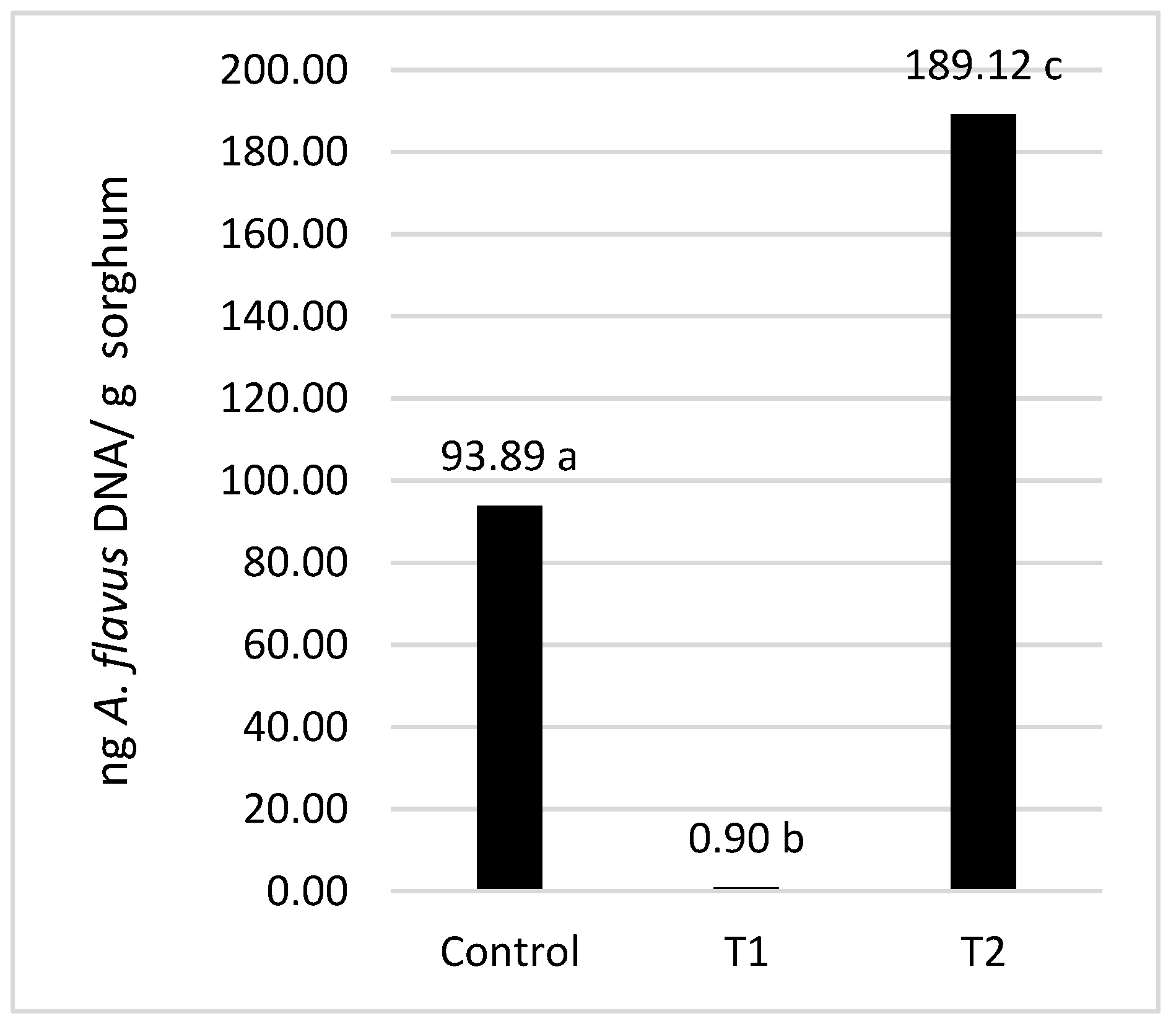
2.1. Quantification of A. flavus DNA by qPCR
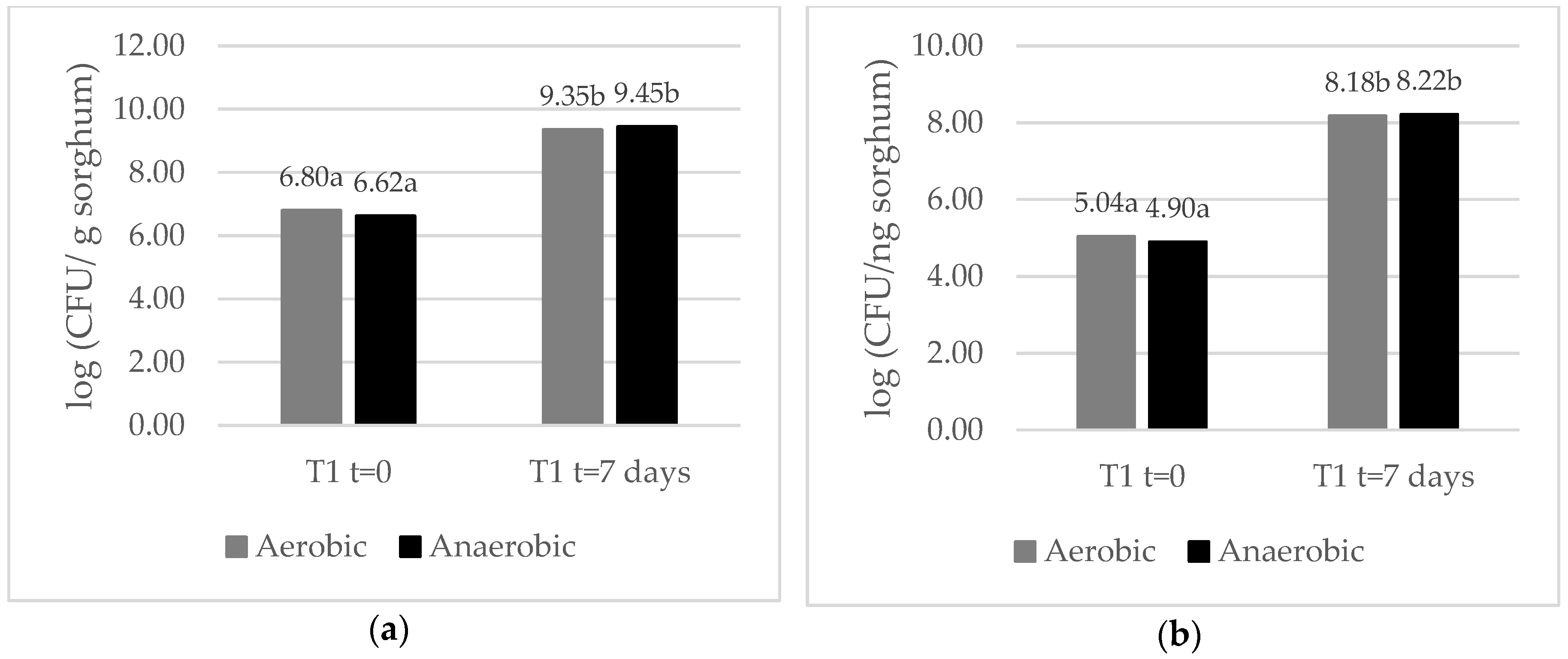
2.2. A. flavus Growth in Mini-Silos
2.3. Microbiological Analysis of T1 Mini-Silos
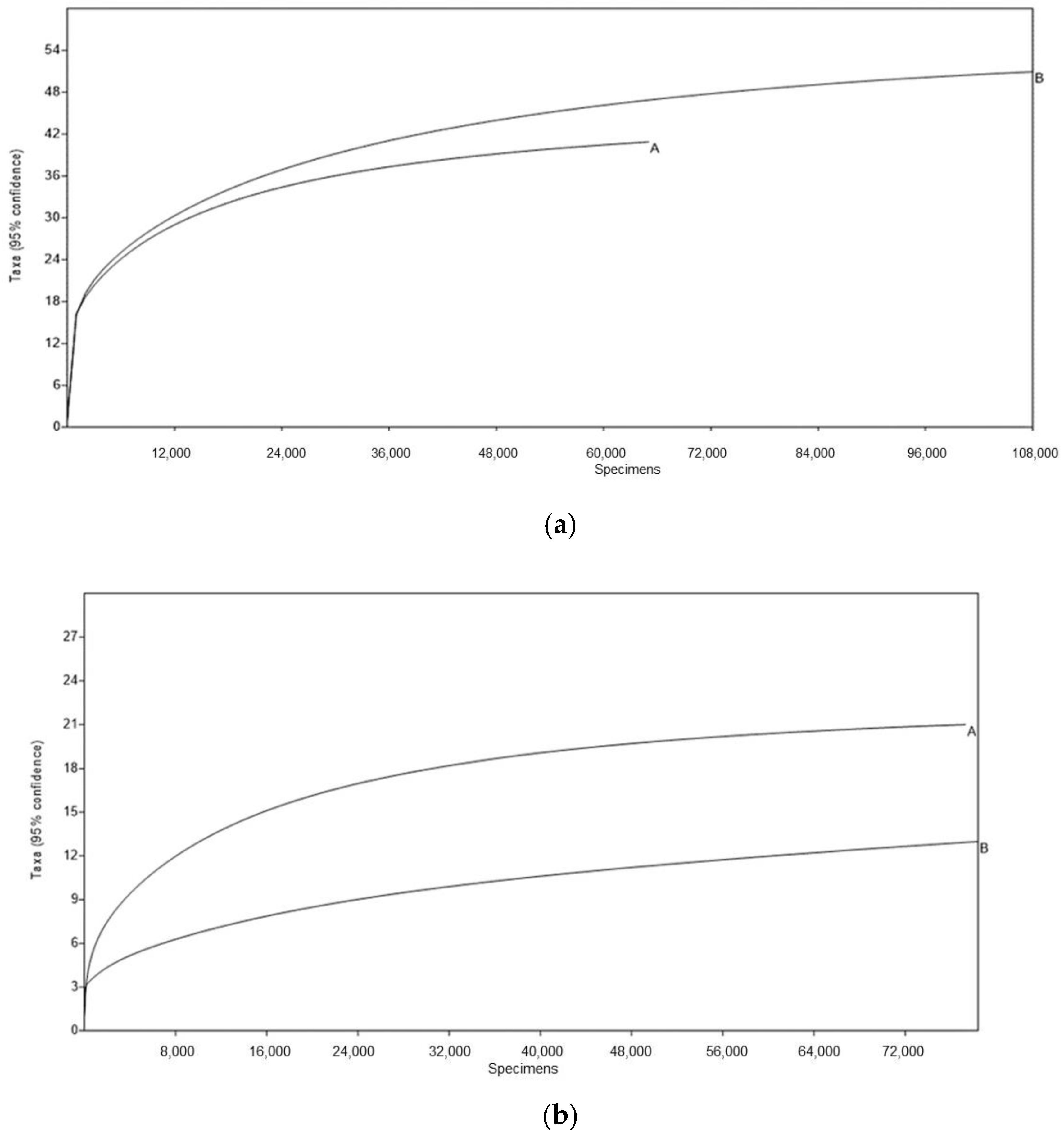
2.4. Data Analysis of High-Throughput Amplicon Sequencing
3. Discussion
4. Materials and Methods
4.1. Pathogen
4.2. Quantification of A. flavus Biomass by qPCR
4.3. Water Kefir Preparation
4.4. Sorghum Grain Preparation and Ensiling
4.5. Analysis of Mini-Silos
4.6. Molecular Identification of Yeast
4.7. Molecular Identification of Bacteria
4.8. Phylogenetic Analysis
4.9. Characterization of the Fungal and Bacterial Microbiota Using Metabarcoding and High-Throughput Sequencing
4.10. Data Analysis of Amplicon Sequences
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Oude Elferink, S.J.W.H.; Spoelstra, S.F. Microbiology of ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J., Eds.; American Society of Agronomy: Madison, WI, USA, 2003; pp. 31–93. ISBN 0891181512. [Google Scholar]
- Scarpitta, N. Qué necesitamos conocer sobre el silo de grano húmedo de sorgo?^ cNicolás Scarpitta. Rev. Plan Agropecu. 2008, 126, 48–54. [Google Scholar]
- Chiossone, J.; Misi, O.; Vicini, R. Available online: http://inta.gob.ar/documentos/silaje-de-sorgo/ (accessed on 1 June 2019).
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Dalcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and mycotoxins in silage: An overview. J. Appl. Microbiol. 2013, 115, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; De Gelder, L.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef] [PubMed]
- García y Santos, C.; Hugo, I.; Suárez, G.; Capella, A.; Domínguez, R.; Sosa, S. Dinámica del pH en microsilos experimentales, con distintos niveles de humedad y el agregado de inoculante. Veterinaria 2012, 48, 142. [Google Scholar]
- Keller, L.; Pereyra, C.M.; Cavaglieri, L.; Keller, K.; Almeida, T.X.; Deveza, M.V.; Assad, R.Q.; Rocha Rosa, C. Fungi and aflatoxin B1 in pre and post fermented sorghum trench type silos destined to bovine intensive-rearing in Brazil. Rev. Bio Ciencias 2012, 2, 81–91. [Google Scholar]
- Rodríguez, A.; Rodríguez, M.; Luque, M.I.; Martín, A.; Córdoba, J.J. Real-time PCR assays for detection and quantification of aflatoxin-producing molds in foods. Food Microbiol. 2012, 31, 89–99. [Google Scholar] [CrossRef]
- Alhaag, H.; Yuan, X.; Mala, A.; Bai, J.; Shao, T. Fermentation Characteristics of Lactobacillus Plantarum and Pediococcus Species Isolated from Sweet Sorghum Silage and Their Application as Silage Inoculants. Appl. Sci. 2019, 9, 1247. [Google Scholar] [CrossRef]
- Gamba, R.R.; Colo, C.N.; Correa, M.; Astoreca, A.; Alconada, T.; De Antoni, G.; Peláez, A.L. Antifungal activity against Aspergillus parasiticus of supernatants from whey permeates fermented with kéfir grains. Adv. Microbiol. 2015, 5, 479–492. [Google Scholar] [CrossRef]
- Passoth, V.; Schnürer, J. Non-conventional yeasts in antifungal application. In Functional Genetics of Industrial Yeasts. Topics in Current Genetics; de Winde, J.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 2, pp. 297–329. ISBN 978-3-540-37003-1. [Google Scholar]
- Druvefors, U.; Schnürer, J. Mold-inhibitory activity of different yeast species during airtight storage of wheat grain. Fems Yeast Res. 2005, 5, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Fredlund, E.; Gidlund, A.; Olsen, M.; Börjesson, T.; Spliid, N.H.H.; Simonsson, M. Method evaluation of Fusarium DNA extraction from mycelia and wheat for down-stream real-time PCR quantification and correlation to mycotoxin levels. J. Microbiol. Methods 2008, 73, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Olstorpe, M.; Borling, J.; Schnürer, J.; Passoth, V. Pichia anomala yeast improves feed hygiene during storage of moist crimped barley grain under Swedish farm conditions. Anim. Feed Sci. Technol. 2010, 156, 47–56. [Google Scholar] [CrossRef]
- Duniere, L.; Jin, L.; Smiley, B.; Qi, M.; Rutherford, W.; Wang, Y.; McAllister, T. Impact of adding Saccharomyces strains on fermentation, aerobic stability, nutritive value, and select lactobacilli populations in corn silage. J. Anim. Sci. 2015, 93, 2322–2335. [Google Scholar] [CrossRef] [PubMed]
- Schnürer, J.; Jonsson, A. Pichia anomala J121: A 30-year overnight near success biopreservation story. Antonie Van Leeuwenhoek 2011, 99, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, A.; Saraf, M. Development of microbial consortia as a biocontrol agent for effective management of fungal diseases in Glycine max L. Arch. Phytopathol. Plant Prot. 2015, 48, 459–474. [Google Scholar] [CrossRef]
- Gamba, R.R.; Caro, C.A.; Martínez, O.L.; Moretti, A.F.; Giannuzzi, L.; De Antoni, G.L.; León Peláez, A. Antifungal effect of kefir fermented milk and shelf life improvement of corn arepas. Int. J. Food Microbiol. 2016, 235, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droby, S.; Wisniewski, M. The fruit microbiome: A new frontier for postharvest biocontrol and postharvest biology. Postharvest Biol. Technol. 2018, 140, 107–112. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Gulitz, A.; Stadie, J.; Wenning, M.; Ehrmann, M.A.; Vogel, R.F. The microbial diversity of water kefir. Int. J. Food Microbiol. 2011, 151, 284–288. [Google Scholar] [CrossRef]
- Laureys, D.; De Vuyst, L. Microbial species diversity, community dynamics, and metabolite kinetics of water Kefir fermentation. Appl. Env. Microbiol. 2014, 80, 2564–2572. [Google Scholar] [CrossRef]
- Guzel-Seydim, Z.B.; Kok-Tas, T.; Greene, A.K.; Seydim, A.C. Review: Functional Properties of Kefir. Crit. Rev. Food Sci. Nutr. 2011, 51, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Dong-Hyeon, K.; Jung-Whan, C.; Hyunsook, K.; Kun-Ho, S. Modulation of intestinal microbiota in mice by kefir administration. Food Sci. Biotechnol. 2015, 24, 1397–1403. [Google Scholar]
- Vinderola, C.G.; Duarte, J.; Thangavel, D.; Perdigón, G.; Farnworth, E.; Matar, C. Immunomodulating capacity of kefir. J. Dairy Res. 2005, 72, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-H.; Jeong, D.; Kim, H.; Kang, I.-B.; Chon, J.-W.; Song, K.-Y.; Seo, K.-H. Antimicrobial Activity of Kefir against Various Food Pathogens and Spoilage Bacteria. Korean J. Food Sci. Anim. Resour. 2016, 36, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismaiel, A.A.; Ghaly, M.F.; El-Naggar, A.K. Milk Kefir: Ultrastructure, Antimicrobial Activity and Efficacy on Aflatoxin B1 Production by Aspergillus flavus. Curr. Microbiol. 2011, 62, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.P.; Rea, M.C.; Hill, C.; Ross, R.P. An application in cheddar cheese manufacture for a strain of Lactococcus lactis producing a novel broad-spectrum bacteriocin, lacticin 3147. Appl. Env. Microbiol. 1996, 62, 612–619. [Google Scholar] [Green Version]
- Rodrigues, K.L.; Caputo, L.R.G.; Carvalho, J.C.T.; Evangelista, J.; Schneedorf, J.M. Antimicrobial and healing activity of kefir and kefiran extract. Int. J. Antimicrob. Agents 2005, 25, 404–408. [Google Scholar] [CrossRef]
- Sun, Z.; Yu, J.; Dan, T.; Zhang, W.; Zhang, H. Phylogenesis and Evolution of Lactic Acid Bacteria. In Lactic Acid Bacteria: Fundamentals and Practice; Zhang, H., Cai, Y., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 1–101. ISBN 978-94-017-8841-0. [Google Scholar]
- De Vos, P.; Garrity, G.; Jones, D.; Krieg, N.; Ludwig, W.; Rainey, F.; Schleifer, K.-H.; Whitman, W. Bergey’s Manual of Systematic Bacteriology. In The Firmicutes, 2nd ed.; Whitman, W.B., Ed.; Springer: New York, NY, USA, 2009; Volune 3, ISBN 978-0-387-95041-9. [Google Scholar]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Shehata, M.G.; Badr, A.N.; El Sohaimy, S.A.; Asker, D.; Awad, T.S. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann. Agric. Sci. 2019. [Google Scholar] [CrossRef]
- Fernandez-Pacheco, P.; Arévalo-Villena, M.; Bevilacqua, A.; Corbo, M.R.; Briones Pérez, A. Probiotic characteristics in Saccharomyces cerevisiae strains: Properties for application in food industries. LWT 2018, 97, 332–340. [Google Scholar] [CrossRef]
- Lutz, M.C.; Lopes, C.A.; Rodriguez, M.E.; Sosa, M.C.; Sangorrín, M.P. Efficacy and putative mode of action of native and commercial antagonistic yeasts against postharvest pathogens of pear. Int. J. Food Microbiol. 2013, 164, 166–172. [Google Scholar] [CrossRef]
- Santos, R.; Buisson, N.; Knight, S.A.B.; Dancis, A.; Camadro, J.-M.; Lesuisse, E. Candida albicans lacking the frataxin homologue: A relevant yeast model for studying the role of frataxin. Mol. Microbiol. 2004, 54, 507–519. [Google Scholar] [CrossRef]
- Druvefors, U.A.; Passoth, V.; Schnürer, J. Nutrient effects on biocontrol of Penicillium roqueforti by Pichia anomala J121 during airtight storage of wheat. Appl. Env. Microbiol. 2005, 71, 1865–1869. [Google Scholar] [CrossRef]
- Fiorda, F.A.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Rakshit, S.K.; Pagnoncelli, M.G.B.; de Souza Vandenberghe, L.P.; Soccol, C.R. Microbiological, biochemical, and functional aspects of sugary kefir fermentation-A review. Food Microbiol. 2017, 66, 86–95. [Google Scholar] [CrossRef]
- Yoo, J.A.; Lim, Y.M.; Yoon, M.H. Production and antifungal effect of 3-phenyllactic acid (PLA) by lactic acid bacteria. J. Appl. Biol. Chem. 2016, 59, 173–178. [Google Scholar] [CrossRef]
- Magnusson, J.; Schnürer, J. Lactobacillus coryniformis subsp. coryniformis Strain Si3 Produces a Broad-Spectrum Proteinaceous Antifungal Compound. Appl. Environ. Microbiol. 2001, 67, 1–5. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torres, M.J.; De Valdez, G.F.; Rollán, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control 2013, 64, 231–237. [Google Scholar] [CrossRef]
- Cleusix, V.; Lacroix, C.; Vollenweider, S.; Duboux, M.; Le Blay, G. Inhibitory activity spectrum of reuterin produced by Lactobacillus reuteri against intestinal bacteria. Bmc Microbiol. 2007, 7, 101. [Google Scholar] [CrossRef]
- Ryu, E.H.; Yang, E.J.; Woo, E.R.; Chang, H.C. Purification and characterization of antifungal compounds from Lactobacillus plantarum HD1 isolated from kimchi. Food Microbiol. 2014, 41, 19–26. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile organic compounds produced by Antarctic strains of Candida sake play a role in the control of postharvest pathogens of apples. Biol. Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Passoth, V.; Fredlund, E.; Druvefors, U.Ä.; Schnürer, J. Biotechnology, physiology and genetics of the yeast Pichia anomala. Fems Yeast Res. 2006, 6, 3–13. [Google Scholar] [CrossRef]
- Muccilli, S.; Restuccia, C. Bioprotective Role of Yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [Green Version]
- Lipinska, L.; Klewicki, R.; Sójka, M.; Bonikowski, R.; Żyżelewicz, D.; Kolodziejczyk, K.; Klewicka, E. Antifungal Activity of Lactobacillus pentosus ŁOCK 0979 in the Presence of Polyols and Galactosyl-Polyols. Probiotics Antimicrob. Proteins 2018, 10, 186–200. [Google Scholar] [CrossRef]
- Shweta, S.; Madhavan, S.; Paranidharan, V.; Velazhahan, R. Detection of Aspergillus flavus in maize kernels by conventional and real-time PCR assays. Int. Food Res. J. 2013, 20, 3329–3335. [Google Scholar]
- Dorak, M.T. Real-Time PCR; Taylor & Francis Group: Oxford, UK, 2006; ISBN 041537734X. [Google Scholar]
- Petersson, S.; Schnurer, J. Biocontrol of mold growth in high-moisture wheat stored under airtight conditions by Pichia anomala, Pichia guilliermondii, and Saccharomyces cerevisiae. Appl. Env. Microbiol. 1995, 61, 1027–1032. [Google Scholar]
- Schena, L.; Ippolito, A.; Zahavi, T.; Cohen, L.; Nigro, F.; Droby, S. Genetic diversity and biocontrol activity of Aureobasidium pullulans isolates against postharvest rots. Postharvest Biol. Technol. 1999, 17, 189–199. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press: San Diego, CA, USA; New York, NY, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Peterson, S.W.; Horn, B.W. Penicillium parvulum and Penicillium georgiense, sp. nov., isolated from the conidial heads of Aspergillus species. Mycologia 2009, 101, 71–83. [Google Scholar] [CrossRef]
- Martinez, A.; Cavello, I.; Garmendia, G.; Rufo, C.; Cavalitto, S.; Vero, S. Yeasts from sub-Antarctic region: Biodiversity, enzymatic activities and their potential as oleaginous microorganisms. Extremophiles 2016, 20, 759–769. [Google Scholar] [CrossRef]
- Hogg, J.C.; Lehane, M.J. Identification of bacterial species associated with the sheep scab mite (Psoroptes ovis) by using amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1999, 65, 4227–4229. [Google Scholar]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H.; Larsson, K.; Gilles, A. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. Plos ONE 2012, 7, e40863. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Yourstone, S.; Mieczkowski, P.; Jones, C.D.; Dangl, J.L. Practical innovations for high-throughput amplicon sequencing. Nat. Methods 2013, 10, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Mahé, F.; Flouri, T.; McDonald, D.; Schloss, P. VSEARCH: a versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi-recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ryan, P.D.; Hammer, Ø.; Harper, D.A.; Paul Ryan, D.D. PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 5–7. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sequences | OTUs | Chao1 | Shannon | |
|---|---|---|---|---|---|
| Bacteria | A | 66571 | 41 | 42.7 | 1.681 |
| B | 109365 | 51 | 52.5 | 1.613 | |
| Fungi | A | 77432 | 21 | 21.2 | 0.1383 |
| B | 78539 | 13 | 16.0 | 0.2657 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonda, M.; Garmendia, G.; Rufo, C.; León Peláez, Á.; Wisniewski, M.; Droby, S.; Vero, S. Biocontrol of Aspergillus flavus in Ensiled Sorghum by Water Kefir Microorganisms. Microorganisms 2019, 7, 253. https://doi.org/10.3390/microorganisms7080253
Gonda M, Garmendia G, Rufo C, León Peláez Á, Wisniewski M, Droby S, Vero S. Biocontrol of Aspergillus flavus in Ensiled Sorghum by Water Kefir Microorganisms. Microorganisms. 2019; 7(8):253. https://doi.org/10.3390/microorganisms7080253
Chicago/Turabian StyleGonda, Mariana, Gabriela Garmendia, Caterina Rufo, Ángela León Peláez, Michael Wisniewski, Samir Droby, and Silvana Vero. 2019. "Biocontrol of Aspergillus flavus in Ensiled Sorghum by Water Kefir Microorganisms" Microorganisms 7, no. 8: 253. https://doi.org/10.3390/microorganisms7080253