Transcriptional Expression of the ompA, cpaf, tarp, and tox Genes of Chlamydia trachomatis Clinical Isolates at Different Stages of the Developmental Cycle

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. C. trachomatis Culture
2.3. Nucleic Acid Extraction and cDNA Synthesis
2.4. Real-Time PCR
3. Results
3.1. Clinical C. trachomatis Isolates
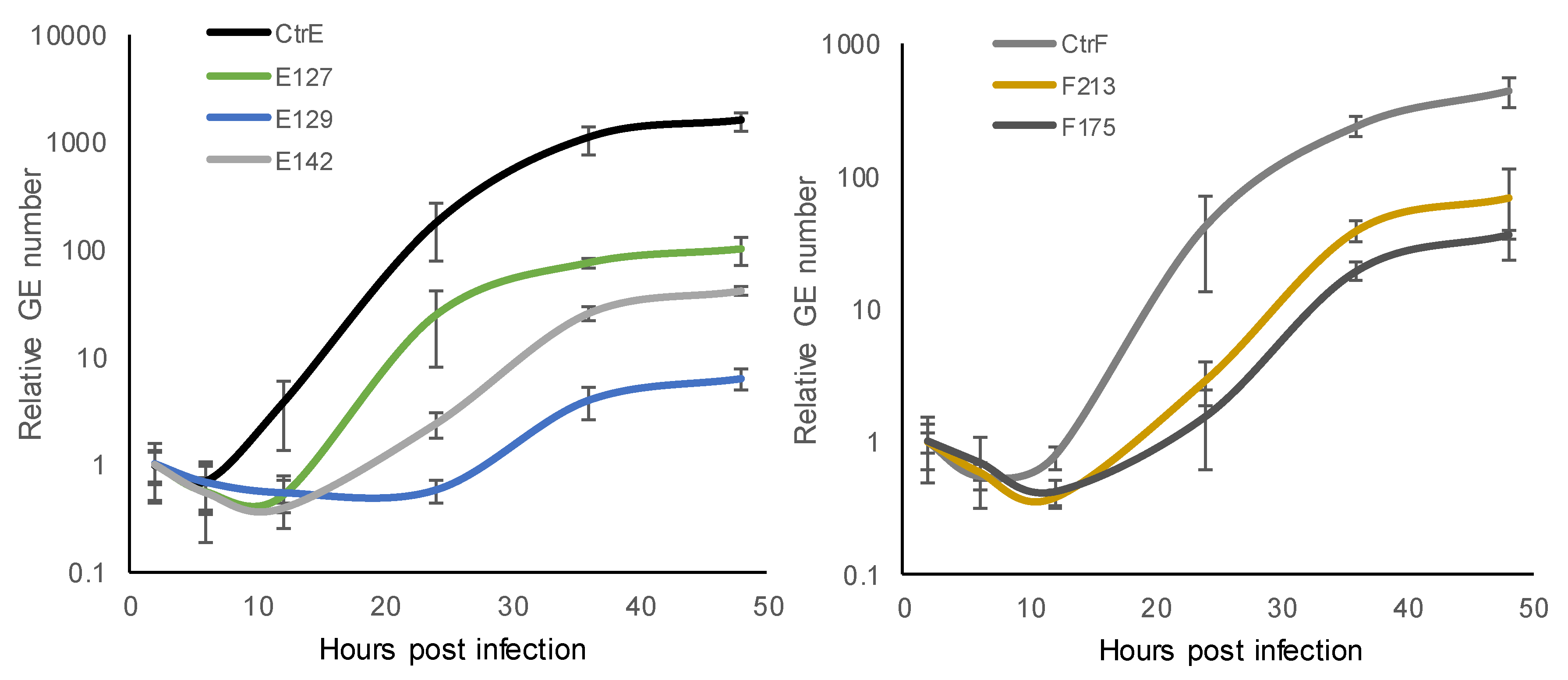
3.2. Growth Kinetics of C. trachomatis Reference Strains and Clinical Isolates
3.3. Performance of C. trachomatis cpaf, tarp, and tox Real-Time PCRs
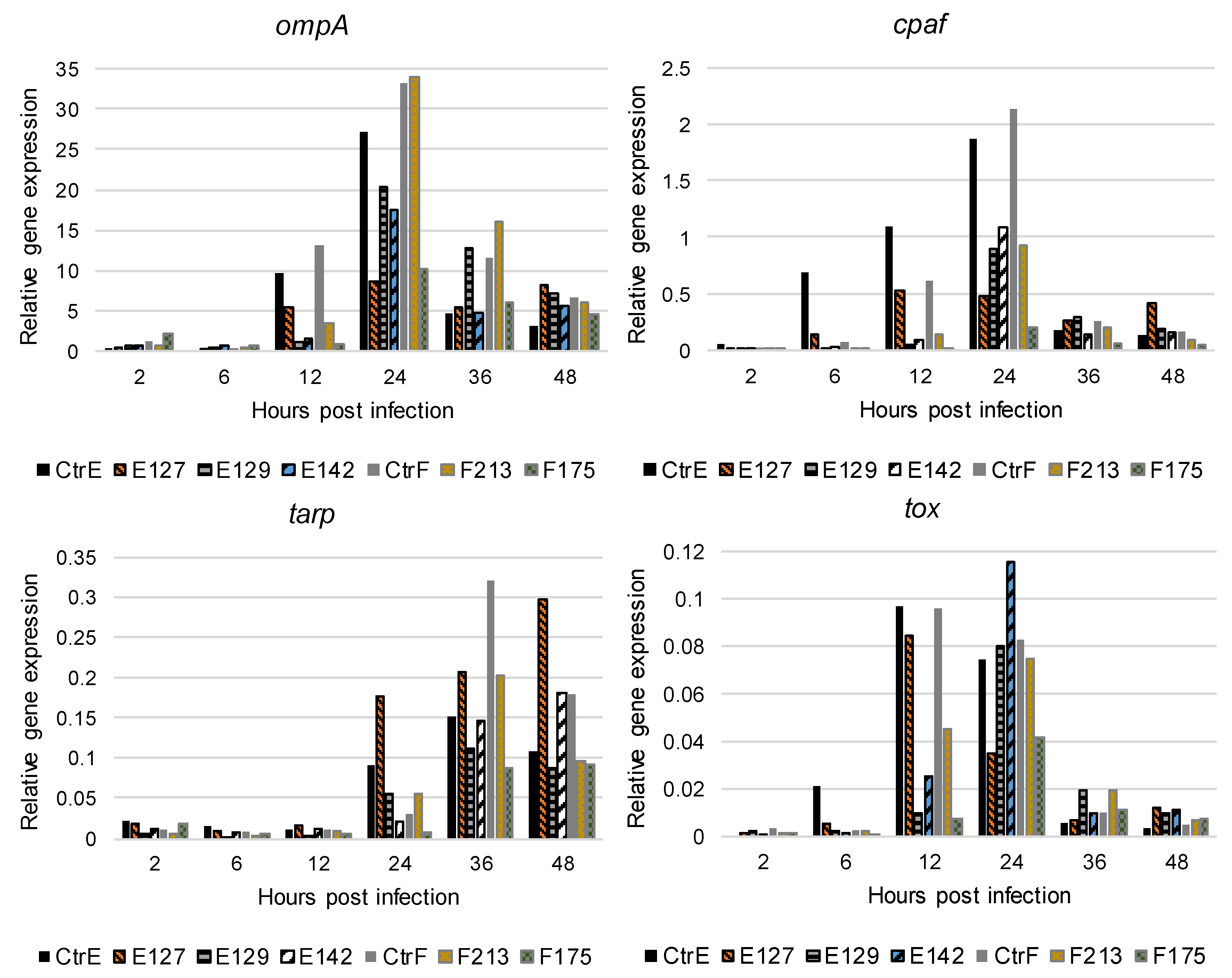
3.4. Expression of C. trachomatis ompA, cpaf, tarp, and tox Genes In Vitro
3.5. Expression of C. trachomatis ompA, cpaf, tarp, and tox Genes in the Cervical Swabs
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, L.; Rowley, J.; Vander Hoorn, S.; Wijesooriya, N.S.; Unemo, M.; Low, N.; Stevens, G.; Gottlieb, S.; Kiarie, J.; Temmerman, M. Global Estimates of the Prevalence and Incidence of Four Curable Sexually Transmitted Infections in 2012 Based on Systematic Review and Global Reporting. PLoS ONE 2015, 10, e0143304. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, C.M.; Ferone, M.E. Chlamydia trachomatis genital infections. Microb. Cell 2016, 3, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Morre, S.A.; Ossewaarde, J.M.; Lan, J.; van Doornum, G.J.; Walboomers, J.M.; MacLaren, D.M.; Meijer, C.J.; van den Brule, A.J. Serotyping and genotyping of genital Chlamydia trachomatis isolates reveal variants of serovars Ba, G, and J as confirmed by omp1 nucleotide sequence analysis. J. Clin. Microbiol. 1998, 36, 345–351. [Google Scholar] [PubMed]
- Tan, M. Temporal Gene Regulation during the Chlamydial Developmental Cycle. In Intracellular Pathogens I Chlamydiales; Tan, M., Bavoil, P.M., Eds.; ASM Press: Washington, DC, USA, 2012; pp. 149–169. [Google Scholar]
- Shaw, E.I.; Dooley, C.A.; Fischer, E.R.; Scidmore, M.A.; Fields, K.A.; Hackstadt, T. Three temporal classes of gene expression during the Chlamydia trachomatis developmental cycle. Mol. Microbiol. 2000, 37, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.L.; Olinger, L.; Chong, K.; Schoolnik, G.; Stephens, R.S. Global stage-specific gene regulation during the developmental cycle of Chlamydia trachomatis. J. Bacteriol. 2003, 185, 3179–3189. [Google Scholar] [CrossRef] [PubMed]
- Belland, R.J.; Zhong, G.; Crane, D.D.; Hogan, D.; Sturdevant, D.; Sharma, J.; Beatty, W.L.; Caldwell, H.D. Genomic transcriptional profiling of the developmental cycle of Chlamydia trachomatis. Proc. Natl. Acad. Sci. USA 2003, 100, 8478–8483. [Google Scholar] [CrossRef] [PubMed]
- Clifton, D.R.; Fields, K.A.; Grieshaber, S.S.; Dooley, C.A.; Fischer, E.R.; Mead, D.J.; Carabeo, R.A.; Hackstadt, T. A chlamydial type III translocated protein is tyrosine-phosphorylated at the site of entry and associated with recruitment of actin. Proc. Natl. Acad. Sci. USA 2004, 101, 10166–10171. [Google Scholar] [CrossRef] [Green Version]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef]
- Zhong, G.; Fan, P.; Ji, H.; Dong, F.; Huang, Y. Identification of a chlamydial protease-like activity factor responsible for the degradation of host transcription factors. J. Exp. Med. 2001, 193, 935–942. [Google Scholar] [CrossRef]
- Snavely, E.A.; Kokes, M.; Dunn, J.D.; Saka, H.A.; Nguyen, B.D.; Bastidas, R.J.; McCafferty, D.G.; Valdivia, R.H. Reassessing the role of the secreted protease CPAF in Chlamydia trachomatis infection through genetic approaches. Pathog. Dis. 2014, 71, 336–351. [Google Scholar] [CrossRef]
- Chen, A.L.; Johnson, K.A.; Lee, J.K.; Sutterlin, C.; Tan, M. CPAF: A Chlamydial protease in search of an authentic substrate. PLoS Pathog. 2012, 8, e1002842. [Google Scholar] [CrossRef] [PubMed]
- Belland, R.J.; Scidmore, M.A.; Crane, D.D.; Hogan, D.M.; Whitmire, W.; McClarty, G.; Caldwell, H.D. Chlamydia trachomatis cytotoxicity associated with complete and partial cytotoxin genes. Proc. Natl. Acad. Sci. USA 2001, 98, 13984–13989. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, J.; Janik, K.; May, M.; Sommer, K.; Ebeling, J.; Hofmann, F.; Genth, H.; Klos, A. Actin re-organization induced by Chlamydia trachomatis serovar D-evidence for a critical role of the effector protein CT166 targeting Rac. PLoS ONE 2010, 5, e9887. [Google Scholar] [CrossRef] [PubMed]
- Bothe, M.; Dutow, P.; Pich, A.; Genth, H.; Klos, A. DXD motif-dependent and -independent effects of the Chlamydia trachomatis cytotoxin CT166. Toxins 2015, 7, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Borges, V.; Ferreira, R.; Nunes, A.; Sousa-Uva, M.; Abreu, M.; Borrego, M.J.; Gomes, J.P. Effect of long-term laboratory propagation on Chlamydia trachomatis genome dynamics. Infect. Genet. Evol. 2013, 17, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Bonner, C.; Caldwell, H.D.; Carlson, J.H.; Graham, M.R.; Kari, L.; Sturdevant, G.L.; Tyler, S.; Zetner, A.; McClarty, G. Chlamydia trachomatis virulence factor CT135 is stable in vivo but highly polymorphic in vitro. Pathog. Dis. 2015, 73, ftv043. [Google Scholar] [CrossRef]
- Borges, V.; Pinheiro, M.; Antelo, M.; Sampaio, D.A.; Vieira, L.; Ferreira, R.; Nunes, A.; Almeida, F.; Mota, L.J.; Borrego, M.J.; et al. Chlamydia trachomatis in vivo to in vitro transition reveals mechanisms of phase variation and down-regulation of virulence factors. PLoS ONE 2015, 10, e0133420. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.; Gomes, J.P.; Mead, S.; Florindo, C.; Correia, H.; Borrego, M.J.; Dean, D. Comparative expression profiling of the Chlamydia trachomatis pmp gene family for clinical and reference strains. PLoS ONE 2007, 2, e878. [Google Scholar] [CrossRef] [PubMed]
- Jalal, H.; Stephen, H.; Alexander, S.; Carne, C.; Sonnex, C. Development of real-time PCR assays for genotyping of Chlamydia trachomatis. J. Clin. Microbiol. 2007, 45, 2649–2653. [Google Scholar] [CrossRef] [PubMed]
- Niemi, S.; Hiltunen-Back, E.; Puolakkainen, M. Chlamydia trachomatis genotypes and the Swedish new variant among urogenital Chlamydia trachomatis strains in Finland. Infect. Dis. Obstet. Gynecol. 2011, 2011, 481890. [Google Scholar] [CrossRef] [PubMed]
- Jalal, H.; Stephen, H.; Curran, M.D.; Burton, J.; Bradley, M.; Carne, C. Development and validation of a rotor-gene real-time PCR assay for detection, identification, and quantification of Chlamydia trachomatis in a single reaction. J. Clin. Microbiol. 2006, 44, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Niemi, S.; Greub, G.; Puolakkainen, M. Chlamydia-related bacteria in respiratory samples in Finland. Microbes Infect. 2011, 13, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Todar´s Online Textbook of Bacteriology. Available online: http://textbookofbacteriology.net/ (accessed on 2 April 2019).
- Borges, V.; Ferreira, R.; Nunes, A.; Nogueira, P.; Borrego, M.J.; Gomes, J.P. Normalization strategies for real-time expression data in Chlamydia trachomatis. J. Microbiol. Methods 2010, 82, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Harris, S.R.; Seth-Smith, H.M.B.; Parmar, S.; Andersson, P.; Giffard, P.M.; Schachter, J.; Moncada, J.; Ellison, L.; Gallo Vaulet, M.L.; et al. Comprehensive global genome dynamics of Chlamydia trachomatis show ancient diversification followed by contemporary mixing and recent lineage expansion. Genome Res. 2017, 27, 1220–1229. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.H.; Hughes, S.; Hogan, D.; Cieplak, G.; Sturdevant, D.E.; McClarty, G.; Caldwell, H.D.; Belland, R.J. Polymorphisms in the Chlamydia trachomatis cytotoxin locus associated with ocular and genital isolates. Infect. Immun. 2004, 72, 7063–7072. [Google Scholar] [CrossRef]
- Miyairi, I.; Mahdi, O.S.; Ouellette, S.P.; Belland, R.J.; Byrne, G.I. Different growth rates of Chlamydia trachomatis biovars reflect pathotype. J. Infect. Dis. 2006, 194, 350–357. [Google Scholar] [CrossRef]
- Belland, R.J.; Nelson, D.E.; Virok, D.; Crane, D.D.; Hogan, D.; Sturdevant, D.; Beatty, W.L.; Caldwell, H.D. Transcriptome analysis of chlamydial growth during IFN-gamma-mediated persistence and reactivation. Proc. Natl. Acad. Sci. USA 2003, 100, 15971–15976. [Google Scholar] [CrossRef]
- Gomes, J.P.; Hsia, R.C.; Mead, S.; Borrego, M.J.; Dean, D. Immunoreactivity and differential developmental expression of known and putative Chlamydia trachomatis membrane proteins for biologically variant serovars representing distinct disease groups. Microbes Infect. 2005, 7, 410–420. [Google Scholar] [CrossRef]
- Ferreira, R.; Borges, V.; Borrego, M.J.; Gomes, J.P. Global survey of mRNA levels and decay rates of Chlamydia trachomatis trachoma and lymphogranuloma venereum biovars. Heliyon 2017, 3, e00364. [Google Scholar] [CrossRef]
- Jha, R.; Vardhan, H.; Bas, S.; Salhan, S.; Mittal, A. Cervical epithelial cells from Chlamydia trachomatis-infected sites coexpress higher levels of chlamydial heat shock proteins 60 and 10 in infertile women than in fertile women. Gynecol. Obstet. Investig. 2009, 68, 160–166. [Google Scholar] [CrossRef]
- Gerard, H.C.; Carter, J.D.; Hudson, A.P. Chlamydia trachomatis is present and metabolically active during the remitting phase in synovial tissues from patients with chronic Chlamydia-induced reactive arthritis. Am. J. Med. Sci. 2013, 346, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, S.P.; Hatch, T.P.; AbdelRahman, Y.M.; Rose, L.A.; Belland, R.J.; Byrne, G.I. Global transcriptional upregulation in the absence of increased translation in Chlamydia during IFNgamma-mediated host cell tryptophan starvation. Mol. Microbiol. 2006, 62, 1387–1401. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, P.; Bailey, L.; Onskog, T.; Bergstrom, S.; Johansson, J. A comparative study of RNA and DNA as internal gene expression controls early in the developmental cycle of Chlamydia pneumoniae. FEMS Immunol. Med. Microbiol. 2010, 58, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Saliba, A.E.; Santos, S.C.; Vogel, J. New RNA-seq approaches for the study of bacterial pathogens. Curr. Opin. Microbiol. 2017, 35, 78–87. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate | Age | Sex | Specimen Type | ompA Genotype | Ctr Serology IgG/IgM/IgA 1 | Clinical Features | Other STIs (Gonorrhea, HIV, or Syphilis) |
|---|---|---|---|---|---|---|---|
| E127 | 27 | F | Cervical swab | E | <32/<20/<20 | Asymptomatic, partner infected, probable first Ctr infection 2 | none |
| E129 | 20 | F | Cervical swab | E | 128/<20/<20 | Asymptomatic, partner infected, probable repeated Ctr infection 3 | none |
| E142 | 20 | F | Cervical swab | E | 32/<20/<20 | Pain in the lower abdomen, vaginal discharge, BV, repeated Ctr infection 4 | none |
| F175 | 24 | F | Cervical swab | F | 32/<20/<20 | Asymptomatic, BV, partner infected, repeated Ctr infection 4 | none |
| F213 | 21 | F | Cervical swab | F | 128/<20/<20 | Asymptomatic, probable repeated Ctr infection 3 | none |
| Ctr Target Gene | Amplicon Length | Reference | Primer/Probe (5′–3′) | |
|---|---|---|---|---|
| cpaf | 86 bp | This study | Forward | TAGGATGGGATCTTGTTCAAAGCT |
| Reverse | CTGCTGGCAAAAACTTGTTGAT | |||
| Probe | 6-FAM-CTGCACAGCAGAAGCTTCGTACACAAGAA-BHQ-1 | |||
| tarp | 108 bp | This study | Forward | CCTCTTCTGGAGATGATTCAGGAA |
| Reverse | TACGCACGGCAGAAAGGATA | |||
| Probe | 6-FAM-CCTCTGTCGGAAATGACGGACCTGCT-BHQ-1 | |||
| tox | 106 bp | This study | Forward | GATTCTTTAATTTCTGCTTGCTGAAA |
| Reverse | TGTTCGATCTCCTCAGTAGGAAGTTT | |||
| Probe | 6-FAM-CTCGGCAATATCAATGACGAAACGCGT-BHQ-1 | |||
| ompA | 219 bp | [22] | Forward | GACTTTGTTTTCGACCGTGTT |
| Reverse | ACARAATACATCAAARCGATCCCA | |||
| Probe | VIC-ATGTTTACVAAYGCYGCTT-MGB-NFQ | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korhonen, S.; Hokynar, K.; Mannonen, L.; Paavonen, J.; Hiltunen-Back, E.; Puolakkainen, M. Transcriptional Expression of the ompA, cpaf, tarp, and tox Genes of Chlamydia trachomatis Clinical Isolates at Different Stages of the Developmental Cycle. Microorganisms 2019, 7, 153. https://doi.org/10.3390/microorganisms7060153
Korhonen S, Hokynar K, Mannonen L, Paavonen J, Hiltunen-Back E, Puolakkainen M. Transcriptional Expression of the ompA, cpaf, tarp, and tox Genes of Chlamydia trachomatis Clinical Isolates at Different Stages of the Developmental Cycle. Microorganisms. 2019; 7(6):153. https://doi.org/10.3390/microorganisms7060153
Chicago/Turabian StyleKorhonen, Suvi, Kati Hokynar, Laura Mannonen, Jorma Paavonen, Eija Hiltunen-Back, and Mirja Puolakkainen. 2019. "Transcriptional Expression of the ompA, cpaf, tarp, and tox Genes of Chlamydia trachomatis Clinical Isolates at Different Stages of the Developmental Cycle" Microorganisms 7, no. 6: 153. https://doi.org/10.3390/microorganisms7060153