A Review: Wolbachia-Based Population Replacement for Mosquito Control Shares Common Points with Genetically Modified Control Approaches


Unit Arboviruses and Insect Vectors, Department of Virology, Institut Pasteur, F-75724 Paris, France
*
Authors to whom correspondence should be addressed.
Pathogens 2020, 9(5), 404; https://doi.org/10.3390/pathogens9050404
Submission received: 15 April 2020
/
Revised: 15 May 2020
/
Accepted: 20 May 2020
/
Published: 22 May 2020
(This article belongs to the Special Issue Untargeted Alternative Routes of Arbovirus Transmission)
{kind=link}
{kind=link}
Abstract
:The growing expansion of mosquito vectors has made mosquito-borne arboviral diseases a global threat to public health, and the lack of licensed vaccines and treatments highlight the urgent need for efficient mosquito vector control. Compared to genetically modified control strategies, the intracellular bacterium Wolbachia, endowing a pathogen-blocking phenotype, is considered an environmentally friendly strategy to replace the target population for controlling arboviral diseases. However, the incomplete knowledge regarding the pathogen-blocking mechanism weakens the reliability of a Wolbachia-based population replacement strategy. Wolbachia infections are also vulnerable to environmental factors, temperature, and host diet, affecting their densities in mosquitoes and thus the virus-blocking phenotype. Here, we review the properties of the Wolbachia strategy as an approach to control mosquito populations in comparison with genetically modified control methods. Both strategies tend to limit arbovirus infections but increase the risk of selecting arbovirus escape mutants, rendering these strategies less reliable.
1. Half of the World’s Population Exposed to Arboviral Diseases
Mosquito-borne arboviral diseases such as chikungunya, dengue, yellow fever, and Zika have been one of the major public health issues over the last few decades, threatening more than half of the world’s population [1]. These arboviruses have dispensed with the need for amplification in wild animals to cause outbreaks in the human population. Human hosts serve simultaneously as the reservoir, amplifier, and disseminator, and the major vectors are the anthropophilic mosquitoes Aedes aegypti and Aedes albopictus [2]. Due to their complex transmission mechanisms with interactions between viruses, mosquito vectors, and vertebrate hosts, all three evolving in changing environments, the control of arboviral diseases is still extremely difficult. The frustrating lack of development of broad-spectrum vaccines against arboviruses has pointed out the importance of new alternatives for arboviral diseases control [3,4].
Chikungunya is a rapidly reemerging arboviral disease. In recent years, as the Indian Ocean lineage (IOL) evolved, this Ae. aegypti adapted chikungunya virus (CHIKV) has caused several outbreaks in tropical countries [5]. The newly emerged CHIKV IOL contains a mutation from Alanine to Valine at position 226 in the E1 protein, which influences the pH threshold for fusion, facilitating the virus entry [6]. Moreover, the E1-A226V mutation also increases the vector competence for Aedes mosquitoes, particularly for Aedes albopictus [7]. Therefore, the epidemic areas have extended beyond tropical regions, reaching the temperate countries in Europe where Ae. albopictus mosquitoes have been established since 1979 [8]; this species is now present in 20 European countries with the highest infestation levels in France [9,10] and Italy [11]. Dengue virus (DENV) alone contributes to approximately 390 million infections and tens of thousands fatal cases annually [12]. There are four DENV serotypes, namely dengue-1, -2, -3, and -4, which trigger distinct immune responses in humans [13]. Although a life-long immunity against a given DENV serotype could be acquired after a first infection, a secondary infection with another DENV serotype could facilitate dengue severe syndrome or dengue hemorrhagic fever (DHF) that increases the burden of this disease even if a secondary infection does not always necessarily lead to DHF [14,15]. Moreover, the nearly 80% of dengue asymptomatic cases enhance the risk of DENV transmission, thus complexifying the disease control. Yellow fever virus (YFV) has not yet become a global threat but heavily affects Sub-Saharan Africa and South America (mainly in Brazil), causing between 94,336 and 118,500 infection incidents annually [16]. In Africa, the urban cycle results from sporadic spillover transmission from the jungle cycle, while in America, human infections are only acquired in forests [17]. Although the YFV live-attenuated vaccine has been available since the 1930s, the global stockpile of YFV vaccines is still an issue due to the production process that requires specific pathogens-free (SPF) eggs; the limited supply of SPF eggs makes it difficult to launch an urgent vaccination campaign [18]. The recent outbreaks in Brazil and the imported cases from Angola to China have raised many concerns on the potential risk of major outbreaks, especially for the immune-naïve populations in Asia [19,20,21]. Zika virus (ZIKV) was first isolated from an infected monkey in Uganda in 1947. Starting from Yap Island in 2007 [22], a much larger outbreak in French Polynesian islands followed [23], and reached Brazil at the end of 2013 [24]. Although ZIKV infection formerly caused only mild illness and was self-limiting [22], more severe outcomes were reported, including microcephaly in newborns and the neurological affections [25,26] giving it the status of a Public Health Emergency of International Concern in 2016 by the World Health Organization (WHO). It should be noted that other emerging arboviruses have not yet caused large outbreaks; they could be the next arboviral threats owing to current global changes, including climate warming and increasing international exchanges [27]. Aedes mosquitoes are also transmitting Mayaro (MAYV; genus: Alphavirus), and Usutu (USUV; genus: Flavivirus) viruses that originated from South America and Sub-Saharan Africa, respectively [28,29]. Since 2000, MAYV has caused several outbreaks in South America, although the number of reported infections remains low. The spread of MAYV to non-endemic areas was reported in Europe after 2008, expanding the risk area of this disease [30]. Similar to MAYV, the USUV was first identified in 1959 [29], but the first human infection was reported at the beginning of 1981 in Africa [31]. Since then, USUV has been introduced into Europe and was repeatedly reported in mosquitoes, birds, and horses in 12 European countries [32].
2. Virus Overcomes Mosquito Immune Barriers to Be Transmitted by Generating Viral Quasispecies
Aedes mosquitoes are the major vectors in transmitting arboviruses. They have expanded their geographic distribution as a consequence of population growth, human activities, and climate change, creating conditions favorable for their proliferation and introducing the means of passive transportation [33]. Their distribution is no longer restricted to tropical regions, spreading to new geographic regions over long distances and stepping up the global impact of arboviral diseases [33].
A mosquito vector acquires an arbovirus by taking a blood meal from a viremic host, after which the virus enters into the midgut epithelial cell and replicates. After infection of the midgut, the virus needs to escape through the midgut basal lamina and disseminate into the hemocele, infecting different tissues or organs including salivary glands. Finally, in a subsequent blood meal, the virus is excreted from the salivary glands and transmitted by the mosquito bite [34].
From the midgut to the salivary glands, the virus encounters different mosquito immune responses, such as RNAi (RNA interference), the Toll, IMD (immune deficiency), and JAK-STAT (Janus kinase signal transducer and activator of transcription protein) pathways, limiting the virus infection, dissemination, and transmission [35]. Viral infections in mosquitoes result from a subtle balance between virus replication and cellular immunity, manifested by a persistent asymptomatic infection in vectors [36]. Considering the error-prone nature of viral RNA-dependent RNA polymerase, RNA viruses generate a collection of variants (namely quasispecies) during replication that facilitates viral adaptation and transmission by mosquitoes [37,38]. Thus, after passing all these filters, only a fraction of the viral population is transmitted to the vertebrate host [39]. This remains dependent on each pairing of virus-mosquito that conditions the mosquito vector competence [34].
3. Population Replacement Strategy to Control Mosquito-Borne Diseases
As the pivot for arboviruses transmission, the mosquito vector is considered the target for efficient arboviral diseases control. Depending on the outcomes, the mosquito population control strategies could be roughly divided into two categories, population reduction and population replacement, which by reducing the target population size or introducing an anti-pathogen phenotype into the target population, respectively, minimizes the contact between arboviruses-carrying mosquitoes and human hosts. The current strategy is to reduce the mosquito vector population with insecticides; however, in the absence of a vector surveillance program, the impact of insecticides on local biodiversity is unpredictable, in addition there is the risk in generating insecticide-resistant mosquitoes that decreases the control efficacy, calling for more species-specific alternative methods [40]. Instead of reducing the target population from the field, which might cause ecological disruption [41,42] and risk of secondary pest emergence, population replacement has been proposed as an alternative [43]. Replacing the target population with a pathogen-refractory strain could specifically reduce the pathogens’ transmission while maintaining the population in its original ecological niche, limiting the risk of secondary pest emergence [43].
A modified genetic-based population replacement approach is composed of an anti-pathogen gene and a gene-drive system, in order to suppress the pathogen replication and to spread the phenotype within the target population [44]. Different mosquito antiviral factors such as siRNA [45,46], miRNA [47,48,49], ribozymes [50,51], immune factors [52], and neutralizing antibodies [53], can act as effectors to reduce the virus infection and transmission in genetically modified mosquitoes. Combined with a proper gene-drive system, the genetically modified mosquitoes expressing a virus-refractory phenotype can replace the wild population in a few generations, that is to say in a few months for mosquitoes. Although the biosafety concerns about using genetically modified insects is still debatable [54], the highly-specific synthetic antiviral immunity used as effectors (RNAi, antiviral ribozymes, overexpressed immune genes, and neutralizing antibodies) has raised issues regarding the selection of escape virus mutants. Thus, strategies generating weaker selection in virus populations might be more sustainable [55].
4. Wolbachia-Based Mosquito Control
The microbiota of mosquito vectors has a strong impact on arbovirus infections [56,57]. The endosymbiont bacteria Wolbachia has been in the spotlight with the discovery of its properties in suppressing the replication of vector-borne human arboviruses such as DENV, YFV, and ZIKV [58,59]. The Wolbachia-based insect control approach is more acceptable than the genetic modification-based approach for the public as it is a naturally existing microbe. Wolbachia is a Gram-negative bacterium, a member of the Alphaproteobacteria (Rickettsiales order). In arthropods, the genome of Wolbachia ranges from 1.2 to 1.6 Mb and contains WO prophages (named after Wolbachia) [60]. It was first discovered in the Culex pipiens mosquito in 1924 by Hertig and Wolbach [61], opening a new avenue of research owing to its high diversity and wide distribution in arthropods [62]. To date, 18 supergroups of Wolbachia have been identified, most of them present in arthropods [63], and more than 65% of insect species harbor Wolbachia [64]. Horizontal transfers of Wolbachia have been demonstrated between species within the same supergroup and, conversely, the same host species can host different Wolbachia strains. Wolbachia bacteria are involved in different symbiotic interactions ranging from parasitic to mutualistic [62]. Mainly transmitted vertically, it intervenes in manipulating the host reproduction in order to optimize its maternal transmission through the eggs. Wolbachia can induce different sex ratio distortion phenotypes in the progeny to favor females: parthenogenesis, feminization, male-killing, and cytoplasmic incompatibility (CI) [65]. In the CI phenotype, Wolbachia-infected females are favored over the non-infected females and males (Figure 1). The molecular mechanisms controlling the CI are today better understood: CI and its rescue are driven by toxin-antidote interactions, whose affinity between partners determines the success of the rescue [65]. For example, in Cx. pipiens during the first embryonic mitosis, cidA and cidB of wPip are the key elements of CI traits [66], where the B factor expresses its toxic effect inducing the CI effect and the A factor acts as an antidote for the rescue. In addition to colonizing reproductive organs, Wolbachia bacteria are also present in somatic tissues; Wolbachia can be acquired from infected embryonic lineages or by passing from cell to cell [67].
5. Wolbachia in Limiting Arbovirus Transmission
Wolbachia can provide fitness advantages for host fertility and/or survival and can alter responses to infections to reduce arbovirus transmission. As an example, the wMel Wolbachia strain artificially introduced in Drosophila melanogaster inhibited Drosophila C virus (DCV) infection, and Wolbachia-infected flies were much more resistant to DCV than the uninfected flies [68,69].
Combining their ability to invade the host population by inducing CI and to interfere negatively with the transmission of disease pathogens, Wolbachia-based control methods have been deployed to prevent the transmission of mosquito-borne diseases [70,71]. The bacterium Wolbachia has been introduced in natural populations of the mosquito Ae. aegypti, the urban vector of dengue, Zika, chikungunya, and yellow fever [72]. Wolbachia bacteria are stably maintained in field mosquito populations and attributed to the pathogen-blocking phenotype [73]. The critical question is whether this effect will persist and if adaptive changes in the mosquito vector, the bacteria, or virus may occur, hampering the success of this strategy.
Wolbachia can share the same niche with the virus, colonizing ovaries, gut, and salivary glands, organs that are essential for the replication and transmission of arboviruses [74]. Usually, high antiviral resistance is associated with high densities of Wolbachia, which could reach several hundred bacteria per cell [75] and cause a significant fitness cost (e.g., reduced fecundity, fertility, and survival) [76]. Mosquito adaptive changes may occur, leading to the evolution towards lower Wolbachia densities and, therefore, a reduction or loss of the antiviral phenotype. Another concern is the evolution of the virus itself; since Wolbachia block the replication of the virus, viral populations could be shaped to overcome such inhibition. Variants able to replicate despite the presence of Wolbachia could be advantaged and spread, weakening the sustainability of the Wolbachia control strategy. As an example, in D. melanogaster, Wolbachia did not present antiviral effects for the Wolbachia-adaptive DCV, which were genetically different from viral populations in Wolbachia-free controls [77]. These findings were obtained from cell cultures, which are far from replicating the conditions in a natural host, with a succession of reduction and restoration of viral diversities after crossing anatomical barriers (midgut and salivary glands) in the mosquito vector [39].
6. The Hypothesized Mechanisms of Wolbachia-Mediated Pathogen Blocking Activity
Several mechanisms have been proposed to explain the molecular basis of the pathogen-blocking phenotype: regulation of immune genes, indirect host gene regulation through other cellular machinery (RNAi, sfRNA), production of reactive oxygen species, or competition for a limited resource such as cholesterol. Growing evidence has shown that the mosquito transcriptome profiles were altered after Wolbachia infection [52,70,78,79]. Insect immune pathways such as Toll, IMD, and JAK/STAT pathways were activated in Wolbachia-infected Ae. aegypti, and led to an efficient reduction in replication of CHIKV, DENV, and Plasmodium [52,70,78,79]. In addition to the immune factors, many genes were reported to be upregulated and subsequently may be able to suppress virus replication; the genes regulating reactive oxygen species production and the upregulation of Wolbachia-mediated methyltransferase are also reported to suppress the replication of DENV in Ae. aegypti [52,80,81]. Mosquito non-coding RNA expression is also influenced upon Wolbachia infection, and might regulate virus replication in infected cells [82]. Although the direct interaction between virus replication and Wolbachia-induced miRNA is not fully understood, an extensively-expressed aae-miRNA-2940 in Wolbachia-infected mosquito cells was proven to upregulate methyltransferase expression and, subsequently, reduce DENV replication [81]. Moreover, this methyltransferase upregulation negatively controls the metalloprotease expression leading to a reduction in West Nile virus replication [83]. Recent studies have suggested that Wolbachia bacteria suppress virus replication through cellular resources allocation (e.g., intracellular space in Wolbachia-infected cells [70,84]), vascular trafficking, and lipid metabolism caused by Wolbachia-mediated cholesterol perturbation [85]. In fact, on one hand, Wolbachia do not have any functional lipopolysaccharide synthase and need cholesterol for membrane formation, while on the other hand, viruses rely on host cholesterol for replication. Therefore, both behave as competitors for access to cholesterol ingested by the mosquito, which is auxotrophic for this element [85]. Moreover, the limited intracellular space in Wolbachia infected cells is also reported to restrict DENV replication [70,84].
7. Viruses Evolve towards an Adaption to Wolbachia-Blocking Activity
Recent studies have raised concerns regarding Wolbachia-mediated selection pressure for arboviruses that might facilitate viral adaptive evolution and escape from pathogen-blocking effects [77]. Evidence has been brought that Wolbachia bacteria do not suppress the replication of viral RNA genomes in mosquito cells and, consequently, are not able to accumulate adaptive viral genomes [86,87]. The same alteration can be observed in Wolbachia-infected D. melanogaster for DCV, where Wolbachia-mediated selective DCV strains could be found after 10 passages in Wolbachia-infected flies. Despite the fact that the adapted DCV strains did not exhibit the same resistance phenotype to Wolbachia in newly Wolbachia-infected flies, evidence points to the risk of Wolbachia-mediated selection for virus adaptive mutations. The resulting adapted viruses might have a better chance of escaping the Wolbachia-mediated virus-blocking effects in drosophila [77]. Moreover, many studies have indicated the Wolbachia naturally infecting Ae. albopictus, wAlbA, and wAlbB do not show significant pathogen-blocking activity against virus infection in this mosquito [75,88,89,90,91]. Even if wAlbB is transinfected in other mosquito species, it cannot trigger the pathogen-blocking phenotype, as observed with other Wolbachia strains [92,93]. These results raise the question of the outcome of co-evolution between Wolbachia and viruses, highlighting the risk for viruses to escape from Wolbachia-mediated immunity (Figure 2a).
8. Environmental Factors That May Affect Wolbachia-Based Control Programs
8.1. An Abiotic Factor, the Temperature
In the field, Wolbachia-infected mosquitoes are exposed (naturally or not) to fluctuating and even extreme temperatures (Figure 2b). Larvae reared under cycling temperatures between 26 °C and 37 °C resulted in adults with a lower density of Wolbachia, an effect persisting across generations [94], and a diminished CI effect that resulted in the inability of Wolbachia to be transmitted vertically [95]. More interesting is the possibility that these effects could be strain-dependent. Temperature significantly affects wMel and wMelPop-CLA in Ae. aegypti, whereas no effect was observed with wAlbB, which naturally colonizes the mosquito Ae. albopictus, suggesting that the Wolbachia adaptation might reduce the pathogen-blocking activity. Infections with wAlbB seem to be more applicable in a population reduction strategy than pathogen suppression in field conditions. The duration of heat stress also alters Wolbachia density [96]. A long duration of heat stress of Ae. aegypti immature stages leads to lower wMel densities in adults. Complementary studies using local mosquitoes and field temperatures are required in Wolbachia-released sites [97].
Temperature might influence Wolbachia density in Ae. aegypti through bacteriophage WO infection [98]. High temperatures may reduce Wolbachia densities in Ae. aegypti through interactions with WO, which infects wMel [99] and wAlbB [100]. This phage undergoes cycles of lysogenic and lytic phases; heat shock triggers the lytic phase during which the phage replicates and causes Wolbachia lysis, reducing its densities [101]. However, the temperature has a positive effect on virus replication with significantly higher titers of DENV at temperature > 28 °C in Ae. aegypti mosquitoes at day 10 post-infection in salivary glands [102]. More globally, high temperature is likely to weaken the Wolbachia-mediated pathogen-blocking activity by shortening the extrinsic incubation period (the time necessary for mosquitoes to become infectious after an infectious blood meal) [103,104,105], and reducing Wolbachia density through bacteriophage WO.
8.2. A Biotic Factor, the Host Diet for Mosquito Vectors
Host diet notably influences Wolbachia load (Figure 2c) [106]. Flies reared with yeast-enriched diets show reduced Wolbachia loads in the host female germline, and flies with a high sucrose diet show an increase of Wolbachia titer in oocytes. Thus, Wolbachia bacteria rely upon host uptake of amino acids and carbohydrates. Exposure to yeast-enriched food alters Wolbachia nucleoid morphology in oogenesis [106]. The yeast-induced Wolbachia depletion is mediated by the somatic target of rapamycin (TOR) and insulin signaling pathways. These findings are critical in programs combating arboviral diseases by releasing Wolbachia-infected vectors [73,107]. Thus, the natural environment including host diet (i.e., nature of blood and sugar absorbed by released mosquitoes) should be considered when evaluating the efficiency of Wolbachia strategies in the field.
9. Obstacles for Population Replacement Program
9.1. Unclear Wolbachia Distribution in Wild Mosquito Populations
Ae. aegypti populations were thought to be free of naturally-harboring Wolbachia; as a result, many Wolbachia-based mosquito control strategies were deployed worldwide to reduce or replace the target mosquito populations via the CI nature of Wolbachia. However, accumulating evidence shows the existence of naturally-occurring Wolbachia in wild Ae. aegypti populations in India [108], Malaysia [109], Panama [110], Thailand [111], Philippines [112], and the United States [113,114,115]. Even though only small proportions of Ae. aegypti were reported in naturally-harboring Wolbachia in field-collected populations, the presence of naturally-infecting Wolbachia tends to increase the instability for replacing the target populations, thus weakening the efficacy of mosquito control programs.
9.2. The Robust CI Effect Limits the Solutions for Wolbachia Re-Replacement
On the one hand, the naturally existing Wolbachia in the target population might be an obstacle for the mosquito population replacement program. On the other hand, the robust CI effect also increases the difficulty to re-replace the target population that has been already replaced by Wolbachia-harboring mosquitoes. Because of this, the Wolbachia-mediated pathogen-blocking strategy implemented can deviate from the objectives initially planned. Thus, a more flexible replacement program is required to adapt to this scenario. The results of computational modeling have suggested that unless a correct Wolbachia strain is used, the selection could favor the strain with a lower fitness cost (commensal infection), thus replacing the existing Wolbachia-harboring population. Furthermore, the second replacement can be less efficient than the first one [116].
10. Conclusions
Mosquito control, as an essential step for mosquito-borne diseases transmission management, either by reducing the target population size or replacing the target population with a pathogen-refractory strain, could efficiently reduce the contact between mosquitoes and hosts, thereby interrupting the disease transmission. Compared to a genetically modified control strategy, a Wolbachia-based control strategy is considered a promising alternative to control mosquito populations due to its natural and environmentally friendly features: It can carry a broad pathogen-blocking activity and robust CI effect, which together ensure the efficacy in reducing mosquito-borne diseases transmission. However, complex interactions and resulting co-evolution processes among mosquito, virus, host, and the environment, make difficult any mosquito-borne disease control strategies. Thus, monitoring and prevention programs to avoid escape mutants in viral populations must be attentively planned if the targeted objective is to reach a sustainable control strategy.
Author Contributions
P.-S.Y. and A.-B.F. wrote the paper. Both authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the French Government’s Investissement d’Avenir program, Laboratoire d’Excellence “Integrative Biology of Emerging Infectious Diseases, IBEID” (grant n_ANR-10-LABX-62-IBEID) (A.-B.F.) and the National Health Research Institutes (NHRI-PP-108-0324-01-17-07) (P.-S.Y.).
Acknowledgments
We thank Marie Vazeille, Adrien Blisnick, and Rachel Bellone for discussions. We also thank Laura Manuellan for help in making figures and Peter Sahlins for editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; De Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antiviral Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flasche, S.; Wilder-Smith, A.; Hombach, J.; Smith, P.G. Estimating the proportion of vaccine-induced hospitalized dengue cases among Dengvaxia vaccinees in the Philippines. Wellcome Open Res. 2019, 4, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder-Smith, A.; Flasche, S.; Smith, P.G. Vaccine-attributable severe dengue in the Philippines. Lancet 2019, 394, 2151–2152. [Google Scholar] [CrossRef] [Green Version]
- Powers, A.M.; Logue, C.H. Changing patterns of chikungunya virus: Re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007, 88, 2363–2377. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef]
- Adhami, J.; Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus skuse (Diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 1998, 14, 340–343. [Google Scholar]
- Delisle, E.; Rousseau, C.; Broche, B.; Leparc-Goffart, I.; L’Ambert, G.; Cochet, A.; Prat, C.; Foulongne, V.; Ferre, J.B.; Catelinois, O.; et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Euro Surveill. 2015, 20. [Google Scholar] [CrossRef] [Green Version]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souares, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef]
- Angelini, R.; Finarelli, A.C.; Angelini, P.; Po, C.; Petropulacos, K.; Macini, P.; Fiorentini, C.; Fortuna, C.; Venturi, G.; Romi, R.; et al. An outbreak of chikungunya fever in the province of Ravenna, Italy. Euro Surveill. 2007, 12. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.S. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: An historical perspective and role of antibody-dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Kliks, S.C.; Nisalak, A.; Brandt, W.E.; Wahl, L.; Burke, D.S. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am. J. Trop Med. Hyg. 1989, 40, 444–451. [Google Scholar] [CrossRef]
- Shearer, F.M.; Longbottom, J.; Browne, A.J.; Pigott, D.M.; Brady, O.J.; Kraemer, M.U.G.; Marinho, F.; Yactayo, S.; De Araujo, V.E.M.; Da Nobrega, A.A.; et al. Existing and potential infection risk zones of yellow fever worldwide: A modelling analysis. Lancet Glob. Health 2018, 6, e270–e278. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.I.O.; Sacchetto, L.; De Rezende, I.M.; Trindade, G.S.; LaBeaud, A.D.; De Thoisy, B.; Drumond, B.P. Recent sylvatic yellow fever virus transmission in Brazil: The news from an old disease. Virol. J. 2020, 17, 9. [Google Scholar] [CrossRef] [Green Version]
- WHO Expert Committee on Biological Standardization. Forty-fifth report. World Health Organ. Tech. Rep. Ser. 1995, 858, 1–101. [Google Scholar]
- Schlagenhauf, P.; Chen, L.H. Yellow Fever importation to China - a failure of pre- and post-travel control systems? Int. J. Infect. Dis. 2017, 60, 91–92. [Google Scholar] [CrossRef] [Green Version]
- Wilder-Smith, A.; Lee, V.; Gubler, D.J. Yellow fever: Is Asia prepared for an epidemic? Lancet Infect. Dis. 2019, 19, 241–242. [Google Scholar] [CrossRef] [Green Version]
- Wasserman, S.; Tambyah, P.A.; Lim, P.L. Yellow fever cases in Asia: Primed for an epidemic. Int. J. Infect. Dis. 2016, 48, 98–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Bossin, H.; Mallet, H.P.; Besnard, M.; Broult, J.; Baudouin, L.; Levi, J.E.; Sabino, E.C.; Ghawche, F.; Lanteri, M.C.; et al. Zika virus in French Polynesia 2013-14: Anatomy of a completed outbreak. Lancet Infect. Dis. 2018, 18, e172–e182. [Google Scholar] [CrossRef]
- Faria, N.R.; Azevedo, R.d.S.d.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Paixao, E.S.; Leong, W.Y.; Rodrigues, L.C.; Wilder-Smith, A. Asymptomatic Prenatal Zika Virus Infection and Congenital Zika Syndrome. Open Forum. Infect. Dis. 2018, 5, 073. [Google Scholar] [CrossRef]
- Moghadas, S.M.; Shoukat, A.; Espindola, A.L.; Pereira, R.S.; Abdirizak, F.; Laskowski, M.; Viboud, C.; Chowell, G. Asymptomatic Transmission and the Dynamics of Zika Infection. Sci. Rep. 2017, 7, 5829. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Anderson, C.R.; Downs, W.G.; Wattley, G.H.; Ahin, N.W.; Reese, A.A. Mayaro virus: A new human disease agent. II. Isolation from blood of patients in Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1957, 6, 1012–1016. [Google Scholar] [CrossRef]
- Engel, D.; Jost, H.; Wink, M.; Borstler, J.; Bosch, S.; Garigliany, M.M.; Jost, A.; Czajka, C.; Luhken, R.; Ziegler, U.; et al. Reconstruction of the Evolutionary History and Dispersal of Usutu Virus, a Neglected Emerging Arbovirus in Europe and Africa. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodriguez, Y.; Pacheco, Y.; Anaya, J.M.; Ramirez-Santana, C. Mayaro: An emerging viral threat? Emerg. Microbes Infect. 2018, 7, 163. [Google Scholar] [CrossRef]
- Nikolay, B.; Diallo, M.; Boye, C.S.; Sall, A.A. Usutu virus in Africa. Vector Borne Zoonotic Dis. 2011, 11, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, U.; Ye, J.; Ruan, X.; Wan, S.; Zhu, B.; Cao, S. Usutu virus: An emerging flavivirus in Europe. Viruses 2015, 7, 219–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ebel, G.D. Dynamics of flavivirus infection in mosquitoes. Adv. Virus Res. 2003, 60, 187–232. [Google Scholar] [PubMed]
- Sim, S.; Jupatanakul, N.; Dimopoulos, G. Mosquito immunity against arboviruses. Viruses 2014, 6, 4479–4504. [Google Scholar] [CrossRef] [Green Version]
- Blair, C.D. Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Patterson, E.I.; Khanipov, K.; Rojas, M.M.; Kautz, T.F.; Rockx-Brouwer, D.; Golovko, G.; Albayrak, L.; Fofanov, Y.; Forrester, N.L. Mosquito bottlenecks alter viral mutant swarm in a tissue and time-dependent manner with contraction and expansion of variant positions and diversity. Virus Evol. 2018, 4, 001. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Weger-Lucarelli, J.; Murrieta, R.A.; Fauver, J.R.; Garcia-Luna, S.M.; Prasad, A.N.; Black, W.C.t.; Ebel, G.D. Genetic Drift during Systemic Arbovirus Infection of Mosquito Vectors Leads to Decreased Relative Fitness during Host Switching. Cell Host Microbe 2016, 19, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Coffey, L.L.; Forrester, N.; Tsetsarkin, K.; Vasilakis, N.; Weaver, S.C. Factors shaping the adaptive landscape for arboviruses: Implications for the emergence of disease. Future Microbiol. 2013, 8, 155–176. [Google Scholar] [CrossRef] [Green Version]
- Flores, H.A.; O’Neill, S.L. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Fang, J. Ecology: A world without mosquitoes. Nature 2010, 466, 432–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostera, G.R.; Gostin, L.O. Biosafety concerns involving genetically modified mosquitoes to combat malaria and dengue in developing countries. JAMA 2011, 305, 930–931. [Google Scholar] [CrossRef] [Green Version]
- Carballar-Lejarazu, R.; James, A.A. Population modification of Anopheline species to control malaria transmission. Pathog. Glob. Health 2017, 111, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Champer, J.; Buchman, A.; Akbari, O.S. Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 2016, 17, 146–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, A.W.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [Green Version]
- Mathur, G.; Sanchez-Vargas, I.; Alvarez, D.; Olson, K.E.; Marinotti, O.; James, A.A. Transgene-mediated suppression of dengue viruses in the salivary glands of the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2010, 19, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Yen, P.S.; James, A.; Li, J.C.; Chen, C.H.; Failloux, A.B. Synthetic miRNAs induce dual arboviral-resistance phenotypes in the vector mosquito Aedes aegypti. Commun. Biol. 2018, 1, 11. [Google Scholar] [CrossRef]
- Ramyasoma, H.; Dassanayake, R.S.; Hapugoda, M.; Capurro, M.L.; Silva Gunawardene, Y.I.N. Multiple dengue virus serotypes resistant transgenic Aedes aegypti fitness evaluated under laboratory conditions. RNA Biol. 2020, 1–21. [Google Scholar] [CrossRef]
- Buchman, A.; Gamez, S.; Li, M.; Antoshechkin, I.; Li, H.H.; Wang, H.W.; Chen, C.H.; Klein, M.J.; Duchemin, J.B.; Paradkar, P.N.; et al. Engineered resistance to Zika virus in transgenic Aedes aegypti expressing a polycistronic cluster of synthetic small RNAs. Proc. Natl. Acad. Sci. USA 2019, 116, 3656–3661. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Furey, C.; Balaraman, V.; Fraser, M.J. Antiviral Hammerhead Ribozymes Are Effective for Developing Transgenic Suppression of Chikungunya Virus in Aedes aegypti Mosquitoes. Viruses 2016, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.R.; Keith, J.H.; Fraser, T.S.; Dawson, J.L.; Kucharski, C.A.; Horne, K.M.; Higgs, S.; Fraser, M.J., Jr. Effective suppression of dengue virus using a novel group-I intron that induces apoptotic cell death upon infection through conditional expression of the Bax C-terminal domain. Virol. J. 2014, 11, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [Green Version]
- Buchman, A.; Gamez, S.; Li, M.; Antoshechkin, I.; Li, H.H.; Wang, H.W.; Chen, C.H.; Klein, M.J.; Duchemin, J.B.; Crowe, J.E., Jr.; et al. Broad dengue neutralization in mosquitoes expressing an engineered antibody. PLoS Pathog. 2020, 16, e1008103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okorie, P.N.; Marshall, J.M.; Akpa, O.M.; Ademowo, O.G. Perceptions and recommendations by scientists for a potential release of genetically modified mosquitoes in Nigeria. Malar J. 2014, 13, 154. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, L.; Saleh, M.C. Manipulating Mosquito Tolerance for Arbovirus Control. Cell Host Microbe 2019, 26, 309–313. [Google Scholar] [CrossRef]
- Cirimotich, C.M.; Ramirez, J.L.; Dimopoulos, G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 2011, 10, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Minard, G.; Tran, F.H.; Raharimalala, F.N.; Hellard, E.; Ravelonandro, P.; Mavingui, P.; Valiente Moro, C. Prevalence, genomic and metabolic profiles of Acinetobacter and Asaia associated with field-caught Aedes albopictus from Madagascar. FEMS Microbiol. Ecol. 2013, 83, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Bourtzis, K.; Dobson, S.L.; Xi, Z.; Rasgon, J.L.; Calvitti, M.; Moreira, L.A.; Bossin, H.C.; Moretti, R.; Baton, L.A.; Hughes, G.L.; et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. 2014, 132, S150–S163. [Google Scholar] [CrossRef]
- Dutra, H.L.C.; Rocha, M.N.; Dias, F.B.S.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host Microbe 2016, 19, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Masui, S.; Kamoda, S.; Sasaki, T.; Ishikawa, H. Distribution and evolution of bacteriophage WO in Wolbachia, the endosymbiont causing sexual alterations in arthropods. J. Mol. Evol. 2000, 51, 491–497. [Google Scholar] [CrossRef]
- Hertig, M.; Wolbach, S.B. Studies on Rickettsia-Like Micro-Organisms in Insects. J. Med. Res. 1924, 44, 329–374.7. [Google Scholar]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Landmann, F. The Wolbachia Endosymbionts. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, M.; Landmann, F.; Labbe, P.; Justy, F.; Weill, M.; Sicard, M. The cellular phenotype of cytoplasmic incompatibility in Culex pipiens in the light of cidB diversity. PLoS Pathog. 2018, 14, e1007364. [Google Scholar] [CrossRef]
- Beckmann, J.F.; Ronau, J.A.; Hochstrasser, M. A Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility. Nat. Microbiol. 2017, 2, 17007. [Google Scholar] [CrossRef]
- Pietri, J.E.; DeBruhl, H.; Sullivan, W. The rich somatic life of Wolbachia. Microbiologyopen 2016, 5, 923–936. [Google Scholar] [CrossRef]
- Osborne, S.E.; Leong, Y.S.; O’Neill, S.L.; Johnson, K.N. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog. 2009, 5, e1000656. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e2. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef]
- Zouache, K.; Voronin, D.; Tran-Van, V.; Mousson, L.; Failloux, A.B.; Mavingui, P. Persistent Wolbachia and cultivable bacteria infection in the reproductive and somatic tissues of the mosquito vector Aedes albopictus. PLoS ONE 2009, 4, e6388. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef]
- Martinez, J.; Ok, S.; Smith, S.; Snoeck, K.; Day, J.P.; Jiggins, F.M. Should Symbionts Be Nice or Selfish? Antiviral Effects of Wolbachia Are Costly but Reproductive Parasitism Is Not. PLoS Pathog. 2015, 11, e1005021. [Google Scholar] [CrossRef]
- Martinez, J.; Bruner-Montero, G.; Arunkumar, R.; Smith, S.C.L.; Day, J.P.; Longdon, B.; Jiggins, F.M. Virus evolution in Wolbachia-infected Drosophila. Proc. Biol. Sci. 2019, 286, 20192117. [Google Scholar] [CrossRef] [Green Version]
- Kambris, Z.; Cook, P.E.; Phuc, H.K.; Sinkins, S.P. Immune activation by life-shortening Wolbachia and reduced filarial competence in mosquitoes. Science 2009, 326, 134–136. [Google Scholar] [CrossRef] [Green Version]
- Kambris, Z.; Blagborough, A.M.; Pinto, S.B.; Blagrove, M.S.; Godfray, H.C.; Sinden, R.E.; Sinkins, S.P. Wolbachia stimulates immune gene expression and inhibits plasmodium development in Anopheles gambiae. PLoS Pathog. 2010, 6, e1001143. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.B.L.; Magalhaes Alves, D.; Bottino-Rojas, V.; Pereira, T.N.; Sorgine, M.H.F.; Caragata, E.P.; Moreira, L.A. Wolbachia and dengue virus infection in the mosquito Aedes fluviatilis (Diptera: Culicidae). PLoS ONE 2017, 12, e0181678. [Google Scholar] [CrossRef]
- Zhang, G.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [Green Version]
- Mayoral, J.G.; Etebari, K.; Hussain, M.; Khromykh, A.A.; Asgari, S. Wolbachia infection modifies the profile, shuttling and structure of microRNAs in a mosquito cell line. PLoS ONE 2014, 9, e96107. [Google Scholar] [CrossRef] [Green Version]
- Slonchak, A.; Hussain, M.; Torres, S.; Asgari, S.; Khromykh, A.A. Expression of mosquito microRNA Aae-miR-2940-5p is downregulated in response to West Nile virus infection to restrict viral replication. J. Virol. 2014, 88, 8457–8467. [Google Scholar] [CrossRef] [Green Version]
- Rainey, S.M.; Martinez, J.; McFarlane, M.; Juneja, P.; Sarkies, P.; Lulla, A.; Schnettler, E.; Varjak, M.; Merits, A.; Miska, E.A.; et al. Wolbachia Blocks Viral Genome Replication Early in Infection without a Transcriptional Response by the Endosymbiont or Host Small RNA Pathways. PLoS Pathog. 2016, 12, e1005536. [Google Scholar] [CrossRef] [Green Version]
- Geoghegan, V.; Stainton, K.; Rainey, S.M.; Ant, T.H.; Dowle, A.A.; Larson, T.; Hester, S.; Charles, P.D.; Thomas, B.; Sinkins, S.P. Perturbed cholesterol and vesicular trafficking associated with dengue blocking in Wolbachia-infected Aedes aegypti cells. Nat. Commun. 2017, 8, 526. [Google Scholar] [CrossRef]
- Koh, C.; Audsley, M.D.; Di Giallonardo, F.; Kerton, E.J.; Young, P.R.; Holmes, E.C.; McGraw, E.A. Sustained Wolbachia-mediated blocking of dengue virus isolates following serial passage in Aedes aegypti cell culture. Virus Evol. 2019, 5, 012. [Google Scholar] [CrossRef] [Green Version]
- Teramoto, T.; Huang, X.; Armbruster, P.A.; Padmanabhan, R. Infection of Aedes albopictus Mosquito C6/36 Cells with the wMelpop Strain of Wolbachia Modulates Dengue Virus-Induced Host Cellular Transcripts and Induces Critical Sequence Alterations in the Dengue Viral Genome. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Blagrove, M.S.; Arias-Goeta, C.; Di Genua, C.; Failloux, A.B.; Sinkins, S.P. A Wolbachia wMel transinfection in Aedes albopictus is not detrimental to host fitness and inhibits Chikungunya virus. PLoS Negl. Trop. Dis. 2013, 7, e2152. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.A.; Vythilingam, I.; Lim, Y.A.L.; Zabari, N.; Lee, H.L. Detection of Wolbachia in Aedes albopictus and Their Effects on Chikungunya Virus. Am. J. Trop. Med. Hyg. 2017, 96, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Mousson, L.; Zouache, K.; Arias-Goeta, C.; Raquin, V.; Mavingui, P.; Failloux, A.B. The native Wolbachia symbionts limit transmission of dengue virus in Aedes albopictus. PLoS Negl. Trop. Dis. 2012, 6, e1989. [Google Scholar] [CrossRef]
- Mousson, L.; Martin, E.; Zouache, K.; Madec, Y.; Mavingui, P.; Failloux, A.B. Wolbachia modulates Chikungunya replication in Aedes albopictus. Mol. Ecol. 2010, 19, 1953–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Xu, Y.; Dimopoulos, G.; Xi, Z. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef]
- Foo, I.J.; Hoffmann, A.A.; Ross, P.A. Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes. Trop. Med. Infect. Dis. 2019, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.A.; Ritchie, S.A.; Axford, J.K.; Hoffmann, A.A. Loss of cytoplasmic incompatibility in Wolbachia-infected Aedes aegypti under field conditions. PLoS Negl. Trop. Dis. 2019, 13, e0007357. [Google Scholar] [CrossRef]
- Ulrich, J.N.; Beier, J.C.; Devine, G.J.; Hugo, L.E. Heat Sensitivity of wMel Wolbachia during Aedes aegypti Development. PLoS Negl. Trop. Dis. 2016, 10, e0004873. [Google Scholar] [CrossRef]
- Ye, Y.H.; Carrasco, A.M.; Dong, Y.; Sgro, C.M.; McGraw, E.A. The Effect of Temperature on Wolbachia-Mediated Dengue Virus Blocking in Aedes aegypti. Am. J. Trop. Med. Hyg. 2016, 94, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Gavotte, L.; Henri, H.; Stouthamer, R.; Charif, D.; Charlat, S.; Bouletreau, M.; Vavre, F. A Survey of the bacteriophage WO in the endosymbiotic bacteria Wolbachia. Mol. Biol. Evol. 2007, 24, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Gavotte, L.; Vavre, F.; Henri, H.; Ravallec, M.; Stouthamer, R.; Bouletreau, M. Diversity, distribution and specificity of WO phage infection in Wolbachia of four insect species. Insect Mol. Biol. 2004, 13, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Chauvatcharin, N.; Ahantarig, A.; Baimai, V.; Kittayapong, P. Bacteriophage WO-B and Wolbachia in natural mosquito hosts: Infection incidence, transmission mode and relative density. Mol. Ecol. 2006, 15, 2451–2461. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Bordenstein, S.R. Temperature affects the tripartite interactions between bacteriophage WO, Wolbachia, and cytoplasmic incompatibility. PLoS ONE 2011, 6, e29106. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.H.; Chen, T.H.; Lin, C.; Shu, P.Y.; Su, C.L.; Teng, H.J. The impact of temperature and Wolbachia infection on vector competence of potential dengue vectors Aedes aegypti and Aedes albopictus in the transmission of dengue virus serotype 1 in southern Taiwan. Parasit Vectors 2017, 10, 551. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Fluctuations at a low mean temperature accelerate dengue virus transmission by Aedes aegypti. PLoS Negl. Trop. Dis. 2013, 7, e2190. [Google Scholar] [CrossRef]
- Dohm, D.J.; O’Guinn, M.L.; Turell, M.J. Effect of environmental temperature on the ability of Culex pipiens (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef]
- Mourya, D.T.; Yadav, P.; Mishra, A.C. Effect of temperature stress on immature stages and susceptibility of Aedes aegypti mosquitoes to chikungunya virus. Am. J. Trop. Med. Hyg. 2004, 70, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Serbus, L.R.; White, P.M.; Silva, J.P.; Rabe, A.; Teixeira, L.; Albertson, R.; Sullivan, W. The impact of host diet on Wolbachia titer in Drosophila. PLoS Pathog. 2015, 11, e1004777. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Balaji, S.; Jayachandran, S.; Prabagaran, S.R. Evidence for the natural occurrence of Wolbachia in Aedes aegypti mosquitoes. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Teo, C.; Lim, P.; Voon, K.; Mak, J. Detection of dengue viruses and Wolbachia in Aedes aegypti and Aedes albopictus larvae from four urban localities in Kuala Lumpur, Malaysia. Trop. Biomed. 2017, 34, 14. [Google Scholar]
- Bennett, K.L.; Gomez-Martinez, C.; Chin, Y.; Saltonstall, K.; McMillan, W.O.; Rovira, J.R.; Loaiza, J.R. Dynamics and diversity of bacteria associated with the disease vectors Aedes aegypti and Aedes albopictus. Sci. Rep. 2019, 9, 12160. [Google Scholar] [CrossRef]
- Thongsripong, P.; Chandler, J.A.; Green, A.B.; Kittayapong, P.; Wilcox, B.A.; Kapan, D.D.; Bennett, S.N. Mosquito vector-associated microbiota: Metabarcoding bacteria and eukaryotic symbionts across habitat types in Thailand endemic for dengue and other arthropod-borne diseases. Ecol. Evol. 2018, 8, 1352–1368. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, T.M.; Hashimoto, K.; Harnandika, R.K.; Amalin, D.M.; Watanabe, K. Detection of Wolbachia in field-collected Aedes aegypti mosquitoes in metropolitan Manila, Philippines. Parasit Vectors 2019, 12, 361. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, A.; Yu, W.; Jiang, J.; Sanchez, C.; Karna, A.K.; Martinez, K.J.L.; Hanley, K.A.; Buenemann, M.; Hansen, I.A.; Xue, R.D.; et al. Wolbachia pipientis occurs in Aedes aegypti populations in New Mexico and Florida, USA. Ecol. Evol. 2019, 9, 6148–6156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, K.L.; Brown, M.R.; Strand, M.R. Mosquitoes host communities of bacteria that are essential for development but vary greatly between local habitats. Mol. Ecol. 2016, 25, 5806–5826. [Google Scholar] [CrossRef] [Green Version]
- Hegde, S.; Khanipov, K.; Albayrak, L.; Golovko, G.; Pimenova, M.; Saldana, M.A.; Rojas, M.M.; Hornett, E.A.; Motl, G.C.; Fredregill, C.L.; et al. Microbiome Interaction Networks and Community Structure From Laboratory-Reared and Field-Collected Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus Mosquito Vectors. Front. Microbiol. 2018, 9, 2160. [Google Scholar] [CrossRef] [Green Version]
- Crain, P.R.; Crowley, P.H.; Dobson, S.L. Wolbachia re-replacement without incompatibility: Potential for intended and unintended consequences. J. Med. Entomol. 2013, 50, 1152–1158. [Google Scholar] [CrossRef]
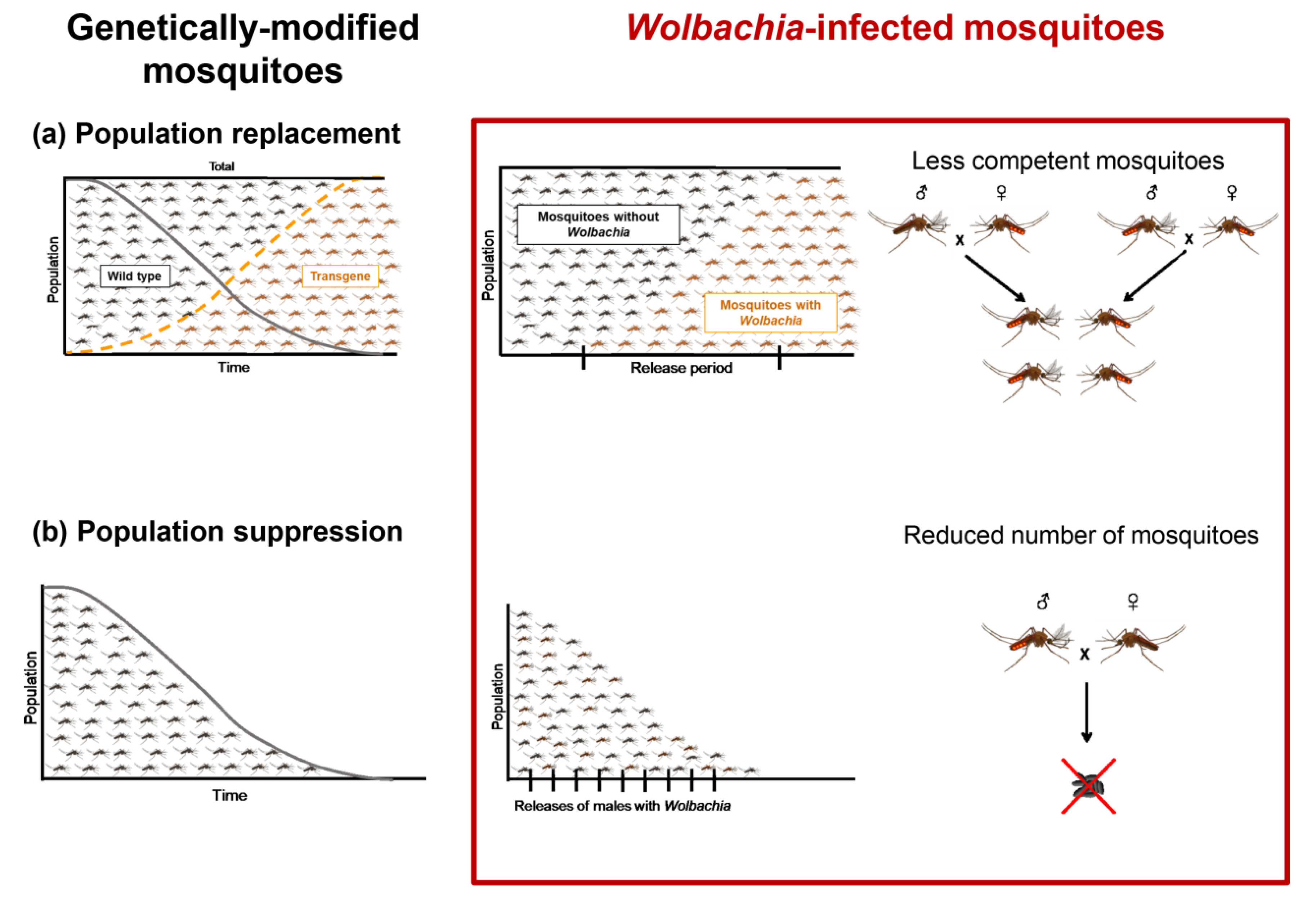
Figure 1.
Wolbachia-based mosquito control strategy can be viewed as a genetically modified control approach, population replacement, or population suppression. (a) The population replacement strategy is to release Wolbachia-infected female mosquitoes that, after mating with males (Wolbachia-infected or not), produce viable offspring, allowing a wide spread of Wolbachia in the field population that harbors less competent individuals, even as the total number of mosquitoes remains unchanged. (b) Population suppression aims to release Wolbachia-infected male mosquitoes that, after mating with wild females, do not produce viable offspring, thus reducing the total number of mosquitoes.
Figure 1.
Wolbachia-based mosquito control strategy can be viewed as a genetically modified control approach, population replacement, or population suppression. (a) The population replacement strategy is to release Wolbachia-infected female mosquitoes that, after mating with males (Wolbachia-infected or not), produce viable offspring, allowing a wide spread of Wolbachia in the field population that harbors less competent individuals, even as the total number of mosquitoes remains unchanged. (b) Population suppression aims to release Wolbachia-infected male mosquitoes that, after mating with wild females, do not produce viable offspring, thus reducing the total number of mosquitoes.

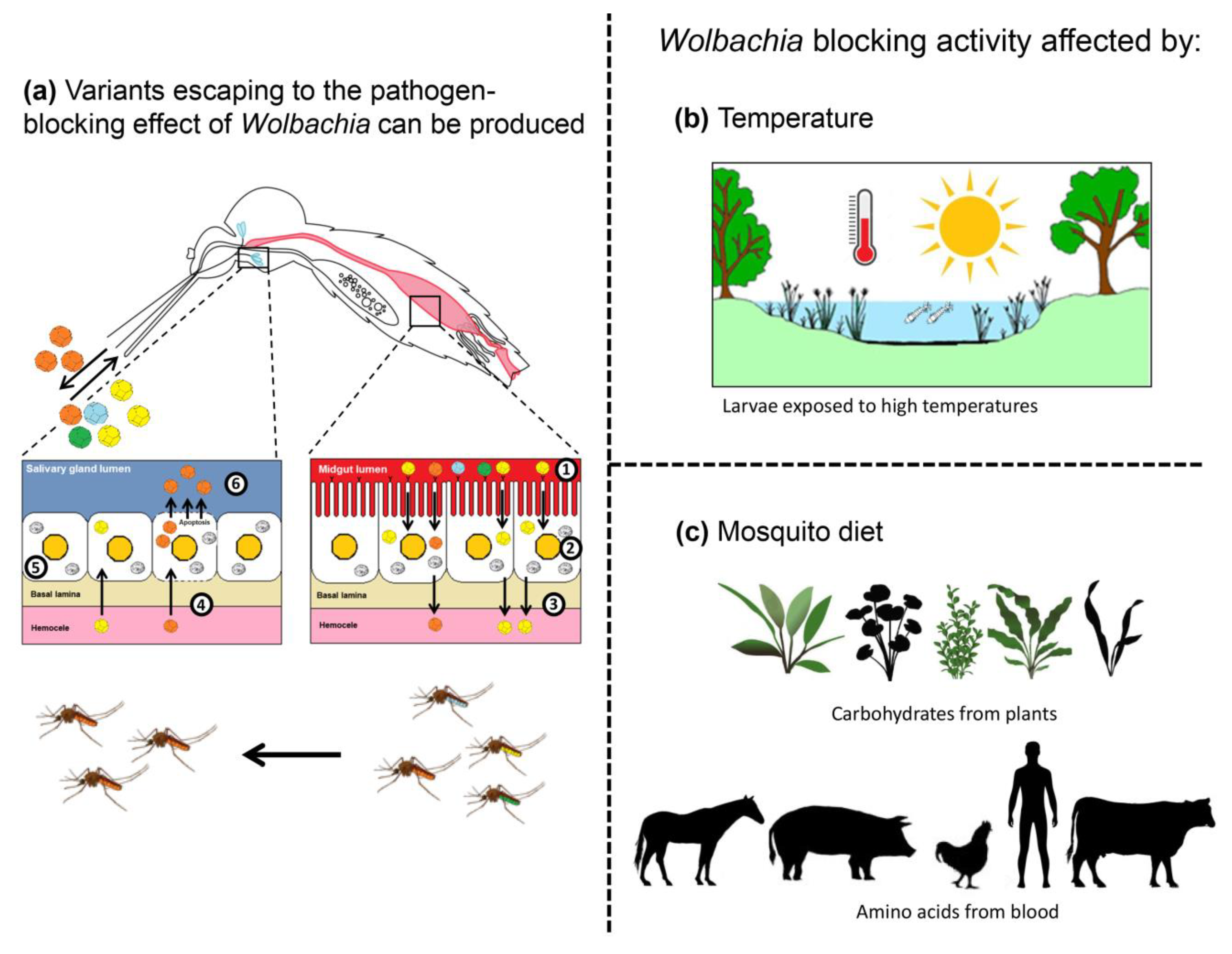
Figure 2.
Viruses escaping from Wolbachia blocking activity. (a) After ingestion of an infectious blood meal (1), viruses enter into the midgut epithelial cells and replicate (2). Newly produced viruses are released into the hemocoel, infecting internal organs/tissues (3). After reaching the salivary glands (4), viruses replicate (5), and new viruses are excreted with saliva expectorated (6) by females when they bite. In the presence of Wolbachia in the midgut and salivary glands, escaping variants can be produced, leading mosquitoes to transmit viruses less sensitive to the pathogen-blocking effect. (b) Wolbachia blocking activity can be altered by environmental factors such as temperature. Mosquito larvae submitted to high temperatures produce adults with a lower density of Wolbachia. (c) Mosquito adults feeding on enriched diets (carbohydrates and amino acids) show reduced Wolbachia loads. Both (b) and (c) lead to a diminished blocking activity of Wolbachia as their bacterial densities are much lower.
Figure 2.
Viruses escaping from Wolbachia blocking activity. (a) After ingestion of an infectious blood meal (1), viruses enter into the midgut epithelial cells and replicate (2). Newly produced viruses are released into the hemocoel, infecting internal organs/tissues (3). After reaching the salivary glands (4), viruses replicate (5), and new viruses are excreted with saliva expectorated (6) by females when they bite. In the presence of Wolbachia in the midgut and salivary glands, escaping variants can be produced, leading mosquitoes to transmit viruses less sensitive to the pathogen-blocking effect. (b) Wolbachia blocking activity can be altered by environmental factors such as temperature. Mosquito larvae submitted to high temperatures produce adults with a lower density of Wolbachia. (c) Mosquito adults feeding on enriched diets (carbohydrates and amino acids) show reduced Wolbachia loads. Both (b) and (c) lead to a diminished blocking activity of Wolbachia as their bacterial densities are much lower.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yen, P.-S.; Failloux, A.-B. A Review: Wolbachia-Based Population Replacement for Mosquito Control Shares Common Points with Genetically Modified Control Approaches. Pathogens 2020, 9, 404. https://doi.org/10.3390/pathogens9050404
AMA Style
Yen P-S, Failloux A-B. A Review: Wolbachia-Based Population Replacement for Mosquito Control Shares Common Points with Genetically Modified Control Approaches. Pathogens. 2020; 9(5):404. https://doi.org/10.3390/pathogens9050404
Chicago/Turabian StyleYen, Pei-Shi, and Anna-Bella Failloux. 2020. "A Review: Wolbachia-Based Population Replacement for Mosquito Control Shares Common Points with Genetically Modified Control Approaches" Pathogens 9, no. 5: 404. https://doi.org/10.3390/pathogens9050404
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.