The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi


 , and
, and
Abstract
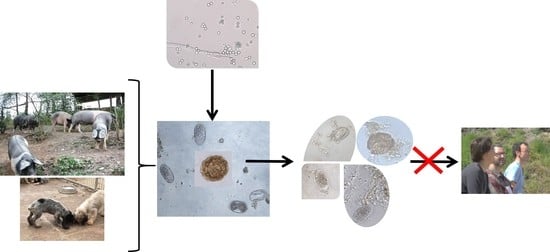
:
1. Introduction
2. Results
2.1. Recovery Rates of Eggs of STHs
2.2. Viability of Eggs of STHs
2.2.1. Ascaris suum
2.2.2. Lemurostrongylus sp.
2.3. Development of Eggs of STHs
2.3.1. Ascaris suum
2.3.2. Lemurostrongylus sp.
2.4. Effect of SSF (Soil Saprophytic Fungi) on the Risk of Infection by Eggs and Larvae of STHs
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Spores of SFF (Soil Filamentous Fungi)
4.3. Soil-Transmitted Helminths (STHs)
4.3.1. Ascaris suum
- Control: boxes provided with 2 mL of water.
- CR: sprayed with 2 mL containing 2 × 105 spores of C. rosea.
- TA: sprayed with 2 mL of 4 × 105 spores of T. atrobrunneum.
4.3.2. Lemurostrongylus sp.
- Control: boxes provided with 2 mL of water.
- CR: sprayed with 2 mL containing 2 × 105 spores of C. rosea.
- TA: sprayed with 2 mL of 4 × 105 spores of T. atrobrunneum.
4.4. Analysis of the Effect of SSF on the Eggs of STHs
4.5. Reduction of Soil Contamination by STHs
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, B.; Maya, C.; Velásquez, G.; Torner, F.; Arambula, F.; Barrios, J.A.; Velasco, M. Identification and quantification of pathogenic helminth eggs using a digital image system. Exp. Parasitol. 2016, 166, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lustigman, S.; Prichard, R.K.; Gazzinelli, A.; Grant, W.N.; Boatin, B.A.; McCarthy, J.S.; Basáñez, M.G. A research agenda for helminth diseases of humans: The problem of helminthiases. PLoS Negl. Trop. Dis. 2012, 6, e1582. [Google Scholar] [CrossRef] [Green Version]
- Inpankaew, T.; Schär, F.; Dalsgaard, A.; Khieu, V.; Chimnoi, W.; Chhoun, C.; Sok, D.; Marti, H.; Muth, S.; Odermatt, P.; et al. High Prevalence of Ancylostoma ceylanicum Hookworm Infections in Humans, Cambodia, 2012. Emerg. Infect. Dis. 2014, 20, 976–982. [Google Scholar] [CrossRef] [PubMed]
- WHO. Integrating Neglected Tropical Diseases into Global Health and Development: Fourth Report on Neglected Tropical Diseases. 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/255011/9789241565448-eng.pdf?sequence=1&isAllowed=y (accessed on 18 August 2020).
- Mascarini-Serra, L. Prevention of Soil-transmitted Helminth Infection. J. Glob. Infect. Dis. 2011, 3, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Noon, J.B.; Aroian, R.V. Recombinant subunit vaccines for soil-transmitted helminths. Parasitology 2017, 144, 1845–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saari, S.; Näreaho, A.; Nikander, S. Canine Parasites and Parasitic Diseases; Saari, S., Näreaho, A., Eds.; Academic Press: Oxford, UK, 2018; Chapter 5; pp. 83–149. [Google Scholar]
- Saelens, G.; Gabriël, S. Currently Available Monitoring and Surveillance Systems for Taenia spp., Echinococcus spp., Schistosoma spp., and Soil-Transmitted Helminths at the Control/Elimination Stage: A Systematic Review. Pathogens 2020, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Graeff-Teixeira, C.; Morassutti, A.L.; Kazacos, K.R. Update on Baylisascariasis, a Highly Pathogenic Zoonotic Infection. Clin. Microbiol. Rev. 2016, 29, 375–399. [Google Scholar] [CrossRef] [Green Version]
- Jia, T.W.; Melville, S.; Utzinger, J.; King, C.H.; Zhou, X.N. Soil-transmitted helminth reinfection after drug treatment: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2012, 6, e1621. [Google Scholar] [CrossRef] [Green Version]
- Soares, F.E.; Braga, F.R.; de Araújo, J.V.; Lima, W.; de Queiroz, J.H. The nematophagous fungus Monacrosporium thaumasium and its nematicidal activity on Angiostrongylus vasorum. Rev. Iberoam. Micol. 2015, 32, 51–53. [Google Scholar] [CrossRef]
- Bilotto, F.; Fusé, L.A.; Sagües, M.F.; Iglesias, L.E.; Fernández, A.S.; Zegbi, S.; Guerrero, I.; Saumell, C.A. Predatory effect of Duddingtonia flagrans on infective larvae of gastro-intestinal parasites under sunny and shaded conditions. Exp. Parasitol. 2018, 193, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Vázquez-Ruiz, R.A.; Cazapal-Monteiro, C.F.; Valderrábano, E.; Arroyo, F.L.; Francisco, I.; Miguélez, S.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Isolation of ovicidal fungi from fecal samples of captive animals maintained in a zoological park. J. Fungi 2017, 3, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapa, S.; Thamsborg, S.M.; Wang, R.; Meyling, N.V.; Dalgaard, T.S.; Petersen, H.H.; Mejer, H. Effect of the nematophagous fungus Pochonia chlamydosporia on soil content of ascarid eggs and infection levels in exposed hens. Parasit. Vectors 2018, 11, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, Í.S.; Oliveira, I.C.; Campos, A.K.; Araújo, J.V. Association and predatory capacity of fungi Pochonia chlamydosporia and Arthrobotrys cladodes in the biological control of parasitic helminths of bovines. Parasitology 2020, 146, 1347–1351. [Google Scholar] [CrossRef]
- Hernández, J.Á.; Cazapal-Monteiro, C.F.; Sanchís, J.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Potential usefulness of filamentous fungi to prevent zoonotic soil-transmitted helminths. Vector Borne Zoonotic Dis. 2018, 18, 690–696. [Google Scholar] [CrossRef]
- Evans, B.R.; Leighton, F.A. A history of One Health. Rev. Sci. Tech. Off. Int. Epiz. 2014, 33, 413–420. [Google Scholar] [CrossRef]
- Tinkler, S.H. Preventive chemotherapy and anthelmintic resistance of soil-transmitted helminths—Can we learn nothing from veterinary medicine? One Health 2019, 9, 100106. [Google Scholar] [CrossRef]
- Cortiñas, F.J.; Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L.; Miguélez, S.; Suárez, J.; López De Arellano, M.E.; Sánchez-Andrade, R.; Mendoza De Gives, P.; Paz-Silva, A.; et al. Potential use of Mucor circinelloides for the biological control of certain helminths affecting livestock reared in a care farm. Biocontrol. Sci. Technol. 2015, 25, 1443–1452. [Google Scholar] [CrossRef]
- Hernández, J.Á.; Cazapal-Monteiro, C.F.; Arroyo, F.L.; Silva, M.I.; Palomero, A.M.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Biological control of soil transmitted helminths (STHs) in a zoological park by using saprophytic fungi. Biol. Control 2018, 122, 24–30. [Google Scholar] [CrossRef]
- Mendoza-de Gives, P.; López-Arellano, M.E.; Aguilar-Marcelino, L.; Olazarán-Jenkins, S.; Reyes-Guerrero, D.; Ramírez-Várgas, G.; Vega-Murillo, V.E. The nematophagous fungus Duddingtonia flagrans reduces the gastrointestinal parasitic nematode larvae population in faeces of orally treated calves maintained under tropical conditions-Dose/response assessment. Vet. Parasitol. 2018, 263, 66–72. [Google Scholar] [CrossRef]
- Traversa, D. Pet Roundworms and Hookworms: A Continuing Need for Global Worming. Parasit. Vectors 2012, 5, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andes, A.L.; Paller, V.G.V. Effect of various composting methods on the concentration and viability of Ascaris suum eggs in organic fertilisers. Pertanika J. Trop. Agric. Sci. 2018, 41, 687–698. [Google Scholar]
- Paller, V.; Babia-Abion, S. Soil-transmitted helminth (STH) eggs contaminating soils in selected organic and conventional farms in the Philippines. Parasite Epidemiol. Control 2019, 7, e00119. [Google Scholar] [CrossRef] [PubMed]
- Kłapeć, T.; Borecka, A. Contamination of vegetables, fruits and soil with geohelmints eggs on organic farms in Poland. Ann. Agric. Environ. Med. 2012, 19, 421–425. [Google Scholar] [PubMed]
- Ordoñez, K.; Paller, V.G.; Goh, X.T.; Lim, Y. Parasite contamination of vegetables from selected organic and conventional farms in Laguna and Benguet Province, Philippines. Trop. Agric. Sci. 2018, 41, 1741–1746. [Google Scholar]
- Kaplan, R.M.; Nielsen, M.K. An evidence-based approach to equine parasite control: It ain’t the 60s anymore. Equine Vet. Educ. 2010, 22, 306–316. [Google Scholar] [CrossRef]
- Vercruysse, J.; Albonico, M.; Behnke, J.M.; Kotze, A.C.; Prichard, R.K.; McCarthy, J.S.; Montresor, A.; Levecke, B. Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int. J. Parasitol. Drugs Drug Resist. 2011, 1, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.; Ellis, K.A.; McGoldrick, J.; Jonsson, N.N.; Stear, M.J.; Forbes, A.B. Targeted anthelmintic treatment of parasitic gastroenteritis in first grazing season dairy calves using daily live weight gain as an indicator. Vet. Parasitol. 2017, 244, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.A.; Arroyo, F.L.; Suárez, J.; Cazapal-Monteiro, C.; Romasanta, A.; López-Arellano, M.E.; Pedreira, J.; Madeira de Carvalho, L.M.; Sánchez-Andrade, R.; Arias, M.S.; et al. Feeding horses with industrially manufactured pellets with fungal spores to promote nematode integrated control. Vet. Parasitol. 2016, 229, 37–44. [Google Scholar] [CrossRef]
- Palomero, A.M.; Cazapal-Monteiro, C.F.; Valderrábano, E.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Soil Fungi enable the control of gastrointestinal nematodes in wild bovidae captive in a zoological park: A 4-Year trial. Parasitology 2020, 147, 791–798. [Google Scholar] [CrossRef]
- Voinot, M.; Cazapal-Monteiro, C.; Hernández, J.; Palomero, A.; Arroyo Balán, F.; Sanchís, J.; Pedreira, J.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M. Integrating the control of helminths in dairy cattle: Deworming, rotational grazing and nutritional pellets with parasiticide fungi. Vet. Parasitol. 2020, 278, 109038. [Google Scholar] [CrossRef] [PubMed]
- Levecke, B.; Mekonnen, Z.; Albonico, M.; Vercruysse, J. The impact of baseline faecal egg counts on the efficacy of single-dose albendazole against Trichuris trichiura. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Moser, W.; Schindler, C.; Keiser, J. Efficacy of recommended drugs against soil transmitted helminths: Systematic review and network meta-analysis. BMJ 2017, 358, j4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, C.; Wangchuk, P.; Loukas, A. Of dogs and hookworms: Man’s best friend and his parasites as a model for translational biomedical research. Parasit. Vectors 2018, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L.; Miguélez, S.; Romasanta, Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Analysis of the effect of soil saprophytic fungi on the eggs of Baylisascaris procyonis. Parasitol. Res. 2015, 114, 2443–2450. [Google Scholar] [CrossRef]
- Cruz, L.M.; Allanson, M.; Kwa, B.; Azizan, A.; Izurieta, R. Morphological changes of Ascaris spp. eggs during their development outside the host. J. Parasitol. 2012, 98, 63–68. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Treatment | Ascaris suum | Lemurostrongylus sp. | ||
|---|---|---|---|---|---|
| EPG | RR | EPG | RR | ||
| 0 | Control | 563 ± 256 | 490 ± 188 | ||
| CR | 625 ± 290 | 750 ± 177 | |||
| TA | 525 ± 155 | 670 ± 303 | |||
| 1 | Control | 475 ± 240 | (84%) | 400 ± 187 | (81%) |
| CR | 475 ± 290 | (76%) | 530 ± 164 | (70%) | |
| TA | 363 ± 95 | (69%) | 440 ± 207 | (65%) | |
| 2 | Control | 413 ± 229 | (73%) | 325 ± 155 | (67%) |
| CR | 350 ± 178 | (56%) | 420 ± 196 | (54%) | |
| TA | 275 ± 65 | (52%) | 340 ± 167 | (50%) | |
| 3 | Control | 313 ± 131 | (56%) | ||
| CR | 313 ± 189 | (50%) | 320 ± 164 | (41%) | |
| TA | 225 ± 50 | (43%) | 270 ± 84 | (43%) | |
| 4 | Control | 288 ± 193 | (51%) | ||
| CR | 263 ± 131 | (42%) | |||
| TA | 200 ± 41 | (38%) | |||
| Week | Treatment | Viable | Non-Viable | |||
|---|---|---|---|---|---|---|
| Zygote | DE | L1 | L2 | |||
| 0 | Control | 127 ± 11 (100%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 ± 1 |
| CR | 121 ± 9 (100%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 ± 1 | |
| TA | 119 ± 3 (100%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 ± 1 | |
| 1 | Control | 34 ± 3 (29%) | 85 ± 13 (71%) | 0 (0%) | 0 (0%) | 6 ± 3 |
| CR | 33 ± 3 (37%) | 55 ± 9 (63%) | 0 (0%) | 0 (0%) | 37 ± 8 | |
| TA | 52 ± 5 (51%) | 49 ± 4 (49%) | 0 (0%) | 0 (0%) | 35 ± 5 | |
| 2 | Control | 7 ± 10 (7%) | 24 ± 4 (22%) | 77 ± 5 (70%) | 1 ± 2 (1%) | 12 ± 7 |
| CR | 15 ± 4 (21%) | 21 ± 6 (29%) | 35 ± 8 (48%) | 1 ± 2 (2%) | 58 ± 24 | |
| TA | 32 ± 5 (39%) | 34 ± 16 (40%) | 16 ± 1 (20%) | 1 ± 1 (1%) | 56 ± 18 | |
| 3 | Control | 2 ± 1 (2%) | 11 ± 3 (10%) | 32 ± 3 (29%) | 66 ± 5 (59%) | 25 ± 10 |
| CR | 7 ± 4 (13%) | 11 ± 4 (20%) | 19 ± 5 (36%) | 17 ± 5 (31%) | 68 ± 13 | |
| TA | 13 ± 3 (27%) | 14 ± 6 (28%) | 12 ± 3 (24%) | 10 ± 2 (21%) | 84 ± 7 | |
| 4 | Control | 1 ± 1 (0.5%) | 3 ± 2 (2%) | 13 ± 5 (12.5%) | 88 ± 4 (85%) | 28 ± 6 |
| CR | 4 ± 1 (10) (10%) | 3 ± 2 (6%) | 10 ± 6 (25%) | 24 ± 2 (59%) | 77 ± 17 | |
| TA | 4 ± 2 (12%) | 6 ± 1 (19%) | 10 ± 2 (30%) | 13 ± 1 (39%) | 84 ± 16 | |
| Week | Treatment | Viable | Non-Viable | ||
|---|---|---|---|---|---|
| Zygote | DE | L1 | |||
| 0 | Control | 142 ± 10 (91%) | 9 ± 4 (6%) | 5 ± 1 (3%) | 2 ± 1 |
| CR | 142 ± 10 (91%) | 9 ± 4 (6%) | 5 ± 1 (3%) | 2 ± 1 | |
| TA | 142 ± 10 (91%) | 9 ± 4 (6%) | 5 ± 1 (3%) | 1 ± 1 | |
| 1 | Control | 9 ± 2 (6%) | 27 ± 6 (17%) | 121 ± 17 (77%) | 11 ± 6 |
| CR | 38 ± 10 (26%) | 43 ± 11 (29%) | 66 ± 27 (45%) | 18 ± 9 | |
| TA | 60 ± 15 (40%) | 50 ± 10 (34%) | 39 ± 9 (26%) | 17 ± 7 | |
| 2 | Control | 4 ± 1 (3%) | 11 ± 2 (7%) | 136 ± 8 (90%) | 17 ± 5 |
| CR | 23 ± 4 (20%) | 29 ± 6 (24%) | 67 ± 13 (56%) | 48 ± 8 | |
| TA | 28 ± 2 (26) | 25 ± 8 (23%) | 56 ± 6 (51%) | 46 ± 7 | |
| 3 | Control | 24 ± 9 | |||
| CR | 13 ± 3 (13%) | 16 ± 8 (16%) | 70 ± 5 (71%) | 52 ± 8 | |
| TA | 17 ± 3 (17%) | 26 ± 7 (26%) | 58 ± 10 (57%) | 60 ± 5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viña, C.; Silva, M.I.; Palomero, A.M.; Voinot, M.; Vilá, M.; Hernández, J.Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Cazapal-Monteiro, C.F.; Arias, M.S. The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens 2020, 9, 1071. https://doi.org/10.3390/pathogens9121071
Viña C, Silva MI, Palomero AM, Voinot M, Vilá M, Hernández JÁ, Paz-Silva A, Sánchez-Andrade R, Cazapal-Monteiro CF, Arias MS. The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens. 2020; 9(12):1071. https://doi.org/10.3390/pathogens9121071
Chicago/Turabian StyleViña, Cándido, María Isabel Silva, Antonio Miguel Palomero, Mathilde Voinot, María Vilá, José Ángel Hernández, Adolfo Paz-Silva, Rita Sánchez-Andrade, Cristiana Filipa Cazapal-Monteiro, and María Sol Arias. 2020. "The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi" Pathogens 9, no. 12: 1071. https://doi.org/10.3390/pathogens9121071