Curcumin Attenuates the Pathogenicity of Entamoeba histolytica by Regulating the Expression of Virulence Factors in an Ex-Vivo Model Infection

 ,
,
Abstract
:1. Introduction
2. Results
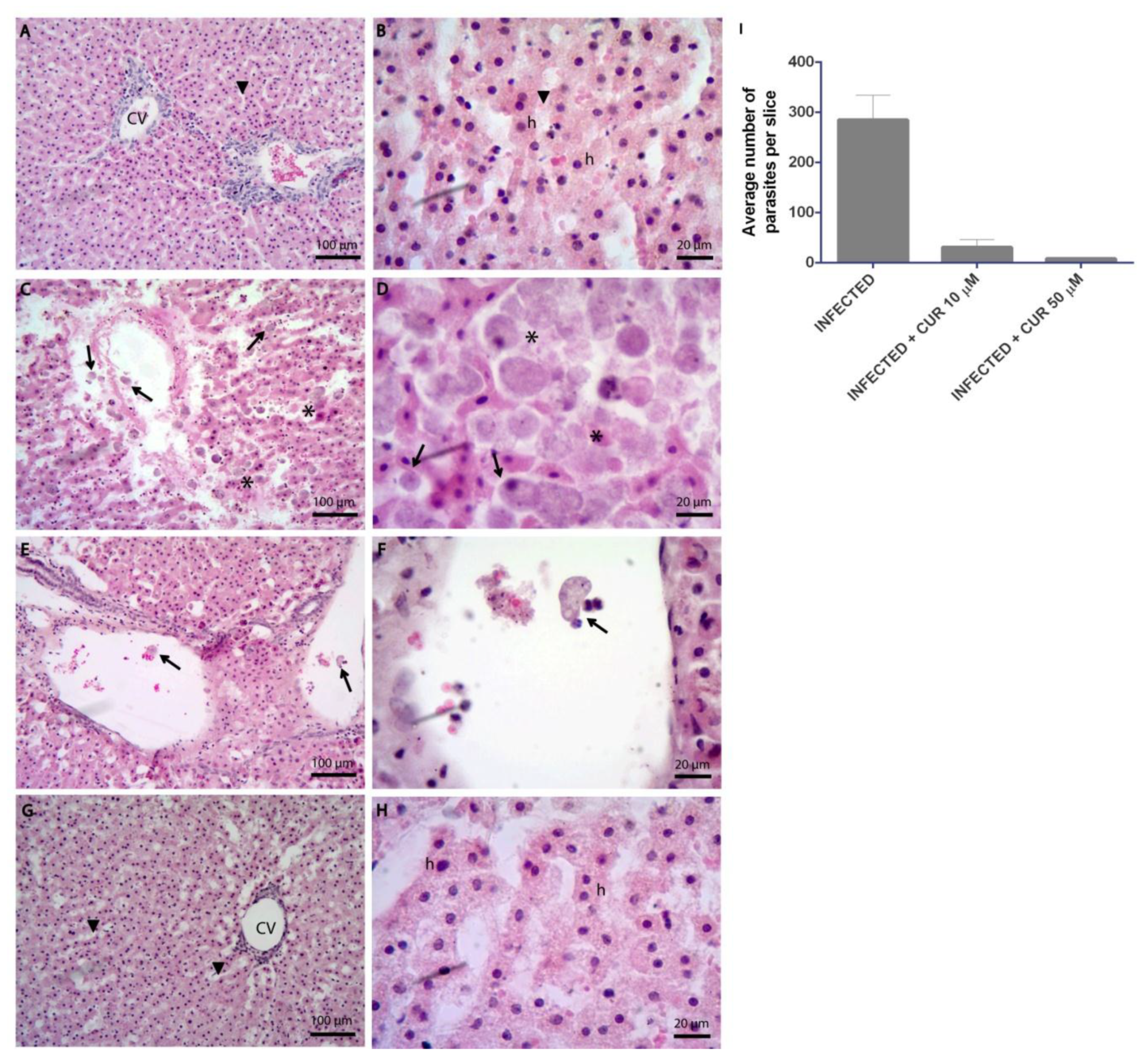
2.1. Curcumin Prevents the Tissue Damage Caused by Entamoeba histolytica
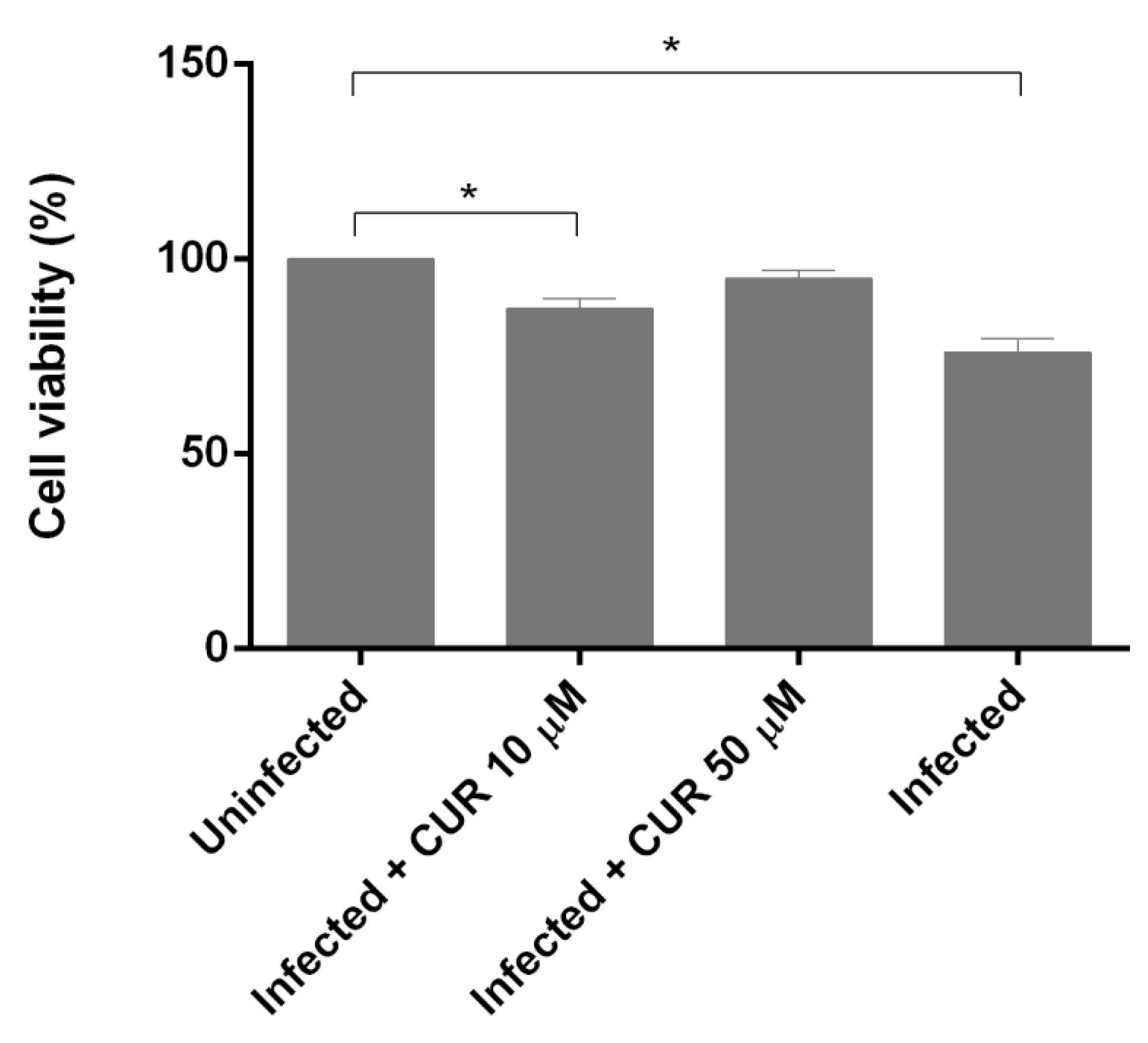
2.2. Effect of Curcumin on PCHLS Viability
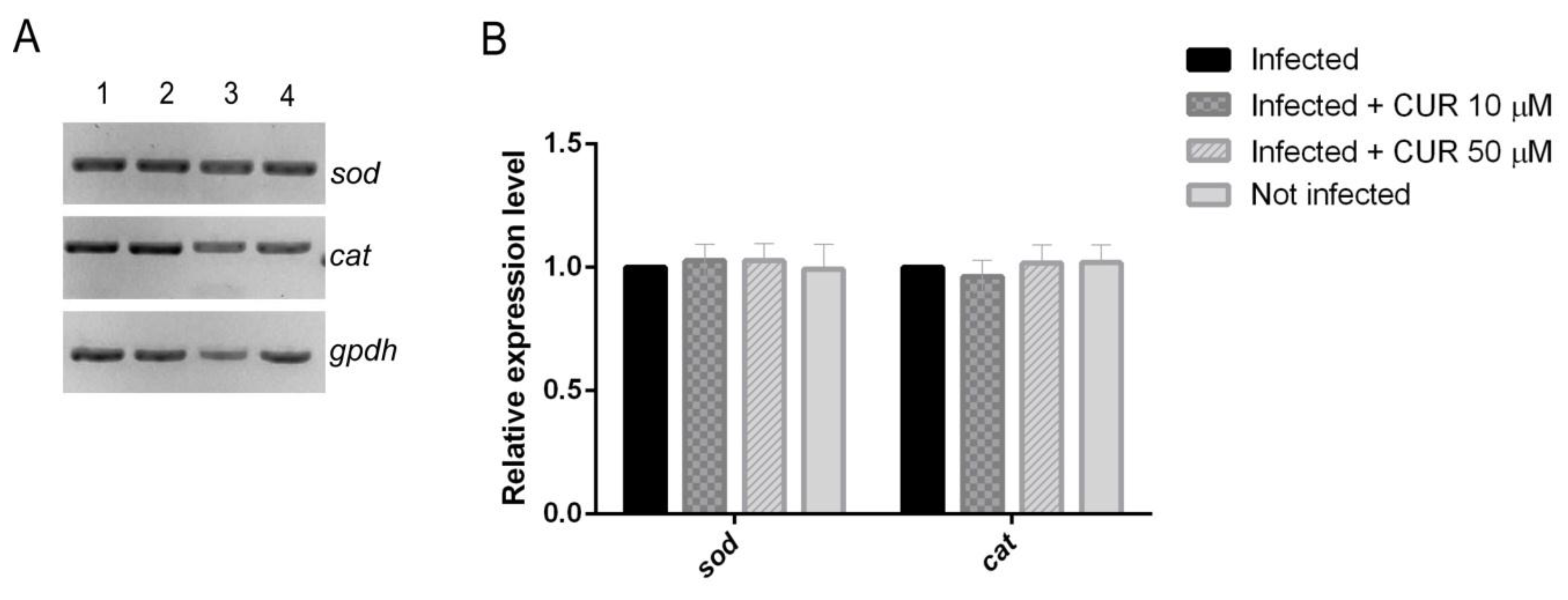
2.3. Effect of Curcumin on Antioxidant Enzymes Expression
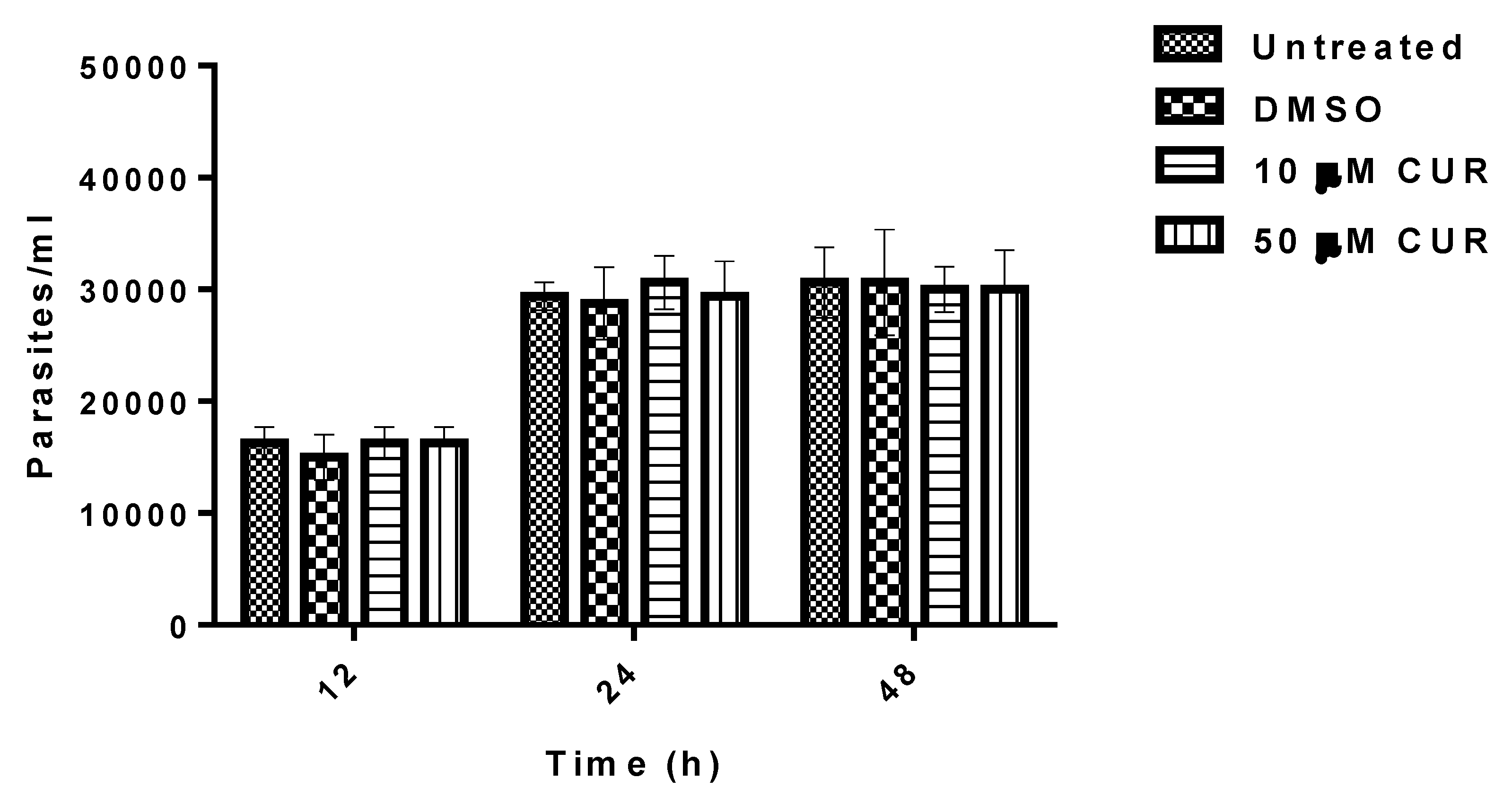
2.4. Effect of Curcumin on the Growth of Entamoeba histolytica
2.5. Curcumin Regulates the Expression of Virulence Factors
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Entamoeba histolytica Trophozoites Culture
4.3. Precision-Cut Hamster Liver Slices Preparation
4.4. Preparation of Curcumin Stocks
4.5. Ex Vivo Infection
4.6. Histopathological Image Analysis
4.7. Scanning Electron Microscopy
4.8. Viability Assay
4.9. Expression of Virulence Factors and Antioxidant Enzymes in PCHLS
4.10. Growth Inhibition Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Espinosa-Cantellano, M.; Martinez-Palomo, A. Pathogenesis of intestinal amebiasis: From molecules to disease. Clin. Microbiol. Rev. 2000, 13, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Salles, J.M.; Moraes, L.A.; Salles, M.C. Hepatic amebiasis. Braz. J. Infect. Dis. 2003, 7, 96–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.; Solaymani-Mohammadi, S.; Parker, S.K. Hahram Hepatic amebiasis: A reminder of the complications. Curr. Opin. Pediatr. 2009, 21, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Moncada, D.; Keller, K.; Chadee, K. Entamoeba histolytica cysteine proteinases disrupt the polymeric structure of colonic mucin and alter its protective function. Infect. Immun. 2003, 71, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, V.; Serrano-Luna, J.; Ramírez-Moreno, E.; Tsutsumi, V.; Shibayama, M. Entamoeba histolytica: Overexpression of the gal/galnac lectin, ehcp2 and ehcp5 genes in an in vivo model of amebiasis. Parasitol. Int. 2016, 65, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Lidell, M.E.; Moncada, D.M.; Chadee, K.; Hansson, G. Entamoeba histolytica cysteine proteases cleave the MUC2 mucin in its C-terminal domain and dissolve the protective colonic mucus gel. Proc. Natl. Acad. Sci. USA 2006, 103, 9298–9303. [Google Scholar] [CrossRef] [PubMed]
- Cornick, S.; Moreau, F.; Chadee, K. Entamoeba histolytica Cysteine Proteinase 5 Evokes Mucin Exocytosis from Colonic Goblet Cells via alphavbeta3 Integrin. PLoS Pathog. 2016, 12, 1005579. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Z.; Alexander, D.; Bracha, R.; Mirelman, D.; Stanley, S.L. Expression of amoebapores is required for full expression of Entamoeba histolytica virulence in amebic liver abscess but is not necessary for the induction of inflammation or tissue damage in amebic colitis. Infect. Immun. 2004, 72, 678–683. [Google Scholar] [CrossRef]
- Melendez-Lopez, S.G. Use of recombinant Entamoeba histolytica cysteine proteinase 1 to identify a potent inhibitor of amebic invasion in a human colonic model. Eukaryot. Cell 2007, 6, 1130–1136. [Google Scholar] [CrossRef]
- Tsutsumi, V.; Shibayama, M. Experimental amebiasis: A selected review of some in vivo models. Arch. Med. Res. 2006, 37, 210–220. [Google Scholar] [CrossRef]
- Bansal, D.; Ave, P.; Kerneis, S.; Frileux, P.; Boché, O.; Baglin, A.C.; Dubost, G.; Leguern, A.S. An ex-vivo human intestinal model to study Entamoeba histolytica pathogenesis. PLoS Negl. Trop. Dis. 2009, 3, e551. [Google Scholar] [CrossRef] [PubMed]
- Ximenez, C. Differential expression of pathogenic genes of Entamoeba histolytica vs E. dispar in a model of infection using human liver tissue explants. PLoS ONE 2017, 12, e0181962. [Google Scholar]
- Lasserre, R. Single-day drug treatment of amebic liver abscess. Am. J. Trop. Med. Hyg. 1983, 32, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F. Nitazoxanide in the treatment of amoebiasis. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Haque, R. Amebiasis. N. Engl. J. Med. 2003, 348, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.K.; Abramovitch, R.B. Small Molecules That Sabotage Bacterial Virulence. Trends Pharmacol. Sci. 2017, 38, 339–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.C. Targeting virulence: Can we make evolution-proof drugs? Nat. Rev. Microbiol. 2014, 12, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Pavan, A.R.; Silva, G.D.; Jornada, D.H.; Chiba, D.E. Unraveling the Anticancer Effect of Curcumin and Resveratrol. Nutrients 2016, 8, 628. [Google Scholar] [CrossRef]
- Gupta, S.C. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef]
- Wachter, B. In vitro efficacy of curcumin on Trichomonas vaginalis. Wien. Klin. Wochenschr. 2014, 126, S32–S36. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, D. Curcumin Generates Oxidative Stress and Induces Apoptosis in Adult Schistosoma mansoni Worms. PLoS ONE 2016, 11, e0167135. [Google Scholar]
- Gutierrez-Gutierrez, F.; Palomo-Ligas, L.; Hernández-Hernández, J.M. Curcumin alters the cytoskeleton and microtubule organization on trophozoites of Giardia lamblia. Acta Trop. 2017, 172, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Qiu, M.; He, D.; Zhang, R.; Tao, P.; Liang, Y.; Yang, Y.; Guo, Y.; Ye, X. Protective effect of curcumin on hepatocytes in rats with sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2017, 29, 162–166. [Google Scholar] [PubMed]
- Gressler, L.T. Trypanocidal activity of free and nanoencapsulated curcumin on Trypanosoma evansi. Parasitology 2015, 142, 439–448. [Google Scholar] [CrossRef]
- Bao, W. Curcumin alleviates ethanol-induced hepatocytes oxidative damage involving heme oxygenase-1 induction. J. Ethnopharmacol. 2010, 128, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Naik, R.S.; Mujumdar, A.M.; Ghaskadbi, S. Protection of liver cells from ethanol cytotoxicity by curcumin in liver slice culture in vitro. J. Ethnopharmacol. 2004, 95, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Castaneda, I.A.; Hernández-Hernández, J.M.; Pérez-Rangel, A.; González-Pozos, S.; Carranza-Rosales, P.; Charles-Niño, C.L. Amoebicidal activity of curcumin on Entamoeba histolytica trophozoites. J. Pharm. Pharmacol. 2018, 70, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Aldaba-Muruato, L.R. Adrenergic regulation during acute hepatic infection with Entamoeba histolytica in the hamster: Involvement of oxidative stress, Nrf2 and NF-KappaB. Parasite 2017, 24, 46. [Google Scholar] [CrossRef]
- Han, W. Curcumin alleviated liver oxidative stress injury of rat induced by paraquat. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi 2014, 32, 352–356. [Google Scholar]
- El-Bahr, S.M. Effect of curcumin on hepatic antioxidant enzymes activities and gene expressions in rats intoxicated with aflatoxin B1. Phytother. Res. 2015, 29, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, Z.; Díaz-Godínez, C.; Mora, N.; Alemán, O.R.; Uribe-Querol, E.; Carrero, J.C.; Rosales, C. Entamoeba histolytica Induce Signaling via Raf/MEK/ERK for Neutrophil Extracellular Trap (NET) Formation. Front. Cell Infect. Microbiol. 2018, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Nakada-Tsukui, K.; Sekizuka, T.; Sato-Ebine, E.; Cadiz, A. AIG1 affects in vitro and in vivo virulence in clinical isolates of Entamoeba histolytica. PLoS Pathog. 2018, 14, e1006882. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Zobeiri, M.; Parvizi, F.; El-Senduny, F.F.; Marmouzi, I.; Coy-Barrera, E. Curcumin in Liver Diseases: A Systematic Review of the Cellular Mechanisms of Oxidative Stress and Clinical Perspective. Nutrients 2018, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Carranza-Rosales, P.; Santiago-Mauricio, M.G.; Guzmán-Delgado, N.E.; Vargas-Villarreal, J.; Lozano-Garza, G. Precision-cut hamster liver slices as an ex vivo model to study amoebic liver abscess. Exp. Parasitol. 2010, 126, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Carranza-Rosales, P. Induction of virulence factors, apoptosis, and cytokines in precision-cut hamster liver slices infected with Entamoeba histolytica. Exp. Parasitol. 2012, 132, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Girish, C.; Pradhan, S.C. Hepatoprotective activities of picroliv, curcumin, and ellagic acid compared to silymarin on carbon-tetrachloride-induced liver toxicity in mice. J. Pharmacol. Pharmacother. 2012, 3, 149–155. [Google Scholar]
- Stanley, S.L. Role of the Entamoeba histolytica cysteine proteinase in amebic liver abscess formation in severe combined immunodeficient mice. Infect. Immun. 1995, 63, 1587–1590. [Google Scholar] [Green Version]
- Ankri, S. Antisense inhibition of expression of cysteine proteinases affects Entamoeba histolytica-induced formation of liver abscess in hamsters. Infect. Immun. 1999, 67, 421–422. [Google Scholar]
- Rudrappa, T.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J. Agric. Food Chem. 2008, 56, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Packiavathy, I.A. Prevention of quorum-sensing-mediated biofilm development and virulence factors production in Vibrio spp. by curcumin. Appl. Microbiol. Biotechnol. 2013, 97, 10177–10187. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, X.; Lin, H. Curcumin as a Promising Antibacterial Agent: Effects on Metabolism and Biofilm Formation in S. mutans. Biomed. Res. Int. 2018, 2018, 4508709. [Google Scholar] [CrossRef] [PubMed]
- Said-Fernandez, S. PEHPS medium: An alternative for axenic cultivation of Entamoeba histolytica and E. invadens. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 249–253. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S. Cell Viability Assays; National Center for Biotechnology Information: Bethesda, MD, USA, 2004.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gen Bank Accession | Sequence (5ʹ-3ʹ) | Amplicon Size (bp) |
|---|---|---|---|
| sod | XM_005073823 | AGGGCACCATCCACTTCGAGCAG | 224 |
| AGCAGTCACGTTGCCCAGGT | |||
| cat | XM_005064849 | GCAGCTCAGAAATCCTACACCA | 332 |
| CCAATGTGCTCAAACACCTTTGC | |||
| gpdh | U10983 | CCAACGTGTCCGTCGTGGAT | 255 |
| ACCCTGTTGCTGTAGCCGAA | |||
| ehcp 1 | XM_645064 | CATGTAGAAGTGATGTGAAAGC | 247 |
| TTCTTTCCCATCAACAACA | |||
| ehcp 5 | XM_645845 | TCCAGCTATTAGAGACCAAGCATC | 398 |
| TAACTCCAGAAGCATCAATAGC | |||
| amoebapore | M83945 | AAGGAGAAATCCTCTGCAAC | 216 |
| CAAATAGCATTGGCATCAAC | |||
| gal/galnac | XM_651089 | CACTTGTCAAATACACAGCAGGAC | 484 |
| GGTTTAGCTTTAGGCCATGGAA | |||
| actin | XM_001914602 | AATGAAAGATTCAGATGCCC | 283 |
| ATTGATCCTCCAATCCAGAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel-Castañeda, I.A.; Carranza-Rosales, P.; Guzmán-Delgado, N.E.; Hernández-Hernández, J.M.; González-Pozos, S.; Pérez-Rangel, A.; Castillo-Romero, A. Curcumin Attenuates the Pathogenicity of Entamoeba histolytica by Regulating the Expression of Virulence Factors in an Ex-Vivo Model Infection. Pathogens 2019, 8, 127. https://doi.org/10.3390/pathogens8030127
Rangel-Castañeda IA, Carranza-Rosales P, Guzmán-Delgado NE, Hernández-Hernández JM, González-Pozos S, Pérez-Rangel A, Castillo-Romero A. Curcumin Attenuates the Pathogenicity of Entamoeba histolytica by Regulating the Expression of Virulence Factors in an Ex-Vivo Model Infection. Pathogens. 2019; 8(3):127. https://doi.org/10.3390/pathogens8030127
Chicago/Turabian StyleRangel-Castañeda, Itzia Azucena, Pilar Carranza-Rosales, Nancy Elena Guzmán-Delgado, José Manuel Hernández-Hernández, Sirenia González-Pozos, Armando Pérez-Rangel, and Araceli Castillo-Romero. 2019. "Curcumin Attenuates the Pathogenicity of Entamoeba histolytica by Regulating the Expression of Virulence Factors in an Ex-Vivo Model Infection" Pathogens 8, no. 3: 127. https://doi.org/10.3390/pathogens8030127