
Impact of Sub-Inhibitory Concentrations of Amoxicillin on Streptococcus suis Capsule Gene Expression and Inflammatory Potential

Abstract
:1. Introduction
2. Results
2.1. Determination of MIC of Amoxicillin for S. suis
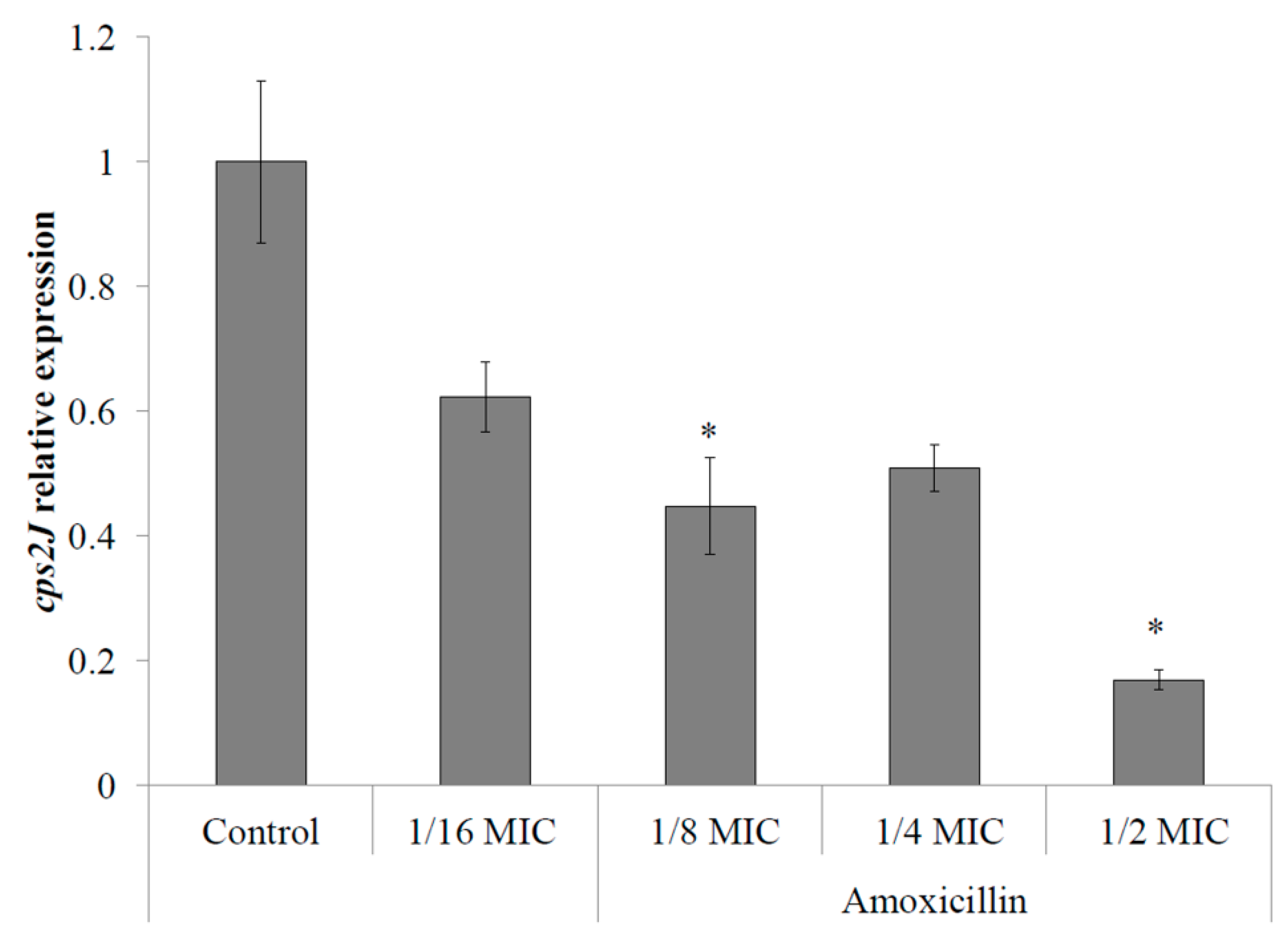
2.2. S. suis Capsule Expression
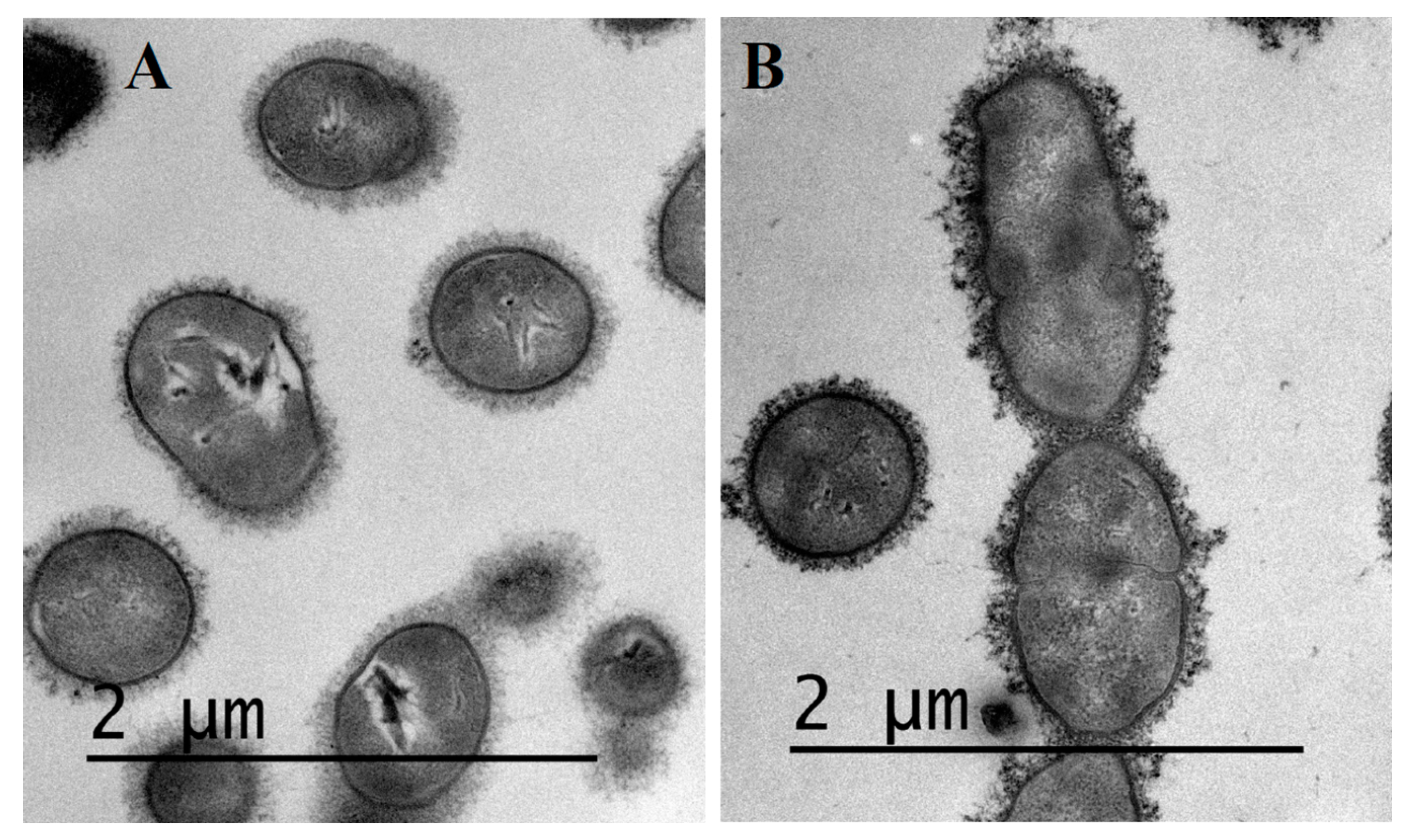
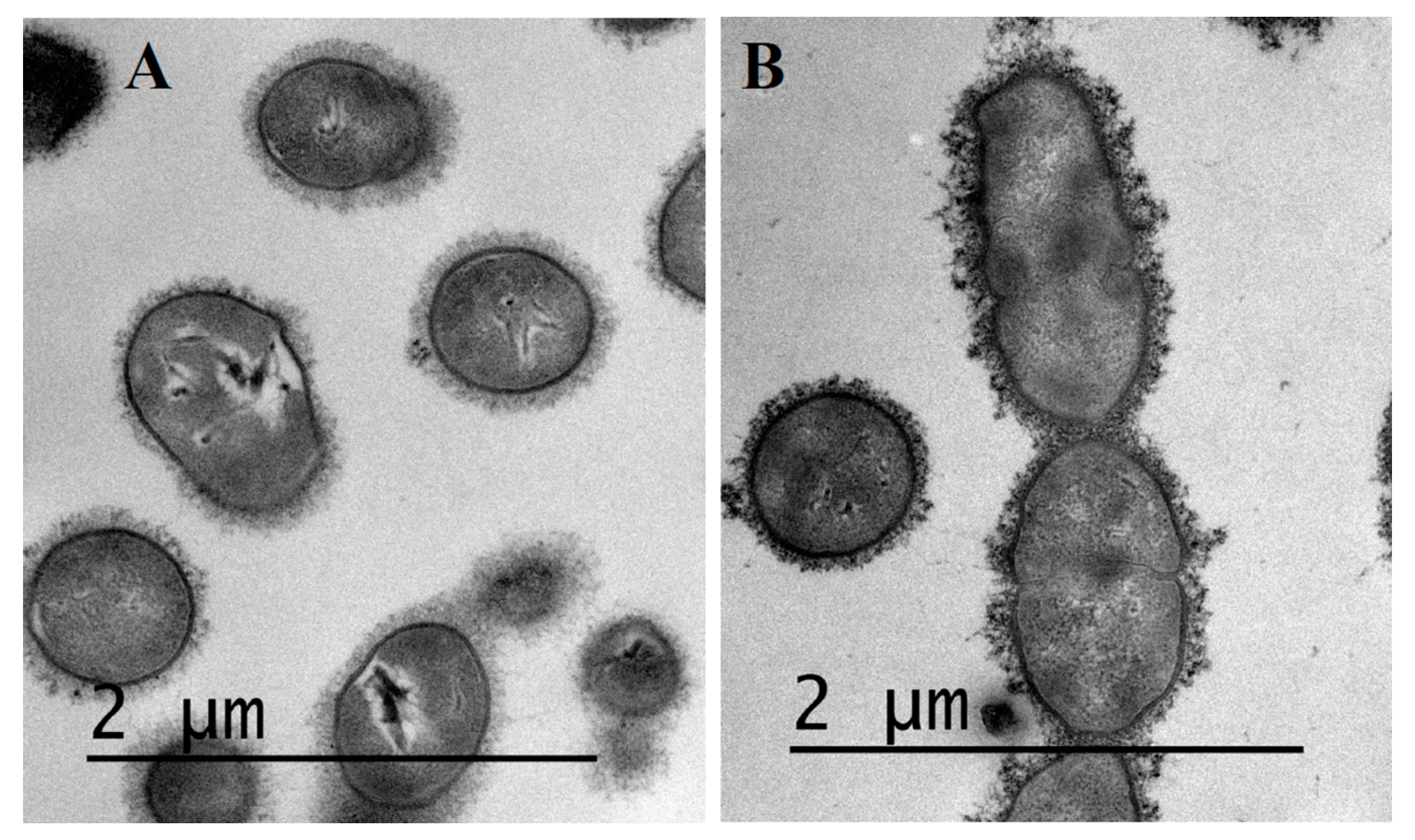
2.3. Visualization of S. suis Capsule by Transmission Electron Microscopy
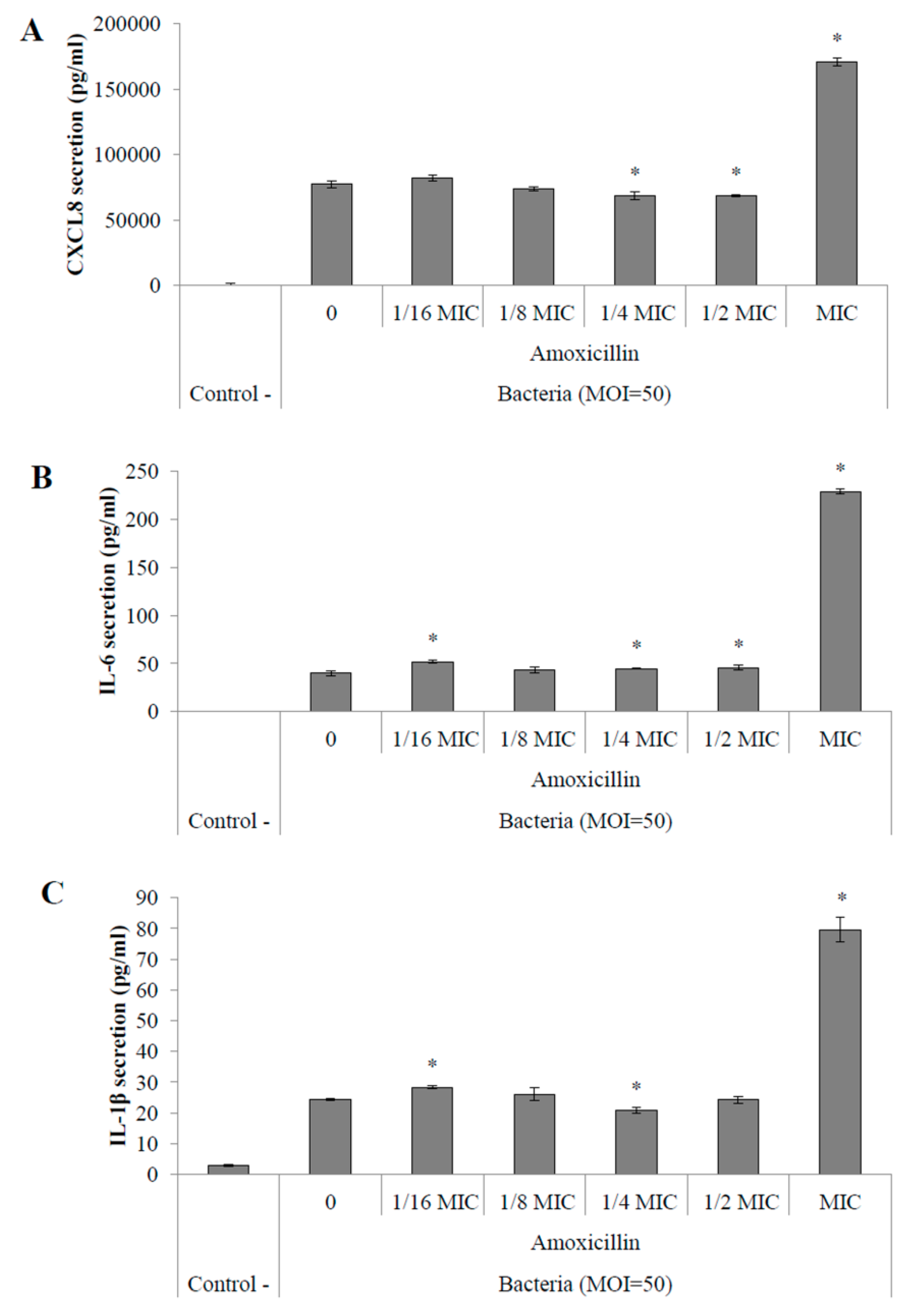
2.4. Pro-Inflammatory Potential of S. suis
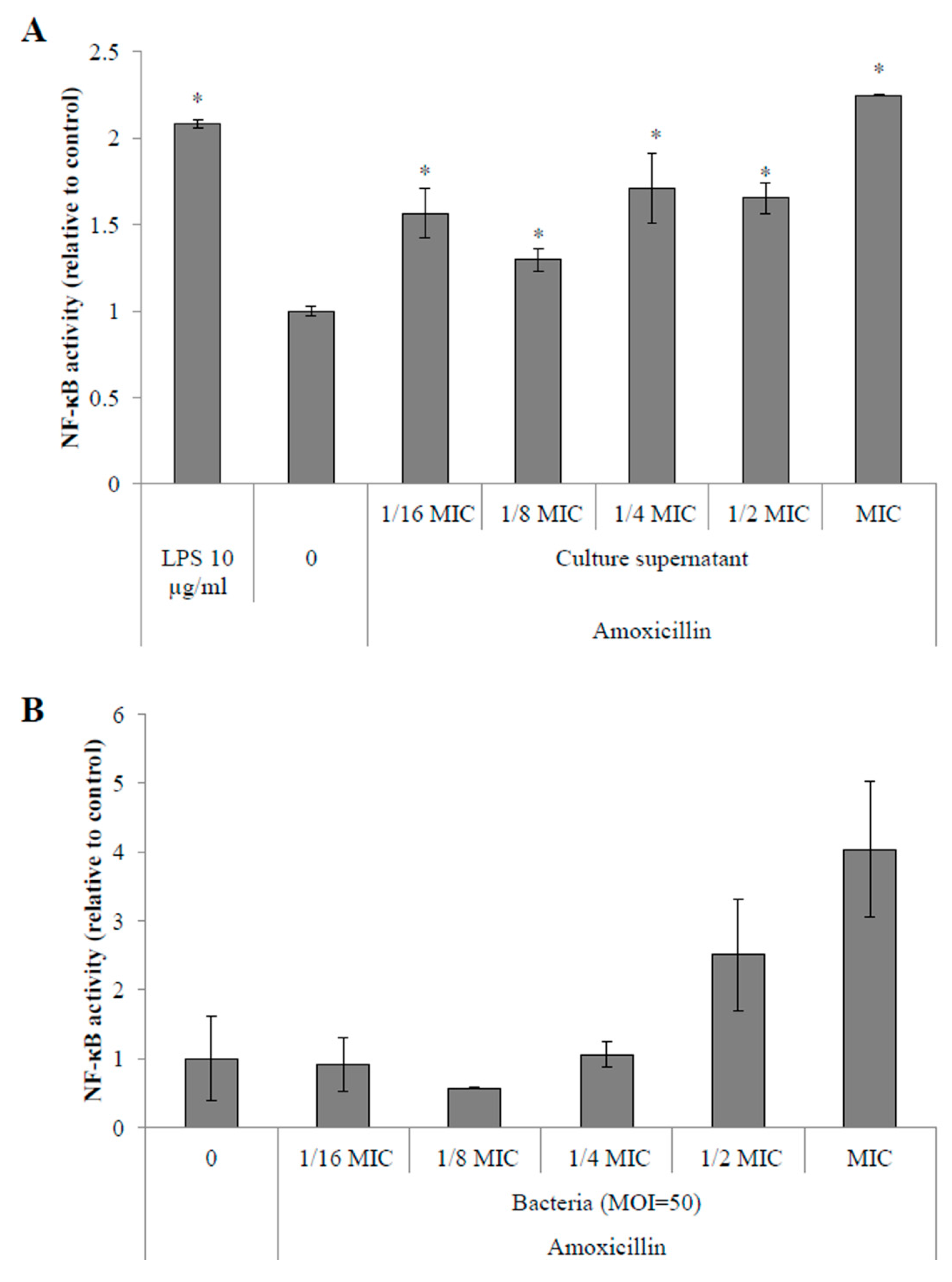
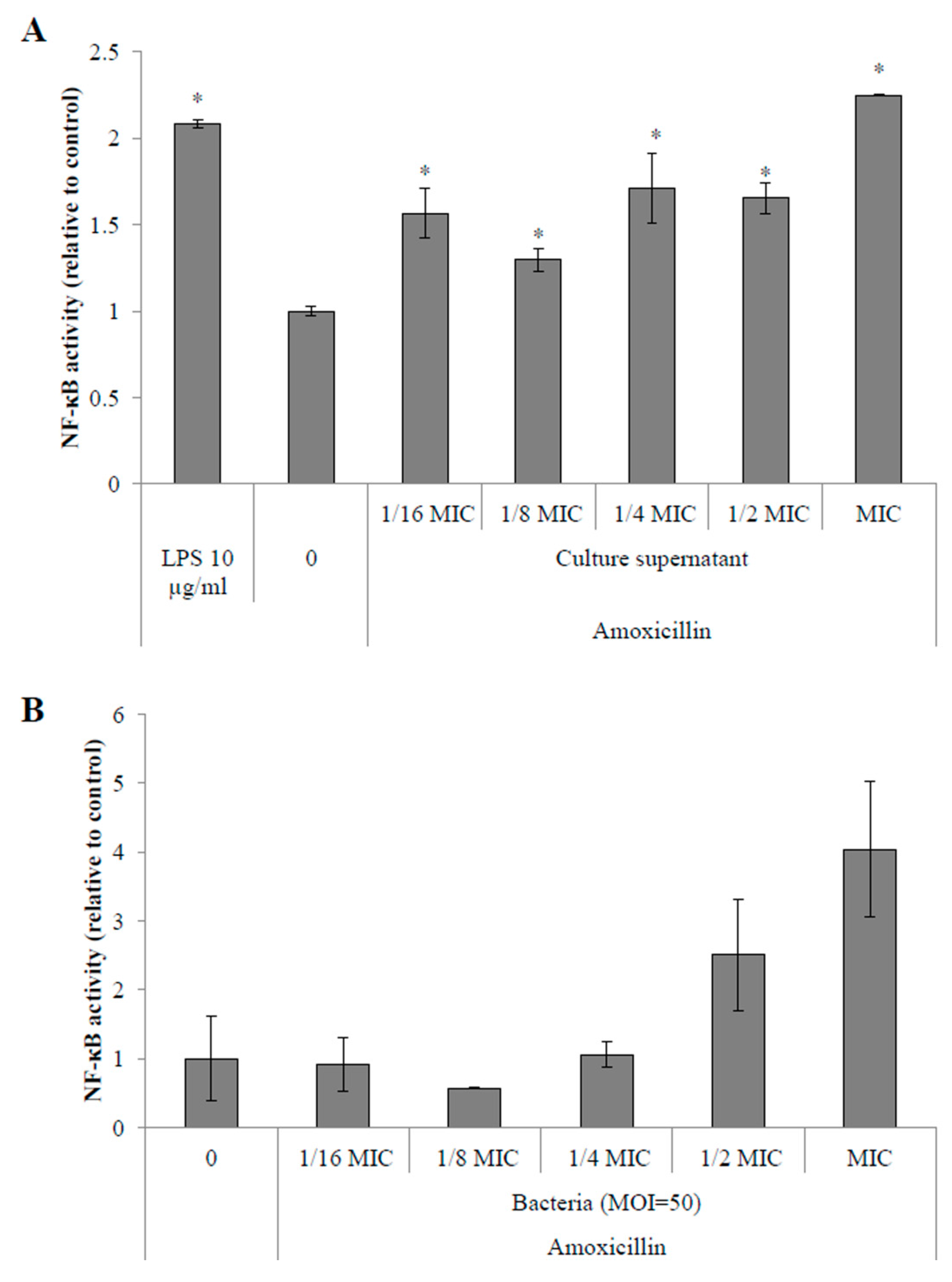
2.4.1. Activation of the NF-κB Pathway in Monocytes
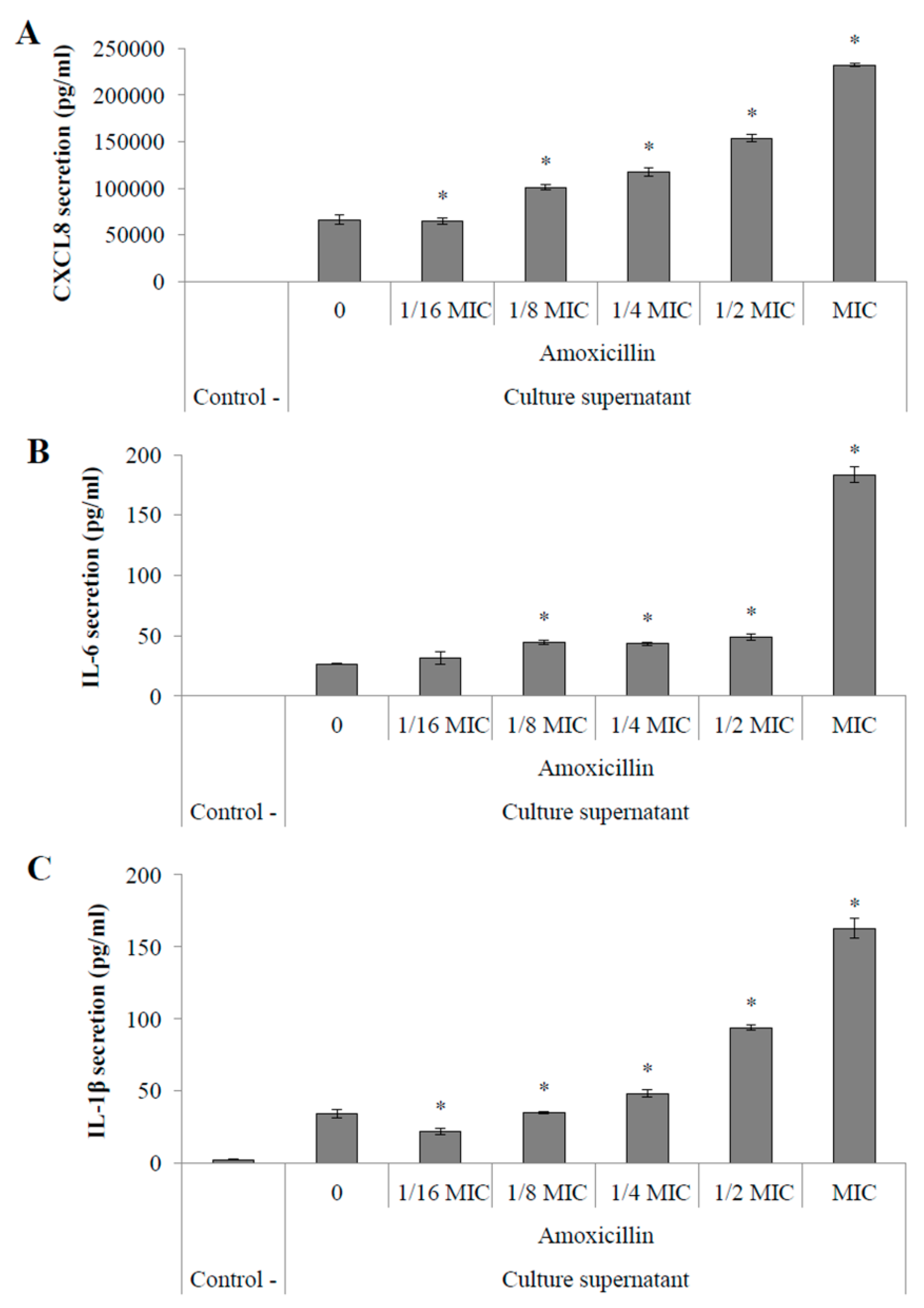
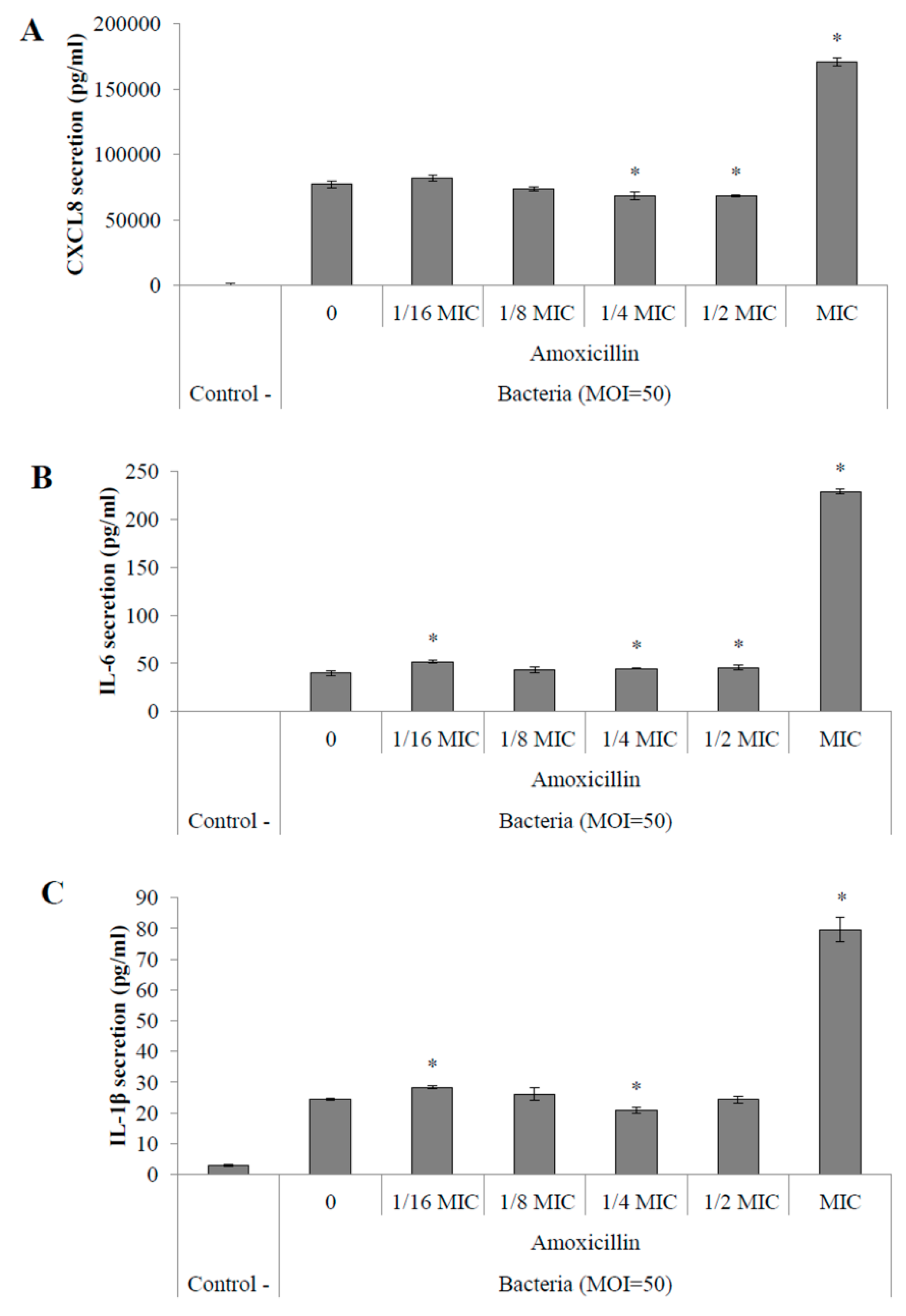
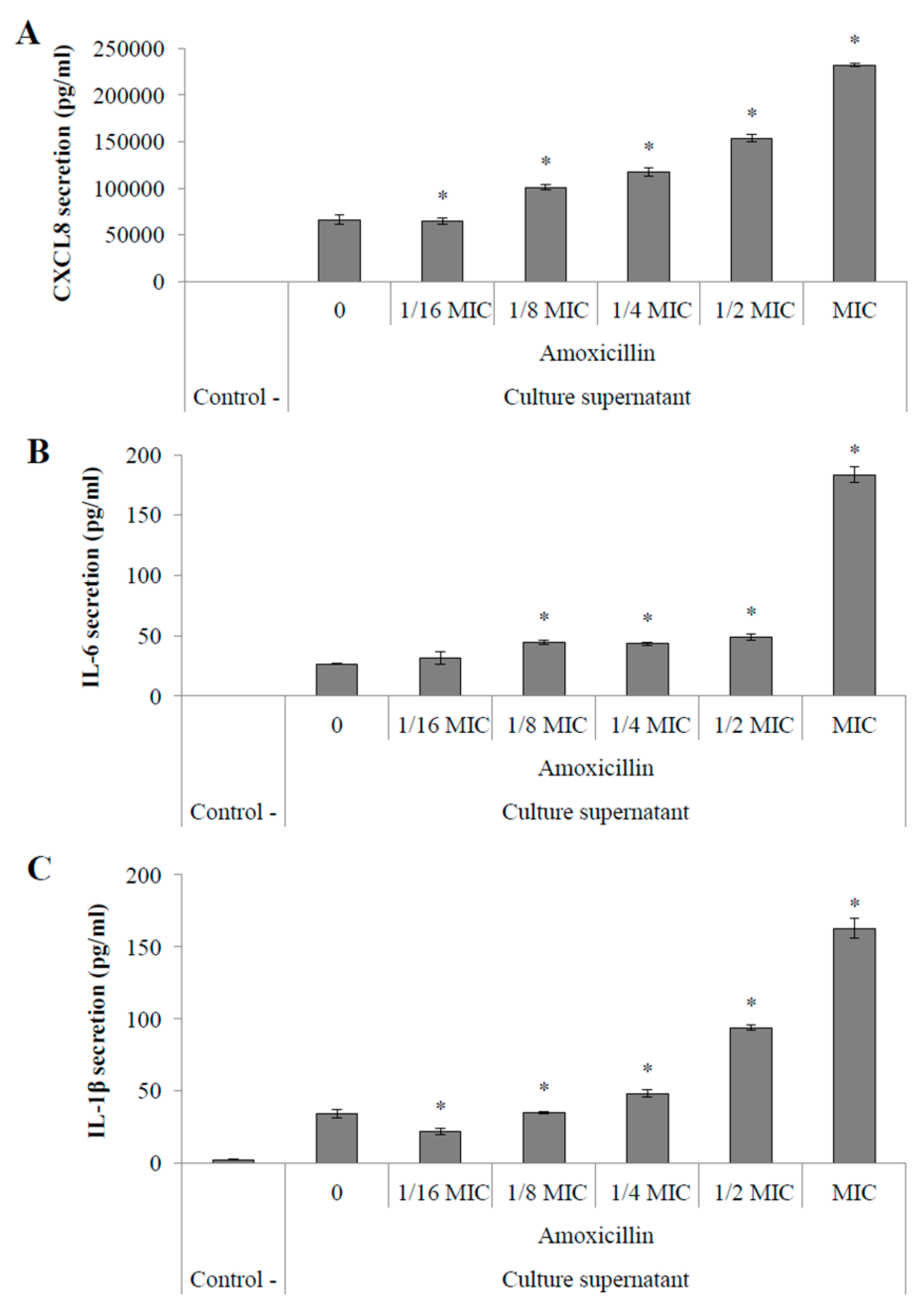
2.4.2. Pro-Inflammatory Cytokine Secretion by Macrophages
3. Discussion
4. Materials and Methods
4.1. Bacteria and Growth Conditions
4.2. Determination of Minimal Inhibitory Concentration (MIC) of Amoxicillin on S. suis
4.3. S. suis Capsule Expression
4.4. Visualization of S. suis Capsule by Transmission Electron Microscopy
4.5. Analysis of the Pro-Inflammatory Potential of S. suis Conditioned with Sub-Inhibitory Concentrations of Amoxicillin
4.5.1. Activation of the NF-κB Pathway in Monocytes
4.5.2. Quantification of Pro-Inflammatory Cytokine Secretion by Macrophages
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| cDNA | complementary deoxyribonucleic acid |
| CXCL8 | interleukin-8 |
| FBS | fetal bovine serum |
| IL | interleukin |
| LPS | lipopolysaccharide |
| MIC | minimal inhibitory concentration |
| MOI | multiplicity of infection |
| NF-κB | nuclear factor-κB |
| PBS | phosphate-buffered saline |
| PCR | polymerase chain reaction |
| PMA | phorbol myristate acetate |
| qRT-PCR | quantitative reverse transcription polymerase chain reaction |
| RNA | ribonucleic acid |
| RPMI-1640 | Roswell Park Memorial Institute medium-1640 |
| THB | Todd-Hewitt broth |
References
- Gottschalk, M. Streptococcosis. In Diseases of Swine; Karriger, L.R.A., Schwartz, K.J., Stevenson, G., Zimmerman, J., Eds.; Wiley Publishers: Hoboken, NJ, USA, 2012; pp. 841–855. [Google Scholar]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16s rdna sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Tien le, H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA-DNA homology and soda and recn phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.L.; Nghia, H.D.T.; Taylor, W.; Schultsz, C. Streptococcus suis: An emerging human pathogen. Clin. Infect. Dis. 2009, 48, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, H.; Wu, Z.; Wang, S.; Cao, M.; Hu, D.; Wang, C. Streptococcus suis infection: An emerging/reemerging challenge of bacterial infectious diseases? Virulence 2014, 5, 477–497. [Google Scholar] [CrossRef] [PubMed]
- Segura, M. Streptococcus suis vaccines: Candidate antigens and progress. Expert Rev. Vaccines 2015, 14, 1587–1608. [Google Scholar] [CrossRef] [PubMed]
- Euzeby, J.P. Streptococcus: A short review. Rev. Med. Vet. Toulouse 1999, 150, 981–988. [Google Scholar]
- Kataoka, Y.; Yoshida, T.; Sawada, T. A 10-year survey of antimicrobial susceptibility of Streptococcus suis isolates from swine in Japan. J. Vet. Med. Sci. 2000, 62, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rotman, E.R.; Shoemaker, N.B.; Salyers, A.A. Translational control of tetracycline resistance and conjugation in the bacteroides conjugative transposon ctndot. J. Bacteriol. 2005, 187, 2673–2680. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Spiegelman, G.B.; Yim, G. The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Braga, P.C.; Sala, M.T.; Dal Sasso, M. Pharmacodynamic effects of subinhibitory concentrations of rufloxacin on bacterial virulence factors. Antimicrob. Agents Chemother. 1999, 43, 1013–1019. [Google Scholar] [PubMed]
- Haddadin, R.N.; Saleh, S.; Al-Adham, I.S.; Buultjens, T.E.; Collier, P.J. The effect of subminimal inhibitory concentrations of antibiotics on virulence factors expressed by Staphylococcus aureus biofilms. J. Appl. Microbiol. 2010, 108, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Doss, S.A.; Tillotson, G.S.; Amyes, S.G. Effect of sub-inhibitory concentrations of antibiotics on the virulence of Staphylococcus aureus. J. Appl. Bacteriol. 1993, 75, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.J.; Keyser, R. Effects of subinhibitory concentrations of macrolide antibiotics on Pseudomonas aeruginosa. Chest 2004, 125, 62S–69S, quiz 69S. [Google Scholar] [CrossRef] [PubMed]
- Subrt, N.; Mesak, L.R.; Davies, J. Modulation of virulence gene expression by cell wall active antibiotics in Staphylococcus aureus. J. Antimicrob. Chemother. 2011, 66, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Shi, Y.; Zhang, D.; Wei, J.; Surette, M.G.; Duan, K. Modulation of secreted virulence factor genes by subinhibitory concentrations of antibiotics in Pseudomonas aeruginosa. J. Microbiol. 2008, 46, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [PubMed]
- Brook, I.; Hausfeld, J.N. In vitro effects of penicillin and telithromycin on the expression of Streptococcus pneumoniae capsule. J. Antimicrob. Chemother. 2006, 58, 678–679. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Rasmussen, S.R.; Artursson, K.; Jensen, N.E. Trends in the resistance to antimicrobial agents of Streptococcus suis isolates from Denmark and Sweden. Vet. Microbiol. 1998, 63, 71–80. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Zhou, Y.H.; Chen, J.Q.; Huang, Q.Y.; Han, Q.; Liu, B.; Cheng, G.D.; Li, Y.H. Quantitative proteomic analysis of sub-mic erythromycin inhibiting biofilm formation of S. suis in vitro. J. Proteom. 2015, 116, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.E.; Damman, M.; van der Velde, J.; Wagenaar, F.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Smits, M.A. Identification and characterization of the cps locus of Streptococcus suis serotype 2: The capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 1999, 67, 1750–1756. [Google Scholar] [PubMed]
- Tanabe, S.I.; Bonifait, L.; Fittipaldi, N.; Grignon, L.; Gottschalk, M.; Grenier, D. Pleiotropic effects of polysaccharide capsule loss on selected biological properties of Streptococcus suis. Can. J. Vet. Res. 2010, 74, 65–70. [Google Scholar] [PubMed]
- Lakkitjaroen, N.; Takamatsu, D.; Okura, M.; Sato, M.; Osaki, M.; Sekizaki, T. Capsule loss or death: The position of mutations among capsule genes sways the destiny of Streptococcus suis. Fems Microbiol. Lett. 2014, 354, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.E.; Veenbergen, V.; van der Velde, J.; Damman, M.; Wisselink, H.J.; Smits, M.A. The cps genes of Streptococcus suis serotypes 1, 2, and 9: Development of rapid serotype-specific pcr assays. J. Clin. Microbiol. 1999, 37, 3146–3152. [Google Scholar] [PubMed]
- Segura, M.; Gottschalk, M.; Olivier, M. Encapsulated Streptococcus suis inhibits activation of signaling pathways involved in phagocytosis. Infect. Immun. 2004, 72, 5322–5330. [Google Scholar] [CrossRef] [PubMed]
- Houde, M.; Gottschalk, M.; Gagnon, F.; Van Calsteren, M.R.; Segura, M. Streptococcus suis capsular polysaccharide inhibits phagocytosis through destabilization of lipid microdomains and prevents lactosylceramide-dependent recognition. Infect. Immun. 2012, 80, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Charland, N.; Harel, J.; Kobisch, M.; Lacasse, S.; Gottschalk, M. Streptococcus suis serotype 2 mutants deficient in capsular expression. Microbiology 1998, 144, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Cao, M.; Shi, J.; Zhang, H.; Hu, D.; Zhu, J.; Zhang, X.; Geng, M.; Zheng, F.; Pan, X.; et al. Attenuation of Streptococcus suis virulence by the alteration of bacterial surface architecture. Sci. Rep. 2012, 2, 710. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Goethe, R.; Rohde, M.; Valentin-Weigand, P. Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cell. Microbiol. 2004, 6, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Lakkitjaroen, N.; Takamatsu, D.; Okura, M.; Sato, M.; Osaki, M.; Sekizaki, T. Loss of capsule among Streptococcus suis isolates from porcine endocarditis and its biological significance. J. Med. Microbiol. 2011, 60, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Raithatha, A.H.; Bryden, D.C. Use of intravenous immunoglobulin therapy in the treatment of septic shock, in particular severe invasive group a streptococcal disease. Indian J. Crit. Care Med. 2012, 16, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, T.S.; Christman, J.W. The role of nuclear factor-kappa B in cytokine gene regulation. Am. J. Resp. Cell Mol. Biol. 1997, 17, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.J.; Pridmore, A.C.; Lee, M.E.; Read, R.C. Induction of pro-inflammatory cytokine release by human macrophages during exposure of Streptococcus pneumoniae to penicillin is influenced by minimum inhibitory concentration ratio. Int. J. Antimicrob. Agents 2005, 26, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.Y.; Zheng, H.; Zhang, J.; Jing, H.Q.; Wang, L.; Xiong, Y.W.; Wang, W.; Zhou, Z.M.; Sun, Q.Z.; Luo, X.; et al. Clinical, experimental, and genomic differences between intermediately pathogenic, highly pathogenic, and epidemic Streptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Stankova, J.; Gottschalk, M. Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect. Immun. 1999, 67, 4646–4654. [Google Scholar] [PubMed]
- Zheng, H.; Punaro, M.C.D.; Segura, M.; Lachance, C.; Rivest, S.; Xu, J.G.; Houde, M.; Gottschalk, M. Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J. Neuroimmunol. 2011, 234, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Luo, X.; Segura, M.; Sun, H.; Ye, C.; Gottschalk, M.; Xu, J. The role of toll-like receptors in the pathogenesis of Streptococcus suis. Vet. Microbiol. 2011, 156, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Phillips, I.; Casewell, M.; Cox, T.; de Groot, B.; Friis, C.; Jones, R.; Nightingale, C.; Preston, R.; Waddell, J. Does the use of antibiotics in food animals pose a risk to human health? A critical review of published data. J. Antimicrob. Chemother. 2004, 53, 28–52. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Liu, M.; Jin, M.; Liu, J.; Chen, Y.; Wu, T.; Fu, T.; Bei, W.; Chen, H. The key virulence-associated genes of Streptococcus suis type 2 are upregulated and differentially expressed in vivo. Fems Microbiol. Lett. 2008, 278, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Vanrobaeys, M.; De Herdt, P.; Charlier, G.; Ducatelle, R.; Haesebrouck, F. Ultrastructure of surface components of Streptococcus gallolytics (S. bovis) strains of differing virulence isolated from pigeons. Microbiology 1999, 145, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, H.; Moskaug, J.O.; Fromm, S.H.; Blomhoff, R. In vivo imaging of NF-kappa b activity. J. Immunol. 2002, 168, 1441–1446. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5'-3') | Expected Amplicon Size (bp) |
|---|---|---|
| 16S rRNA | Forward: TAGGGTTTCTCTTCGGAGCATCG | 123 |
| Reverse: AACTGAATGATGGCAACT | ||
| cps2J | Forward: AGAGTGTTTAGATAGCATTATTTC | 198 |
| Reverse: TAATTTGCTGTGCTATTTTTGATAC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haas, B.; Grenier, D. Impact of Sub-Inhibitory Concentrations of Amoxicillin on Streptococcus suis Capsule Gene Expression and Inflammatory Potential. Pathogens 2016, 5, 37. https://doi.org/10.3390/pathogens5020037
Haas B, Grenier D. Impact of Sub-Inhibitory Concentrations of Amoxicillin on Streptococcus suis Capsule Gene Expression and Inflammatory Potential. Pathogens. 2016; 5(2):37. https://doi.org/10.3390/pathogens5020037
Chicago/Turabian StyleHaas, Bruno, and Daniel Grenier. 2016. "Impact of Sub-Inhibitory Concentrations of Amoxicillin on Streptococcus suis Capsule Gene Expression and Inflammatory Potential" Pathogens 5, no. 2: 37. https://doi.org/10.3390/pathogens5020037