Animal Models of Tick-Borne Hemorrhagic Fever Viruses

Abstract
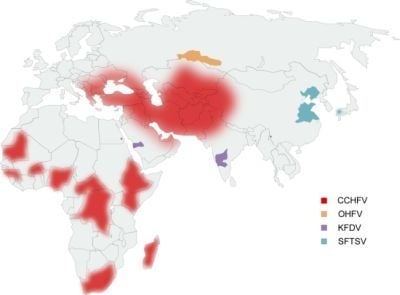
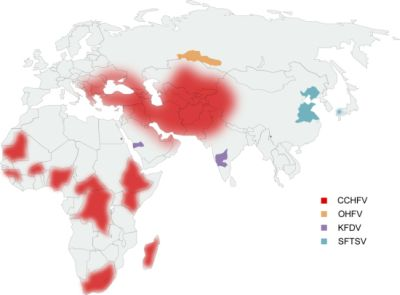
:
1. Introduction
2. Omsk Hemorrhagic Fever Virus
2.1. Epidemiology
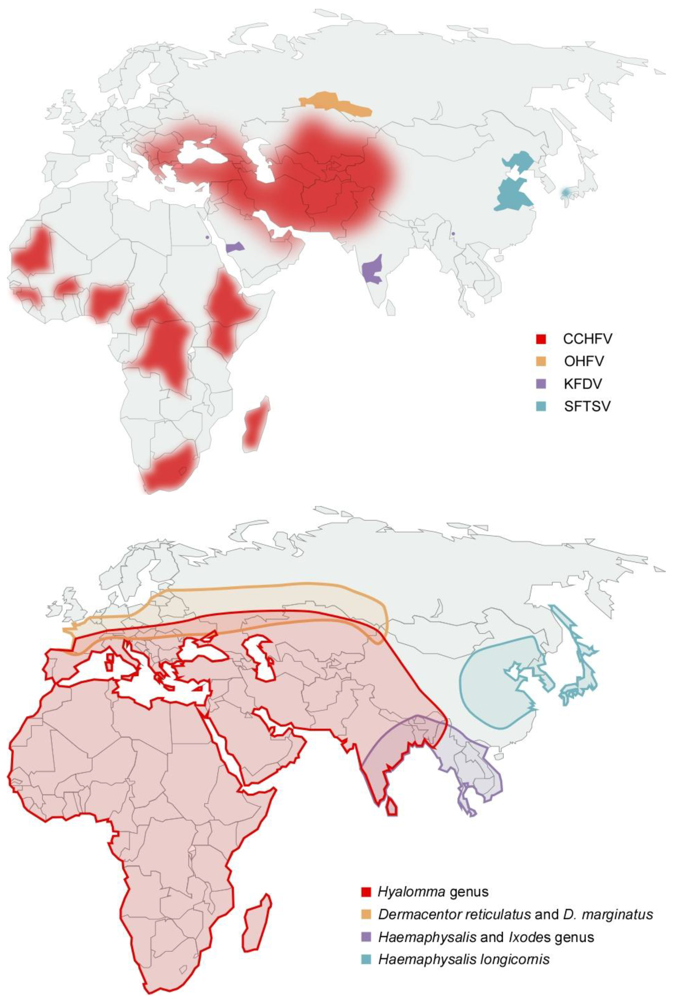
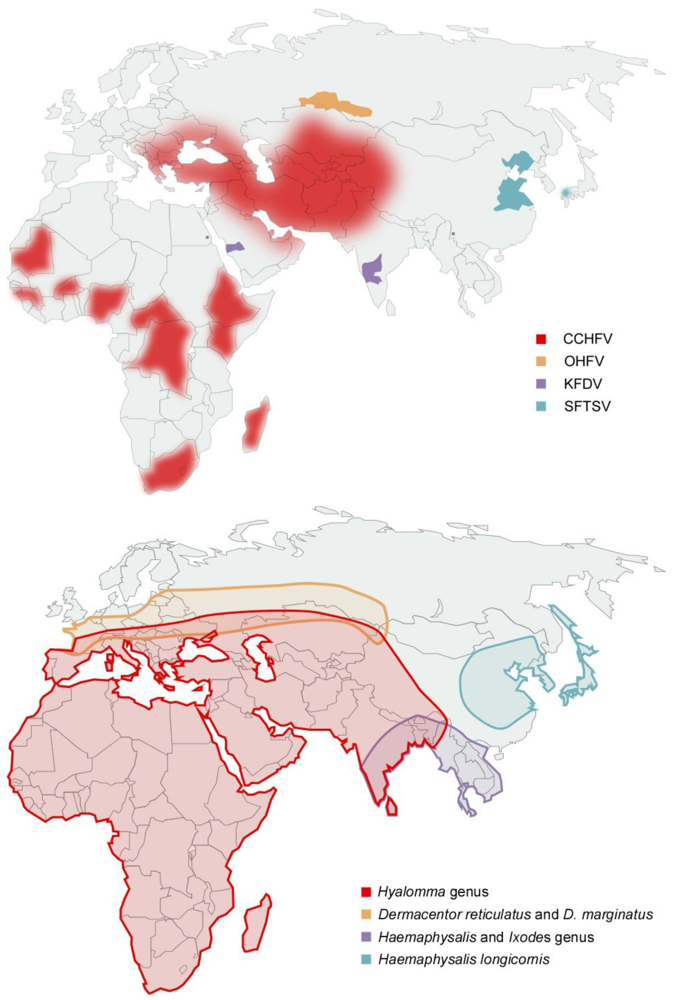
2.2. Clinical Presentation

{kind=link}
{kind=link}
| Species | Human | Mouse |
|---|---|---|
| Incubation period | 3–7 days (average) | 5–8 days |
| Disease duration | 8–14 days (up to 5 weeks in biphasic patients) | 9–13 days |
| Biphasic | Yes | No |
| Disease signs | Fever, cephalalgia, cough, myalgia, dehydration, gastrointestinal symptoms, facial hyperemia, epistaxis, gingival and vaginal hemorrhage, hemorrhagic rash, hepatomegaly | Weight loss, piloerection, lethargy, hunched posture, conjunctival suffusion, neurological signs |
| Hematologic changes | Thrombocytopenia, leukopenia, neutrophilia, monocytosis | Leukopenia, neutrophilia |
| Pathologic changes | Respiratory tract, uterus, liver, GI tract | Brain, thymus, spleen, urinary tract, GI tract |
| Case fatality | 0.4–2.5% | 100% |
2.3. Animal Models of Omsk Hemorrhagic Fever
2.4. Omsk Hemorrhagic Fever Virus Summary
3. Kyasanur Forest Disease Virus
3.1. Epidemiology
3.2. Clinical Presentation
| Species | Human | Bonnet Macaque | Mouse |
|---|---|---|---|
| Incubation period | 3–8 days | 3 days | 3–5 days |
| Disease duration | 8–12 days | 3–8 days | 6–8 days |
| Biphasic | Yes | No | No |
| Disease signs | Fever, cephalalgia, myalgia, diarrhea, cough, hepatomegaly, splenomegaly, epistaxis, bradycardia, oral and intestinal hemorrhage, meningoencephalitis | Fever, diarrhea, bradycardia, hypotension | Paralysis, weight loss |
| Hematologic changes | Leukopenia, thrombocytopenia, neutropenia, eosinopenia, elevated liver enzymes | thrombocytopenia, leukopenia, elevated liver enzymes | Not reported |
| Pathologic changes | Kidney, liver, lung | Kidney, liver, lung, spleen and lymph nodes | Brain, lungs |
| Case fatality | 2–10% | 100% | ≥97% |
3.3. Animal Models of Kyasanur Forest Disease
3.3.1. Rodent Models of Kyasanur Forest Disease
3.3.2. Non-human Primate Models of Kyasanur Forest Disease
3.4. Kyasanur Forest Disease Summary
4. Severe Fever with Thrombocytopenia Syndrome Virus
4.1. Epidemiology
4.2. Clinical Presentation
4.3. Rodent Models of Severe Fever with Thrombocytopenia Syndrome
| Species | Human | Mouse | Suckling Mouse | |
|---|---|---|---|---|
| Untreated | Mitomycin C treated | |||
| Incubation period | ≤21 days | ≤3 days | Not reported | 6–9 days |
| Disease duration | 3–7 days | ≤18 days | ≤21 days | 2–8 days |
| Biphasic | No | No | No | No |
| Disease signs | Fever, cephalalgia, myalgia, arthralgia, dizziness, malaise, mucosal hemorrhage, hemorrhagic rash, multi-organ failure, disseminated intravascular coagulopathy, neurological symptoms | None | Weight Loss | Weight loss, piloerection, lethargy and hind limb paralysis |
| Hematologic changes | Leukocytopenia, thrombocytopenia, elevated liver enzymes | Leukocytopenia, thrombocytopenia, elevated liver enzymes | Unknown | Elevated total bilirubin, decrease in albumin |
| Pathologic changes | Unknown | Liver, spleen | Unknown | Liver |
| Case fatality | 12% | 0% | 50% | 35–100% |
4.4. Severe Fever with Thrombocytopenia Syndrome Virus Summary
5. Crimean-Congo Hemorrhagic Fever Virus
5.1. Epidemiology
5.2. Clinical Presentation
| Species | Human | Newborn mice | IFNAR−/− mice | STAT1−/− mice |
|---|---|---|---|---|
| Incubation period | 1–7 days | ≤3 days | ≤3 days | ≤2 days |
| Disease duration | 10–12 days (average) | 4–5 days | 2–3 days | 1–2 days |
| Biphasic | No | No | No | No |
| Disease signs | Fever, cephalalgia, myalgia, arthralgia, dizziness, malaise, mucosal hemorrhage, hemorrhagic rash, multi-organ failure, disseminated intravascular coagulopathy | Weight loss, piloerection, lethargy, hind limb paralysis | Weight loss, piloerection, lethargy, occasional hemorrhage | Weight loss, piloerection, lethargy, occasional hemorrhage |
| Hematologic changes | Leukocytopenia, thrombocytopenia, increase clotting times, increased pro-inflammatory cytokines/chemokines, elevated liver enzymes | Unknown | Thrombocytopenia, increased clotting times, elevated liver enzymes, increased pro-inflammatory cytokines/chemokines | Thrombocytopenia, elevated liver enzymes, increased pro-inflammatory cytokines/chemokines |
| Pathologic changes | Liver, spleen, lung | Liver, spleen, lymph node | Liver, spleen | |
| Case fatality | 3–60% | 100% | 100% | 100% |
5.3. Animal Models of Crimean-Congo Hemorrhagic Fever
5.4. Crimean-Congo Hemorrhagic Fever virus Summary
6. Conclusions
Acknowledgements
Conflict of Interest
References
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–1955. [Google Scholar] [CrossRef]
- Suss, J. Tick-borne encephalitis 2010: epidemiology, risk areas, and virus strains in Europe and Asia-an overview. Ticks Tick Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus - a review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Ruzek, D.; Dobler, G.; Donoso Mantke, O. Tick-borne encephalitis: pathogenesis and clinical implications. Travel Med. Infect. Dis. 2010, 8, 223–232. [Google Scholar] [CrossRef]
- Dorrbecker, B.; Dobler, G.; Spiegel, M.; Hufert, F.T. Tick-borne encephalitis virus and the immune response of the mammalian host. Travel Med. Infect. Dis. 2010, 8, 213–222. [Google Scholar] [CrossRef]
- Turtle, L.; Griffiths, M.J.; Solomon, T. Encephalitis caused by flaviviruses. QJM 2012, 105, 219–223. [Google Scholar] [CrossRef]
- Sips, G.J.; Wilschut, J.; Smit, J.M. Neuroinvasive flavivirus infections. Rev. Med. Virol. 2012, 22, 69–87. [Google Scholar] [CrossRef]
- Ternovoi, V.A.; Kurzhukov, G.P.; Sokolov, Y.V.; Ivanov, G.Y.; Ivanisenko, V.A.; Loktev, A.V.; Ryder, R.W.; Netesov, S.V.; Loktev, V.B. Tick-borne encephalitis with hemorrhagic syndrome, Novosibirsk region, Russia, 1999. Emerg. Infect. Dis. 2003, 9, 743–746. [Google Scholar] [CrossRef]
- Ruzek, D.; Yakimenko, V.V.; Karan, L.S.; Tkachev, S.E. Omsk haemorrhagic fever. Lancet 2010, 376, 2104–2113. [Google Scholar] [CrossRef]
- Pattnaik, P. Kyasanur forest disease: an epidemiological view in India. Rev. Med. Virol. 2006, 16, 151–165. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, C.; Zhan, F.; Wang, X.; Liang, M.; Zhang, Q.; Ding, S.; Guan, X.; Huo, X.; Li, C.; et al. Host cytokine storm is associated with disease severity of severe fever with thrombocytopenia syndrome. J. Infect. Dis. 2012, 206, 1085–1094. [Google Scholar] [CrossRef]
- Gai, Z.T.; Zhang, Y.; Liang, M.F.; Jin, C.; Zhang, S.; Zhu, C.B.; Li, C.; Li, X.Y.; Zhang, Q.F.; Bian, P.F.; et al. Clinical progress and risk factors for death in severe fever with thrombocytopenia syndrome patients. J. Infect. Dis. 2012, 206, 1095–1102. [Google Scholar] [CrossRef]
- Casals, J.; Henderson, B.E.; Hoogstraal, H.; Johnson, K.M.; Shelokov, A. A review of Soviet viral hemorrhagic fevers, 1969. J. Infect. Dis. 1970, 122, 437–453. [Google Scholar] [CrossRef]
- Kolonin, G.V. Fauna of Ixodid Ticks of the World (Acari, Ixodidae). 2009. Available online: http://www.kolonin.org/1.html (access on 22 May 2013).
- Holbrook, M.R.; Aronson, J.F.; Campbell, G.A.; Jones, S.; Feldmann, H.; Barrett, A.D. An animal model for the tickborne flavivirus--Omsk hemorrhagic fever virus. J. Infect. Dis. 2005, 191, 100–108. [Google Scholar] [CrossRef]
- Kenyon, R.H.; Rippy, M.K.; McKee, K.T., Jr.; Zack, P.M.; Peters, C.J. Infection of Macaca radiata with viruses of the tick-borne encephalitis group. Microb. Pathog. 1992, 13, 399–409. [Google Scholar] [CrossRef]
- Tigabu, B.; Juelich, T.; Holbrook, M.R. Comparative analysis of immune responses to Russian spring-summer encephalitis and Omsk hemorrhagic fever viruses in mouse models. Virology 2010, 408, 57–63. [Google Scholar] [CrossRef]
- Tigabu, B.; Juelich, T.; Bertrand, J.; Holbrook, M.R. Clinical evaluation of highly pathogenic tick-borne flavivirus infection in the mouse model. J. Med. Virol. 2009, 81, 1261–1269. [Google Scholar] [CrossRef]
- Shestopalova, N.M.; Reingold, V.N.; Gagarina, A.V.; Kornilova, E.A.; Popov, G.V.; Chumakov, M.P. Electron microscopic study of the central nervous system in mice infected by Omsk hemorrhagic fever (OHF) virus. Virus reproduction in cerebellum neurons. J. Ultrastruct. Res. 1972, 40, 458–469. [Google Scholar] [CrossRef]
- Pripuzova, N.S.; Gmyl, L.V.; Romanova, L.; Tereshkina, N.V.; Rogova, Y.V.; Terekhina, L.L.; Kozlovskaya, L.I.; Vorovitch, M.F.; Grishina, K.G.; Timofeev, A.V.; et al. Exploring of primate models of tick-borne flaviviruses infection for evaluation of vaccines and drugs efficacy. PLoS One 2013, 8, e61094. [Google Scholar] [CrossRef]
- Work, T.H.; Trapido, H. Kyasanur Forest Disease A New Virus Disease in India. Indian J. Med. Sci. 1957, 11, 341–345. [Google Scholar]
- Iyer, C.G.S.; Work, T.H.; Narasimha Murthy, D.P.; Trapido, H.; Rajagopalan, P.K. Pathological Findings in Monkeys, Presbytis entellus and Macaca radiata, Found Dead in the Forest. Indian J. Med. Res. 1960, 48, 276–286. [Google Scholar]
- Pavri, K. Clinical, clinicopathologic, and hematologic features of Kyasanur Forest disease. Rev. Infect. Dis. 1989, 11 (Suppl 4), S854–S859. [Google Scholar] [CrossRef]
- Verma, M.G.; Webb, H.E.; Pavri, K. Studies on the transmission of Kyasanur Forest disease virus by Haemaphysalis spinigera Newman. Transfusion (Paris) 1960, 54, 509–516. [Google Scholar]
- Trapido, H.; Rajagopalan, P.K.; Work, T.H.; Varma, M.G.R. Isolation of Kyasanur Forest Disease Virus From Naturally Infected Ticks of the Genus Haemaphysalis. Indian J. Med. Res. 1959, 47, 133–138. [Google Scholar]
- Singh, K.R.; Pavri, K.; Anderson, C.R. Experimental Transovarial Transmission of Kyasanur Forest Disease Virus in Haemaphysalis Spinigera. Nature 1963, 199, 513. [Google Scholar] [CrossRef]
- Singh, K.R.; Pavri, K.; Anderson, C.R. Transmission of Kyasanur forest disease by Haemaphysalis tururis, Haemaphysalis papuana kinneari and Haemaphysalis minuta. Indian J. Med. Res. 1964, 52, 566–573. [Google Scholar]
- Bhat, H.R.; Sreenivasan, M.K.; Goverdhan, M.K.; Naik, S.V. Transmission of Kyasanur Forest Disease Virus by Haemaphysalis Kyasanurensis Trapido, Hoogstraal and Rajagopalan, 1964 (Acarina : Ixodidae). Indian J. Med. Res. 1975, 63, 879–887. [Google Scholar]
- Singh, K.R.; Bhatt, P.N. Transmission of Kyasanur Forest disease virus by Hyalomma marginatum isaaci. Indian J. Med. Res. 1968, 56, 610–613. [Google Scholar]
- Singh, K.R.; Goverdhan, M.K.; Rao, T.R. Experimental transmission of Kyasanur forest disease virus to small mammals by Ixodes petauristae, I. ceylonensis and Haemaphysalis spinigera. Indian J. Med. Res. 1968, 56, 594–609. [Google Scholar]
- Singh, K.R.; Goverdhan, M.K.; Bhat, U.K. Transmission of Kyasanur Forest disease virus by soft tick, Argas persicus (Ixodoidea: Argasidae). Indian J. Med. Res. 1971, 59, 213–218. [Google Scholar]
- Bhat, U.K.; Goverdhan, M.K. Transmission of Kyasanur Forest disease virus by the soft tick, Ornithodoros crossi. Acta Virol. 1973, 17, 337–342. [Google Scholar]
- Boshell, J.; Rajagopalan, P.K. Preliminary studies on experimental transmission of Kyasanur Forest disease virus by nymphs of Ixodes petauristae Warburton, 1933, infected as larvae on Suncus murinus and Rattus blanfordi. Indian J. Med. Res. 1968, 56, 589–593. [Google Scholar]
- Boshell, J.; Rajagopalan, P.K.; Patil, A.P.; Pavri, K.M. Isolation of Kyasanur Forest disease virus from ixodid ticks: 1961–1964. Indian J. Med. Res. 1968, 56, 541–568. [Google Scholar]
- Boshell, J. Kyasanur Forest disease: ecologic considerations. Am. J. Trop. Med. Hyg. 1969, 18, 67–80. [Google Scholar]
- Wang, J.; Zhang, H.; Fu, S.; Wang, H.; Ni, D.; Nasci, R.; Tang, Q.; Liang, G. Isolation of kyasanur forest disease virus from febrile patient, yunnan, china. Emerg. Infect. Dis. 2009, 15, 326–328. [Google Scholar] [CrossRef]
- Mehla, R.; Kumar, S.R.; Yadav, P.; Barde, P.V.; Yergolkar, P.N.; Erickson, B.R.; Carroll, S.A.; Mishra, A.C.; Nichol, S.T.; Mourya, D.T. Recent ancestry of Kyasanur Forest disease virus. Emerg. Infect. Dis. 2009, 15, 1431–1437. [Google Scholar] [CrossRef]
- Madani, T.A. Alkhumra virus infection, a new viral hemorrhagic fever in Saudi Arabia. J. Infect. 2005, 51, 91–97. [Google Scholar] [CrossRef]
- Zaki, A.M. Isolation of a flavivirus related to the tick-borne encephalitis complex from human cases in Saudi Arabia. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 179–181. [Google Scholar] [CrossRef]
- Charrel, R.N.; Gould, E.A. Alkhurma hemorrhagic fever in travelers returning from Egypt, 2010. Emerg. Infect. Dis. 2011, 17, 1573–1574; author reply 1574. [Google Scholar]
- Madani, T.A.; Kao, M.; Azhar, E.I.; Abuelzein el, T.M.; Al-Bar, H.M.; Abu-Araki, H.; Ksiazek, T.G. Successful propagation of Alkhumra (misnamed as Alkhurma) virus in C6/36 mosquito cells. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 180–185. [Google Scholar] [CrossRef]
- Madani, T.A.; Azhar, E.I.; Abuelzein el, T.M.; Kao, M.; Al-Bar, H.M.; Abu-Araki, H.; Niedrig, M.; Ksiazek, T.G. Alkhumra (Alkhurma) virus outbreak in Najran, Saudi Arabia: epidemiological, clinical, and laboratory characteristics. J. Infect. 2011, 62, 67–76. [Google Scholar] [CrossRef]
- Dodd, K.A.; Bird, B.H.; Khristova, M.L.; Albarino, C.G.; Carroll, S.A.; Comer, J.A.; Erickson, B.R.; Rollin, P.E.; Nichol, S.T. Ancient ancestry of KFDV and AHFV revealed by complete genome analyses of viruses isolated from ticks and mammalian hosts. PLoS Negl. Trop. Dis. 2011, 5, e1352. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses. January 2013. Available online: http://www.ictvonline.org/virusTaxonomy.asp?bhcp=1 (access on 22 May 2013).
- Memish, Z.A.; Fagbo, S.F.; Assiri, A.M.; Rollin, P.; Zaki, A.M.; Charrel, R.; Mores, C.; MacNeil, A. Alkhurma viral hemorrhagic fever virus: proposed guidelines for detection, prevention, and control in Saudi Arabia. PLoS Negl. Trop. Dis. 2012, 6, e1604. [Google Scholar] [CrossRef]
- Bhat, H.R.; Sreenivasan, M.A.; Goverdhan, M.K.; Naik, S.V.; Banerjee, K. Susceptibility of Hystrix indica Kerr, 1792, Indian crested porcupine (Rodentia Hystricidae) to KFD virus. Indian J. Med. Res. 1976, 64, 1566–1570. [Google Scholar]
- Sreenivasan, M.A.; Bhat, H.R. Susceptibility of Vandeleuria oleracea Bennet, 1832 (Rodentia, Muridae) to experimental infection with Kyasanur forest disease virus. Indian J. Med. Res. 1976, 64, 568–572. [Google Scholar]
- Goverdhan, M.K.; Anderson, C.R. The reaction of Mus platythrix to Kyasanur Forest Disease Virus. Indian J. Med. Res. 1972, 60, 1002–1006. [Google Scholar]
- Sreenivasan, M.A.; Bhat, H.R. Susceptibility of Lepus nigricollis Cuvier, to experimental infection with Kyasanur Forest Disease virus. Indian J. Med. Res. 1977, 65, 17–20. [Google Scholar]
- Goverdhan, M.K.; Anderson, C.R. The reaction of Funambulus tristriatus tristriatus Rattus blanfordi and Suncus murinus to Kyasanur forest disease virus. Indian J. Med. Res. 1981, 74, 141–146. [Google Scholar]
- Anderson, C.R.; Singh, K.R. The reaction of cattle to Kyasanur Forest disease virus. Indian J. Med. Res. 1971, 59, 195–198. [Google Scholar]
- Webb, H.E. Kyasanur Forest Disease Virus in Three Species on Rodents. Trans. R. Soc. Trop. Med. Hyg. 1965, 59, 205–211. [Google Scholar] [CrossRef]
- Goverdhan, M.K.; Rajagopalan, P.K.; Narasimha Murthy, D.P.; Upadhyaya, S.; Boshell, M.J.; Trapido, H.; Ramachandra Rao, T. Epizootiology of Kyasanur Forest Disease in wild monkeys of Shimoga district, Mysore State (1957–1964). Indian J. Med. Res. 1974, 62, 497–510. [Google Scholar]
- Webb, H.E.; Burston, J. Clinical and pathological observations with special reference to the nervous system in Macaca radiata infected with Kyasanur Forest Disease virus. Trans. R. Soc. Trop. Med. Hyg. 1966, 60, 325–331. [Google Scholar] [CrossRef]
- Work, T.H. Russian Spring-Summer Virus in India - Kyasanur Forest Disease. Prog. Med. Virol. 1958, 1, 248–279. [Google Scholar]
- Nayar, M. Kyasanur Forest disease virus encephalitis in mice. Effect of cortisone on the encephalitic lesions. Indian J. Med. Res. 1972, 60, 1427–1431. [Google Scholar]
- Nayar, M. Histological changes in mice infected with Kyasanur Forest disease virus. Indian J. Med. Res. 1972, 60, 1421–1426. [Google Scholar]
- Price, W.H. Chronic Disease and Virus Persistence in Mice Inoculated with Kyasanur Forest Disease Virus. Virology 1966, 29, 679–681. [Google Scholar] [CrossRef]
- Mansharamani, H.J.; Dandawate, C.N. Experimental vaccine against Kyasanur Forest disease (KFD) virus from tissue culture source. II. Safety testing of the vaccine in cortisone sensitized Swiss albino mice. Indian J. Pathol. Bacteriol. 1967, 10, 25–32. [Google Scholar]
- Mansharamani, H.J.; Dandawate, C.N.; Krishnamurthy, B.G. Experimental vaccine against Kyasanur Forest disease (KFD) virus from tissue culture source. I. Some data on the preparation and antigenicity tests of vaccines. Indian J. Pathol. Bacteriol. 1967, 10, 9–24. [Google Scholar]
- Xu, B.; Liu, L.; Huang, X.; Ma, H.; Zhang, Y.; Du, Y.; Wang, P.; Tang, X.; Wang, H.; Kang, K.; et al. Metagenomic Analysis of Fever, Thrombocytopenia and Leukopenia Syndrome (FTLS) in Henan Province, China: Discovery of a New Bunyavirus. PLoS Pathog. 2011, 7, e1002369. [Google Scholar] [CrossRef]
- Yu, X.-J.; Liang, M.-F.; Zhang, S.-Y.; Liu, Y.; Li, J.-D.; Sun, Y.-L.; Zhang, L.; Zhang, Q.-F.; Popov, V.L.; Li, C.; et al. Fever with Thrombocytopenia Associated with a Novel Bunyavirus in China. New Eng. J. Med. 2011, 364, 1523–1532. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Zhou, D.J.; Xiong, Y.; Chen, X.P.; He, Y.W.; Sun, Q.; Yu, B.; Li, J.; Dai, Y.A.; Tian, J.H.; et al. Hemorrhagic fever caused by a novel tick-borne Bunyavirus in Huaiyangshan, China. Zhonghua Liu Xing Bing Xue Za Zhi 2011, 32, 209–220. [Google Scholar]
- Ministry of Health, Labor and Welfare, report on SFTSV. 11 February 2013. Available online: http://www.mhlw.go.jp/stf/houdou/2r9852000002u1pm.html (access on 22 May 2013).
- ProMED Severe Fever With Thrombocytopenia Syndrome - Japan: (Ehime) Fatalities. 14 Feb. 2013. Available online: http://www.promedmail.org/direct.php?id=20130214.1542725 (access on 22 May 2013).
- Chen, H.; Hu, K.; Zou, J.; Xiao, J. A cluster of cases of human-to-human transmission caused by severe fever with thrombocytopenia syndrome bunyavirus. Int. J. Infect. Dis. 2012, 17, e206–e208. [Google Scholar] [CrossRef]
- Bao, C.J.; Guo, X.L.; Qi, X.; Hu, J.L.; Zhou, M.H.; Varma, J.K.; Cui, L.B.; Yang, H.T.; Jiao, Y.J.; Klena, J.D.; et al. A Family Cluster of Infections by a Newly Recognized Bunyavirus in Eastern China, 2007: Further Evidence of Person-to-Person Transmission. Clin. Infect. Dis. 2011, 53, 1208–1214. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Q.; Hu, W.; Wu, J.; Wang, Y.; Mei, L.; Walker, D.H.; Ren, J.; Yu, X.J. Person-to-person transmission of severe fever with thrombocytopenia syndrome virus. Vector Borne Zoonotic Dis. 2012, 12, 156–160. [Google Scholar] [CrossRef]
- Gai, Z.; Liang, M.; Zhang, Y.; Zhang, S.; Jin, C.; Wang, S.W.; Sun, L.; Zhou, N.; Zhang, Q.; Sun, Y.; et al. Person-to-person transmission of severe fever with thrombocytopenia syndrome bunyavirus through blood contact. Clin. Infect. Dis. 2012, 54, 249–252. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Zhang, Y.; Wang, H.; Wang, C.; Sun, J.; Lin, J.; Chai, C.; Yan, J. Severe fever with thrombocytopenia syndrome bunyavirus (SFTSV) infections in Zhejiang Province, China. Int. J. Infect. Dis. 2012, 17, e137–e138. [Google Scholar]
- Liu, L.; Guan, X.H.; Xing, X.S.; Shen, X.F.; Xu, J.Q.; Yue, J.L.; Huo, X.X.; Sha, S.; Wu, H.X.; Huang, J.; et al. Epidemiologic analysis on severe fever with thrombocytopenia syndrome in Hubei province, 2010. Zhonghua Liu Xing Bing Xue Za Zhi 2012, 33, 168–172. [Google Scholar]
- Jiao, Y.; Zeng, X.; Guo, X.; Qi, X.; Zhang, X.; Shi, Z.; Zhou, M.; Bao, C.; Zhang, W.; Xu, Y.; et al. Preparation and evaluation of recombinant severe fever with thrombocytopenia syndrome virus nucleocapsid protein for detection of total antibodies in human and animal sera by double-antigen sandwich enzyme-linked immunosorbent assay. J. Clin. Microbiol. 2012, 50, 372–377. [Google Scholar] [CrossRef]
- Chen, X.P.; Cong, M.L.; Li, M.H.; Kang, Y.J.; Feng, Y.M.; Plyusnin, A.; Xu, J.; Zhang, Y.Z. Infection and pathogenesis of Huaiyangshan virus (a novel tick-borne bunyavirus) in laboratory rodents. J. Gen. Virol. 2012, 93, 1288–1293. [Google Scholar] [CrossRef]
- Jin, C.; Liang, M.; Ning, J.; Gu, W.; Jiang, H.; Wu, W.; Zhang, F.; Li, C.; Zhang, Q.; Zhu, H.; et al. Pathogenesis of emerging severe fever with thrombocytopenia syndrome virus in C57/BL6 mouse model. Proc. Natl. Acad. Sci. USA 2012, 109, 10053–10058. [Google Scholar] [CrossRef]
- Ergonul, O. Crimean-Congo haemorrhagic fever. Lancet Infect. Dis. 2006, 6, 203–214. [Google Scholar] [CrossRef]
- Whitehouse, C.A. Crimean-Congo hemorrhagic fever. Antiviral Res. 2004, 64, 145–160. [Google Scholar]
- Flick, R.; Whitehouse, C.A. Crimean-Congo hemorrhagic fever virus. Curr. Mol. Med. 2005, 5, 753–760. [Google Scholar] [CrossRef]
- Vorou, R.; Pierroutsakos, I.N.; Maltezou, H.C. Crimean-Congo hemorrhagic fever. Curr. Opin. Infect. Dis. 2007, 20, 495–500. [Google Scholar] [CrossRef]
- Chumakov, M.P.; Smirnova, S.E.; Tkachenko, E.A. Relationship between strains of Crimean haemorrhagic fever and Congo viruses. Acta Virol. 1970, 14, 82–85. [Google Scholar]
- Casals, J. Antigenic similarity between the virus causing Crimean hemorrhagic fever and Congo virus. Proc. Soc. Exp. Biol. Med. 1969, 131, 233–236. [Google Scholar]
- Casals, J.; Tignor, G.H. Neutralization and hemagglutination-inhibition tests with Crimean hemorrhagic fever-Congo virus. Proc. Soc. Exp. Biol. Med. 1974, 145, 960–966. [Google Scholar]
- Ergonul, O. Crimean-Congo hemorrhagic fever virus: new outbreaks, new discoveries. Curr. Opin. Virol. 2012, 2, 215–220. [Google Scholar] [CrossRef]
- Gordon, S.W.; Linthicum, K.J.; Moulton, J.R. Transmission of Crimean-Congo hemorrhagic fever virus in two species of Hyalomma ticks from infected adults to cofeeding immature forms. Am. J. Trop. Med. Hyg. 1993, 48, 576–580. [Google Scholar]
- Zeller, H.G.; Cornet, J.P.; Camicas, J.L. Experimental transmission of Crimean-Congo hemorrhagic fever virus by west African wild ground-feeding birds to Hyalomma marginatum rufipes ticks. Am. J. Trop. Med. Hyg. 1994, 50, 676–681. [Google Scholar]
- Logan, T.M.; Linthicum, K.J.; Bailey, C.L.; Watts, D.M.; Moulton, J.R. Experimental transmission of Crimean-Congo hemorrhagic fever virus by Hyalomma truncatum Koch. Am. J. Trop. Med. Hyg. 1989, 40, 207–212. [Google Scholar]
- Dohm, D.J.; Logan, T.M.; Linthicum, K.J.; Rossi, C.A.; Turell, M.J. Transmission of Crimean-Congo hemorrhagic fever virus by Hyalomma impeltatum (Acari:Ixodidae) after experimental infection. J. Med. Entomol. 1996, 33, 848–851. [Google Scholar]
- Shepherd, A.J.; Swanepoel, R.; Cornel, A.J.; Mathee, O. Experimental studies on the replication and transmission of Crimean-Congo hemorrhagic fever virus in some African tick species. Am. J. Trop. Med. Hyg. 1989, 40, 326–331. [Google Scholar]
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J. Med. Entomol. 1979, 15, 307–417. [Google Scholar]
- Shepherd, A.J.; Leman, P.A.; Swanepoel, R. Viremia and antibody response of small African and laboratory animals to Crimean-Congo hemorrhagic fever virus infection. Am. J. Trop. Med. Hyg. 1989, 40, 541–547. [Google Scholar]
- Shepherd, A.J.; Swanepoel, R.; Shepherd, S.P.; McGillivray, G.M.; Searle, L.A. Antibody to Crimean-Congo hemorrhagic fever virus in wild mammals from southern Africa. Am. J. Trop. Med. Hyg. 1987, 36, 133–142. [Google Scholar]
- Swanepoel, R.; Leman, P.A.; Burt, F.J.; Jardine, J.; Verwoerd, D.J.; Capua, I.; Bruckner, G.K.; Burger, W.P. Experimental infection of ostriches with Crimean-Congo haemorrhagic fever virus. Epidemiol. Infect. 1998, 121, 427–432. [Google Scholar] [CrossRef]
- Kubar, A.; Haciomeroglu, M.; Ozkul, A.; Bagriacik, U.; Akinci, E.; Sener, K.; Bodur, H. Prompt administration of Crimean-Congo hemorrhagic fever (CCHF) virus hyperimmunoglobulin in patients diagnosed with CCHF and viral load monitorization by reverse transcriptase-PCR. Jpn. J. Infect. Dis. 2011, 64, 439–443. [Google Scholar]
- Schwarz, T.F.; Nsanze, H.; Longson, M.; Nitschko, H.; Gilch, S.; Shurie, H.; Ameen, A.; Zahir, A.R.; Acharya, U.G.; Jager, G. Polymerase chain reaction for diagnosis and identification of distinct variants of Crimean-Congo hemorrhagic fever virus in the United Arab Emirates. Am. J. Trop. Med. Hyg. 1996, 55, 190–196. [Google Scholar]
- Khan, A.S.; Maupin, G.O.; Rollin, P.E.; Noor, A.M.; Shurie, H.H.; Shalabi, A.G.; Wasef, S.; Haddad, Y.M.; Sadek, R.; Ijaz, K.; et al. An outbreak of Crimean-Congo hemorrhagic fever in the United Arab Emirates, 1994-1995. Am. J. Trop. Med. Hyg. 1997, 57, 519–525. [Google Scholar]
- Rodriguez, L.L.; Maupin, G.O.; Ksiazek, T.G.; Rollin, P.E.; Khan, A.S.; Schwarz, T.F.; Lofts, R.S.; Smith, J.F.; Noor, A.M.; Peters, C.J.; et al. Molecular investigation of a multisource outbreak of Crimean-Congo hemorrhagic fever in the United Arab Emirates. Am. J. Trop. Med. Hyg. 1997, 57, 512–518. [Google Scholar]
- Cevik, M.A.; Erbay, A.; Bodur, H.; Eren, S.S.; Akinci, E.; Sener, K.; Onguru, P.; Kubar, A. Viral load as a predictor of outcome in Crimean-Congo hemorrhagic fever. Clin. Infect. Dis. 2007, 45, e96–e100. [Google Scholar] [CrossRef]
- Cevik, M.A.; Erbay, A.; Bodur, H.; Gulderen, E.; Bastug, A.; Kubar, A.; Akinci, E. Clinical and laboratory features of Crimean-Congo hemorrhagic fever: predictors of fatality. Int. J. Infect. Dis. 2008, 12, 374–379. [Google Scholar]
- Onguru, P.; Dagdas, S.; Bodur, H.; Yilmaz, M.; Akinci, E.; Eren, S.; Ozet, G. Coagulopathy parameters in patients with Crimean-Congo hemorrhagic fever and its relation with mortality. J. Clin. Lab. Anal. 2010, 24, 163–166. [Google Scholar] [CrossRef]
- Papa, A.; Bino, S.; Velo, E.; Harxhi, A.; Kota, M.; Antoniadis, A. Cytokine levels in Crimean-Congo hemorrhagic fever. J. Clin. Virol. 2006, 36, 272–276. [Google Scholar] [CrossRef]
- Saksida, A.; Duh, D.; Wraber, B.; Dedushaj, I.; Ahmeti, S.; Avsic-Zupanc, T. Interacting roles of immune mechanisms and viral load in the pathogenesis of crimean-congo hemorrhagic fever. Clin. Vaccine Immunol. 2010, 17, 1086–1093. [Google Scholar] [CrossRef]
- Weber, F.; Mirazimi, A. Interferon and cytokine responses to Crimean Congo hemorrhagic fever virus; an emerging and neglected viral zonoosis. Cytokine Growth Factor Rev. 2008, 19, 395–404. [Google Scholar] [CrossRef]
- Yesilyurt, M.; Gul, S.; Ozturk, B.; Kayhan, B.C.; Celik, M.; Uyar, C.; Erdogan, F. The early prediction of fatality in Crimean Congo hemorrhagic fever patients. Saudi Med. J. 2011, 32, 742–743. [Google Scholar]
- Yilmaz, G.; Koksal, I.; Topbas, M.; Yilmaz, H.; Aksoy, F. The effectiveness of routine laboratory findings in determining disease severity in patients with Crimean-Congo hemorrhagic fever: severity prediction criteria. J. Clin. Virol. 2010, 47, 361–365. [Google Scholar] [CrossRef]
- Duh, D.; Saksida, A.; Petrovec, M.; Ahmeti, S.; Dedushaj, I.; Panning, M.; Drosten, C.; Avsic-Zupanc, T. Viral load as predictor of Crimean-Congo hemorrhagic fever outcome. Emerg. Infect. Dis. 2007, 13, 1769–1772. [Google Scholar] [CrossRef]
- Ergonul, O.; Celikbas, A.; Baykam, N.; Eren, S.; Dokuzoguz, B. Analysis of risk-factors among patients with Crimean-Congo haemorrhagic fever virus infection: severity criteria revisited. Clin. Microbiol. Infect. 2006, 12, 551–554. [Google Scholar] [CrossRef]
- Ergonul, O.; Tuncbilek, S.; Baykam, N.; Celikbas, A.; Dokuzoguz, B. Evaluation of serum levels of interleukin (IL)-6, IL-10, and tumor necrosis factor-alpha in patients with Crimean-Congo hemorrhagic fever. J. Infect. Dis. 2006, 193, 941–944. [Google Scholar] [CrossRef]
- Papa, A.; Drosten, C.; Bino, S.; Papadimitriou, E.; Panning, M.; Velo, E.; Kota, M.; Harxhi, A.; Antoniadis, A. Viral load and Crimean-Congo hemorrhagic fever. Emerg. Infect. Dis. 2007, 13, 805–806. [Google Scholar] [CrossRef]
- Wolfel, R.; Paweska, J.T.; Petersen, N.; Grobbelaar, A.A.; Leman, P.A.; Hewson, R.; Georges-Courbot, M.C.; Papa, A.; Gunther, S.; Drosten, C. Virus detection and monitoring of viral load in Crimean-Congo hemorrhagic fever virus patients. Emerg. Infect. Dis. 2007, 13, 1097–1100. [Google Scholar] [CrossRef]
- Burt, F.J.; Swanepoel, R.; Shieh, W.J.; Smith, J.F.; Leman, P.A.; Greer, P.W.; Coffield, L.M.; Rollin, P.E.; Ksiazek, T.G.; Peters, C.J.; et al. Immunohistochemical and in situ localization of Crimean-Congo hemorrhagic fever (CCHF) virus in human tissues and implications for CCHF pathogenesis. Arch. Pathol. Lab. Med. 1997, 121, 839–846. [Google Scholar]
- Smirnova, S.E. A comparative study of the Crimean hemorrhagic fever-Congo group of viruses. Arch. Virol. 1979, 62, 137–143. [Google Scholar] [CrossRef]
- Fagbami, A.H.; Tomori, O.; Fabiyi, A.; Isoun, T.T. Experimantal Congo virus (Ib -AN 7620) infection in primates. Virologie 1975, 26, 33–37. [Google Scholar]
- Blagoveshchenskaya, N.M.; Butenko, A.M.; Vyshnivetskaya, L.K.; Zavodova, T.I.; Zarubina, L.V.; Karinskaya, G.A.; Kuchin, V.V.; Milyutin, V.N.; Novikova, E.M.; Rubin, S.G.; et al. Experimental infection of horses with Crimean hemorrhagic fever virus. Report 2. Virological and serological observations. Mater. 16 Nauchn. Sess. Inst. Polio Virusn. Entsefalitov. (Moscow, October 1969) 1969, 2, 126–127. [Google Scholar]
- Tignor, G.H.; Hanham, C.A. Ribavirin efficacy in an in vivo model of Crimean-Congo hemorrhagic fever virus (CCHF) infection. Antiviral. Res. 1993, 22, 309–325. [Google Scholar] [CrossRef]
- Bente, D.A.; Alimonti, J.B.; Shieh, W.J.; Camus, G.; Stroher, U.; Zaki, S.; Jones, S.M. Pathogenesis and immune response of Crimean-Congo hemorrhagic fever virus in a STAT-1 knockout mouse model. J. Virol. 2010, 84, 11089–11100. [Google Scholar] [CrossRef]
- Bereczky, S.; Lindegren, G.; Karlberg, H.; Akerstrom, S.; Klingstrom, J.; Mirazimi, A. Crimean-Congo hemorrhagic fever virus infection is lethal for adult type I interferon receptor-knockout mice. J. Gen. Virol. 2010, 91, 1473–1477. [Google Scholar] [CrossRef]
- Zivcec, M.; Safronetz, D.; Scott, D.; Robertson, S.; Ebihara, H.; Feldmann, H. Lethal Crimean-Congo Hemorrhagic Fever virus infection in interferon alpha/beta receptor knockout mice is associated with high viral loads, pro-inflammatory responses and coagulopathy. J. Infect. Dis. 2013. [Google Scholar] [CrossRef]
- New drug and biological drug products; evidence needed to demonstrate effectiveness of new drugs when human efficacy studies are not ethical or feasible. Final rule. Fed. Regist. 2002, 67, 37988–37998.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zivcec, M.; Safronetz, D.; Feldmann, H. Animal Models of Tick-Borne Hemorrhagic Fever Viruses. Pathogens 2013, 2, 402-421. https://doi.org/10.3390/pathogens2020402
Zivcec M, Safronetz D, Feldmann H. Animal Models of Tick-Borne Hemorrhagic Fever Viruses. Pathogens. 2013; 2(2):402-421. https://doi.org/10.3390/pathogens2020402
Chicago/Turabian StyleZivcec, Marko, David Safronetz, and Heinz Feldmann. 2013. "Animal Models of Tick-Borne Hemorrhagic Fever Viruses" Pathogens 2, no. 2: 402-421. https://doi.org/10.3390/pathogens2020402