

Evaluation of Four Adjuvant Combinations, IVAX-1, IVAX-2, CpG-1826+Montanide ISA 720 VG and CpG-1018+Montanide ISA 720 VG, for Safety and for Their Ability to Elicit Protective Immune Responses in Mice against a Respiratory Challenge with Chlamydia muridarum
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stocks of C. muridarum
2.2. Cloning, Expression and Purification of C. muridarum MOMP
2.3. Mice Vaccination, Intranasal Challenge and Assessment of the Course of the Disease and the C. muridarum Infection in Mice
2.4. Determination of the Humoral Immune Responses following Vaccination
2.5. Evaluation of Memory-Cell-Mediated Immune Responses following Vaccination
2.6. Statistical Analyses
3. Results
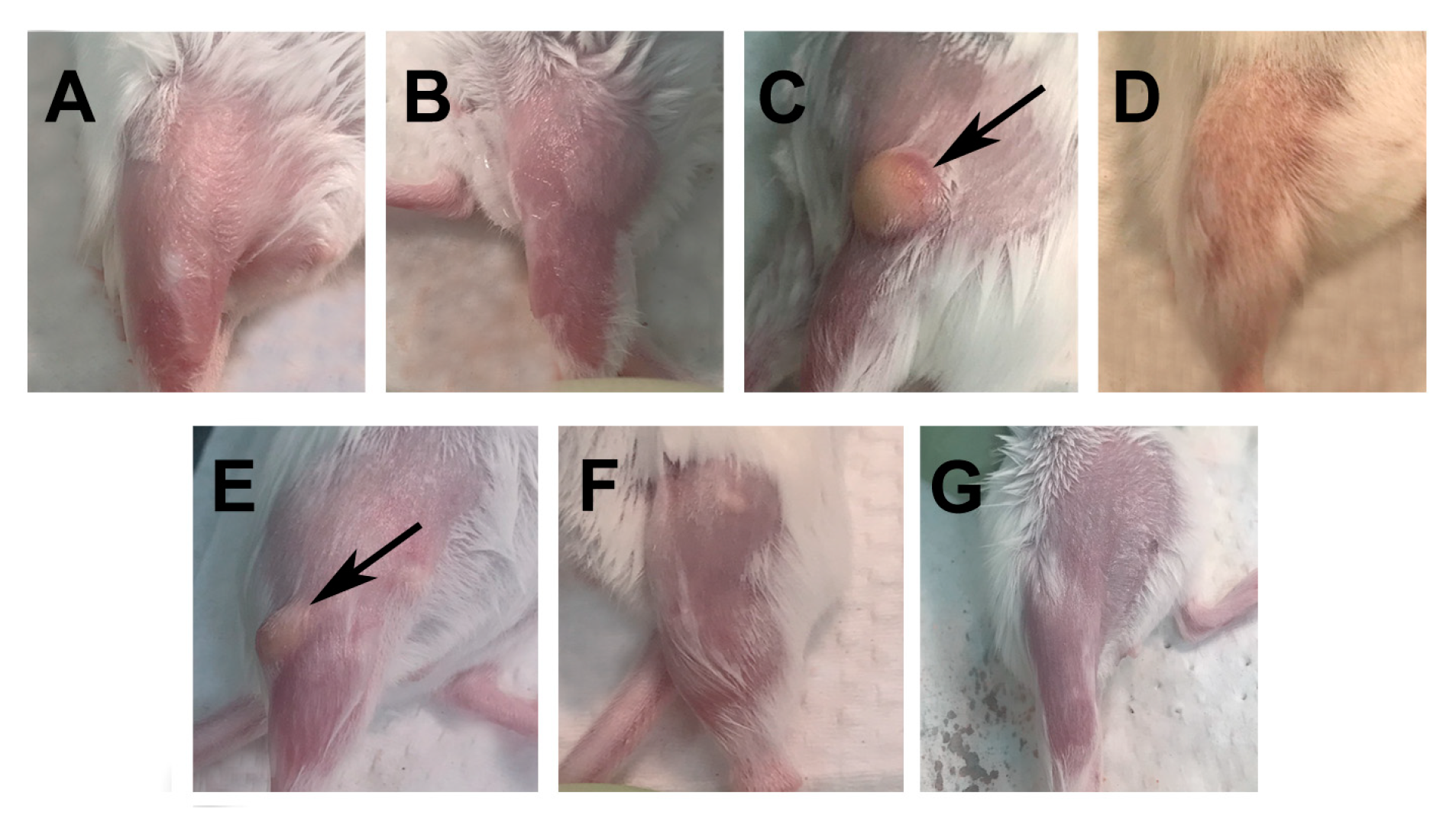
3.1. Assessment of the Immune Reaction at the Site of Vaccination
3.2. Humoral Immune Responses Elicited by Vaccination
3.3. Memory Cellular Immune Responses Induced by Vaccination
3.4. Changes in Body Weight of Mice following the C. muridarum i.n. Challenge
3.5. Lung Weights following the i.n. Challenge
3.6. Burden of C. muridarum Infection in the Lungs
3.7. Local Immune Responses in the Lungs at 10 d.p.c.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC. Sexually Transmitted Disease Surveillance 2019. In Prevention DoS; U.S. Department of Health and Human Services: Atlanta, GE, USA, 2021; pp. 1–168. [Google Scholar]
- Gottlieb, S.L.; Johnston, C. Future prospects for new vaccines against sexually transmitted infections. Curr. Opin. Infect. Dis. 2017, 30, 77–86. [Google Scholar] [CrossRef]
- Schachter, J.; Dawson, C.R. Human Chlamydial Infections; PSG Pub. Co.: Littleton, MA, USA, 1978; Volume xi, p. 273. [Google Scholar]
- Beem, M.O.; Saxon, E.M. Respiratory-Tract Colonization and a Distinctive Pneumonia Syndrome in Infants Infected with Chlamydia trachomatis. New Engl. J. Med. 1977, 296, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R. Trachoma: A Blinding Scourge from the Bronze Age to the Twenty-First Century, 1st ed.; Haddington Press Pry Ltd.: Melbourne, VIC, Australia, 2008. [Google Scholar]
- Nichols, R.L.; Bell, S.D., Jr.; Murray, E.S.; Haddad, N.A.; Bobb, A.A. Studies on trachoma. V. Clinical observations in a field trial of bivalent trachoma vaccine at three dosage levels in Saudi Arabia. Am. J. Trop. Med. Hyg. 1966, 15, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Dawson, C.; Wood, T.R.; Rose, L.; Hanna, L. Experimental Inclusion Conjunctivitis in Man. 3. Keratitis and other complications. Arch. Ophthalmol. 1967, 78, 341–349. [Google Scholar] [CrossRef]
- Morrison, R.P.; Belland, R.J.; Lyng, K.; Caldwell, H.D. Chlamydial disease pathogenesis. The 57-kD chlamydial hypersensitivity antigen is a stress response protein. J. Exp. Med. 1989, 170, 1271–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farris, C.M.; Morrison, R.P. Vaccination against Chlamydia Genital Infection Utilizing the Murine, C. muridarum Model. Infect. Immun. 2011, 79, 986–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockey, D.D.; Wang, J.; Lei, L.; Zhong, G. Chlamydia vaccine candidates and tools for chlamydial antigen discovery. Expert Rev. Vaccines 2009, 8, 1365–1377. [Google Scholar] [CrossRef] [PubMed]
- De la Maza, L.M.; Darville, T.L.; Pal, S. Chlamydia trachomatis vaccines for genital infections: Where are we and how far is there to go? Expert. Rev. Vaccines. 2021, 20, 1–15. [Google Scholar] [CrossRef]
- De la Maza, L.M.; Pal, S.; Olsen, A.W.; Follmann, F. Chlamydia Vaccines; Tan, M., Hegemann, J.H., Sutterlin, C., Eds.; Caister Academic Press: Norfolk, UK, 2020; pp. 339–383. [Google Scholar]
- Phillips, S.; Quigley, B.L.; Timms, P. Seventy Years of Chlamydia Vaccine Research—Limitations of the Past and Directions for the Future. Front. Microbiol. 2019, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Stephens, R.S.; Wagar, E.A.; Schoolniki, G.K. High-resolution mapping of serovar-specific and common antigenic determinants of the major outer membrane protein of Chlamydia trachomatis. J. Exp. Med. 1988, 167, 817–831. [Google Scholar] [CrossRef]
- Zhong, G.; Brunham, R.C.; de la Maza, L.M.; Darville, T.; Deal, C. National Institute of Allergy and Infectious Diseases workshop report: “Chlamydia vaccines: The way forward”. Vaccine 2017, 37, 7346–7354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Maza, L.M.; Zhong, G.; Brunham, R.C. Update on Chlamydia trachomatis Vaccinology. Clin. Vaccine Immunol. 2017, 24, e00543-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Peterson, E.M.; de la Maza, L.M. Vaccination with the Chlamydia trachomatis Major Outer Membrane Protein Can Elicit an Immune Response as Protective as That Resulting from Inoculation with Live Bacteria. Infect. Immun. 2005, 73, 8153–8160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Pal, S.; Weiland, J.; Peterson, E.M.; de la Maza, L.M. Protection against an intranasal challenge by vaccines formulated with native and recombinant preparations of the Chlamydia trachomatis major outer membrane protein. Vaccine 2009, 27, 5020–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kari, L.; Whitmire, W.M.; Crane, D.D.; Reveneau, N.; Carlson, J.H.; Goheen, M.M.; Peterson, E.M.; Pal, S.; de la Maza, L.M.; Caldwell, H.D. Chlamydia trachomatis Native Major Outer Membrane Protein Induces Partial Protection in Nonhuman Primates: Implication for a Trachoma Transmission-Blocking Vaccine. J. Immunol. 2009, 182, 8063–8070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, A.; Cruz-Fisher, M.I.; Cheng, C.; Pal, S.; Sun, G.; Ralli-Jain, P.; Molina, D.M.; Felgner, P.L.; Liang, X.; de la Maza, L.M. Proteomic identification of immunodominant chlamydial antigens in a mouse model. J. Proteom. 2012, 77, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Tifrea, D.F.; Pal, S.; de la Maza, L.M. A Recombinant Chlamydia trachomatis MOMP Vaccine Elicits Cross-serogroup Protection in Mice Against Vaginal Shedding and Infertility. J. Infect. Dis. 2020, 221, 191–200. [Google Scholar] [CrossRef]
- Carmichael, J.R.; Pal, S.; Tifrea, D.; de la Maza, L.M. Induction of protection against vaginal shedding and infertility by a recombinant Chlamydia vaccine. Vaccine 2011, 29, 5276–5283. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.; Juel, H.B.; Bang, P.; Cheeseman, H.M.; Dohn, R.B.; Cole, T.; Kristiansen, M.P.; Korsholm, K.S.; Lewis, D.; Olsen, A.W.; et al. Safety and immunogenicity of the chlamydia vaccine candidate CTH522 adjuvanted with CAF01 liposomes or aluminium hydroxide: A first-in-human, randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Infect. Dis. 2019, 19, 1091–1100. [Google Scholar] [CrossRef]
- O’meara, C.P.; Armitage, C.W.; Harvie, M.C.G.; Timms, P.; Lycke, N.Y.; Beagley, K.W. Immunization with a MOMP-Based Vaccine Protects Mice against a Pulmonary Chlamydia Challenge and Identifies a Disconnection between Infection and Pathology. PLoS ONE 2013, 8, e61962. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, L.; Demick, K.P.; Petersen, J.W.; Polka, M.; Rudersdorf, R.A.; Van Der Pol, B.; Jones, R.; Angevine, M.; Demars, R. Chlamydia trachomatis major outer membrane protein (MOMP) epitopes that activate HLA class II-restricted T cells from infected humans. J. Immunol. 1996, 157, 4554–4567. [Google Scholar] [CrossRef] [PubMed]
- Baehr, W.; Zhang, Y.X.; Joseph, T.; Su, H.; Nano, F.E.; Everett, K.D.; Caldwell, H.D. Mapping antigenic domains expressed by Chlamydia trachomatis major outer membrane protein genes. Proc. Natl. Acad. Sci. USA 1988, 85, 4000–4004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitch, W.M.; Peterson, E.M.; De La Maza, L.M. Phylogenetic analysis of the outer-membrane-protein genes of Chlamydiae, and its implication for vaccine development. Mol. Biol. Evol. 1993, 10, 892–913. [Google Scholar]
- Plotkin, S.A.; Orenstein, W.A.; Offit, P.A. Plotkin’s Vaccines, 7th ed.; Elsevier: Philadelphia, PA, USA, 2018; Volume xxi, 1691p. [Google Scholar]
- Morrison, S.G.; Morrison, R.P. A Predominant Role for Antibody in Acquired Immunity to Chlamydial Genital Tract Reinfection. J. Immunol. 2005, 175, 7536–7542. [Google Scholar] [CrossRef] [Green Version]
- Morrison, R.P.; Feilzer, K.; Tumas, D.B. Gene knockout mice establish a primary protective role for major histocompatibility complex class II-restricted responses in Chlamydia trachomatis genital tract infection. Infect. Immun. 1995, 63, 4661–4668. [Google Scholar] [CrossRef] [Green Version]
- Farris, C.M.; Morrison, S.G.; Morrison, R.P. CD4+T Cells and Antibody Are Required for Optimal Major Outer Membrane Protein Vaccine-Induced Immunity to Chlamydia muridarum Genital Infection. Infect. Immun. 2010, 78, 4374–4383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, H. The immunogenicity of CpG-antigen conjugates. Adv. Drug Deliv. Rev. 2009, 61, 243–247. [Google Scholar] [CrossRef]
- Krieg, A.M. Immune effects and mechanisms of action of CpG motifs. Vaccine 2000, 19, 618–622. [Google Scholar] [CrossRef]
- Pal, S.; Davis, H.L.; Peterson, E.M.; de la Maza, L.M. Immunization with the Chlamydia trachomatis Mouse Pneumonitis Major Outer Membrane Protein by Use of CpG Oligodeoxynucleotides as an Adjuvant Induces a Protective Immune Response against an Intranasal Chlamydial Challenge. Infect. Immun. 2002, 70, 4812–4817. [Google Scholar] [CrossRef] [Green Version]
- Tifrea, D.F.; Pal, S.; le Bon, C.; Cocco, M.J.; Zoonens, M.; de la Maza, L.M. Improved protection against Chlamydia muridarum using the native major outer membrane protein trapped in Resiquimod-carrying amphipols and effects in protection with addition of a Th1 (CpG-1826) and a Th2 (Montanide ISA 720) adjuvant. Vaccine 2020, 38, 4412–4422. [Google Scholar] [CrossRef]
- Girndt, M.; Pluer, M.; Dellanna, F.; Michelsen, A.K.; Beige, J.; Toussaint, K.; Wehweck, H.J.; Koch, M.; Rachti, S.H.; Faust, J.; et al. Immunogenicity and safety of a booster dose of the hepatitis B vaccine HepB-CpG (HEPLISAV-B(R)) compared with HepB-Eng (Engerix-B(R)) and HepB-AS04 (Fendrix(R)) in adults receiving hemodialysis who previously received hepatitis B vaccination and are not seroprotected: Results of a randomized, multicenter phase 3 study. Hum. Vaccines Immunother. 2022, 18, 2136912. [Google Scholar]
- Miles, A.P.; McClellan, H.A.; Rausch, K.M.; Zhu, D.; Whitmore, M.D.; Singh, S.; Martin, L.B.; Wu, Y.; Giersing, B.K.; Stowers, A.W.; et al. Montanide ISA 720 vaccines: Quality control of emulsions, stability of formulated antigens, and comparative immunogenicity of vaccine formulations. Vaccine 2005, 23, 2530–2539. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, E.J.; Femel, J.; Andersson, K.; Bjorkelund, H.; Hellman, L.; Olsson, A.K. The non-toxic and biodegradable adjuvant Montanide ISA 720/CpG can replace Freund’s in a cancer vaccine targeting ED-B—A prerequisite for clinical development. Vaccine 2012, 30, 225–230. [Google Scholar] [CrossRef]
- Toledo, H.; Baly, A.; Castro, O.; Resik, S.; Laferté, J.; Rolo, F.; Navea, L.; Lobaina, L.; Cruz, O.; Mıíguez, J.; et al. A phase I clinical trial of a multi-epitope polypeptide TAB9 combined with Montanide ISA 720 adjuvant in non-HIV-1 infected human volunteers. Vaccine 2001, 19, 4328–4336. [Google Scholar] [CrossRef] [PubMed]
- Saul, A.; Lawrence, G.; Allworth, A.; Elliott, S.; Anderson, K.; Rzepczyk, C.; Martin, L.B.; Taylor, D.; Eisen, D.P.; Irving, D.O.; et al. A human phase 1 vaccine clinical trial of the Plasmodium falciparum malaria vaccine candidate apical membrane antigen 1 in Montanide ISA720 adjuvant. Vaccine 2005, 23, 3076–3083. [Google Scholar] [CrossRef]
- Feitsma, E.A.; Janssen, Y.F.; Boersma, H.H.; van Sleen, Y.; van Baarle, D.; Alleva, D.G.; Lancaster, T.M.; Sathiyaseelan, T.; Murikipudi, S.; Delpero, A.R.; et al. A randomized phase I/II safety and immunogenicity study of the Montanide-adjuvanted SARS-CoV-2 spike protein-RBD-Fc vaccine, AKS-452. Vaccine 2023, 41, 2184–2197. [Google Scholar] [CrossRef]
- Ross, T.M.; Gokanapudi, N.; Ge, P.; Shi, H.; Richardson, R.A.; Pierce, S.R.; Sanchez, P.; Ullah, S.; De Luca, E.; Sautto, G.A.; et al. Kinetic of the Antibody Response Following AddaVax-Adjuvanted Immunization with Recombinant Influenza Antigens. Vaccines 2022, 10, 1315. [Google Scholar] [CrossRef]
- Hernandez-Davies, J.E.; Dollinger, E.P.; Pone, E.J.; Felgner, J.; Liang, L.; Strohmeier, S.; Jan, S.; Albin, T.J.; Jain, A.; Nakajima, R.; et al. Magnitude and breadth of antibody cross-reactivity induced by recombinant influenza hemagglutinin trimer vaccine is enhanced by combination adjuvants. Sci. Rep. 2022, 12, 9198. [Google Scholar] [CrossRef] [PubMed]
- Pouliot, K.; Buglione-Corbett, R.; Marty-Roix, R.; Montminy-Paquette, S.; West, K.; Wang, S.; Lu, S.; Lien, E. Contribution of TLR4 and MyD88 for adjuvant monophosphoryl lipid A (MPLA) activity in a DNA prime–protein boost HIV-1 vaccine. Vaccine 2014, 32, 5049–5056. [Google Scholar] [CrossRef]
- Weeratna, R.D.; Makinen, S.R.; McCluskie, M.J.; Davis, H.L. TLR agonists as vaccine adjuvants: Comparison of CpG ODN and Resiquimod (R-848). Vaccine 2005, 23, 5263–5270. [Google Scholar] [CrossRef]
- Pulendran, B.; Arunachalam, P.S.; O’hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Chearwae, W.; Castellino, F.; Manischewitz, J.; King, L.R.; Honorkiewicz, A.; Rock, M.T.; Edwards, K.M.; Del Giudice, G.; Rappuoli, R.; et al. Vaccines with MF59 Adjuvant Expand the Antibody Repertoire to Target Protective Sites of Pandemic Avian H5N1 Influenza Virus. Sci. Transl. Med. 2010, 2, 15ra5. [Google Scholar] [CrossRef] [PubMed]
- Galson, J.D.; Trück, J.; Kelly, D.F.; van der Most, R. Investigating the effect of AS03 adjuvant on the plasma cell repertoire following pH1N1 influenza vaccination. Sci. Rep. 2016, 6, 37229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerschlag, M.R. Chlamydial and Gonococcal Infections in Infants and Children. Clin. Infect. Dis. 2011, 53 (Suppl. S3), S99–S102. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L.; Aronson, M.D.; Schachter, J. Chlamydia trachomatis infection in adults with community-acquired pneumonia. JAMA 1981, 245, 1319–1322. [Google Scholar] [CrossRef]
- Tack, K.J.; Peterson, P.K.; Rasp, F.L.; O’Leary, M.; Hanto, D.; Simmons, R.L.; Sabath, L.D. Isolation of Chlamydia Trachomatis from The Lower Respiratory Tract of Adults. Lancet 1980, 1, 116–120. [Google Scholar] [CrossRef]
- Grayston, J.T.; Kuo, C.C.; Wang, S.P.; Altman, J. A New Chlamydia psittaci Strain, TWAR, Isolated in Acute Respiratory Tract Infections. New Engl. J. Med. 1986, 315, 161–168. [Google Scholar] [CrossRef]
- Murdin, A.D.; Dunn, P.; Sodoyer, R.; Wang, J.; Caterini, J.; Brunham, R.C.; Aujame, L.; Oomen, R. Use of a Mouse Lung Challenge Model to Identify Antigens Protective against Chlamydia pneumoniae Lung Infection. J. Infect. Dis. 2000, 181 (Suppl. S3), S544–S551. [Google Scholar] [CrossRef] [Green Version]
- Finco, O.; Bonci, A.; Agnusdei, M.; Scarselli, M.; Petracca, R.; Norais, N.; Ferrari, G.; Garaguso, I.; Donati, M.; Sambri, V.; et al. Identification of new potential vaccine candidates against Chlamydia pneumoniae by multiple screenings. Vaccine 2005, 23, 1178–1188. [Google Scholar] [CrossRef]
- Chacko, A.; Delbaz, A.; Walkden, H.; Basu, S.; Armitage, C.W.; Eindorf, T.; Trim, L.K.; Miller, E.; West, N.P.; John, J.A.S.; et al. Chlamydia pneumoniae can infect the central nervous system via the olfactory and trigeminal nerves and contributes to Alzheimer’s disease risk. Sci. Rep. 2022, 12, 2759. [Google Scholar] [CrossRef]
- Jantos, C.A.; Augustin, J.; Durchfeld-Meyer, B.; Baumgärtner, W.; Schiefer, H.G. Experimental genital tract infection with Chlamydia psittaci (GPIC agent) in male rats. Infection 1998, 26, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Knittler, M.R.; Sachse, K. Chlamydia psittaci: Update on an underestimated zoonotic agent. Pathog. Dis. 2015, 73, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.; Wen, Y.; Ran, O.; Chen, L.; Wang, C.; Li, L.; Xie, Y.; Zhang, Y.; Chen, C.; Wu, Y. Protective immunity induced by recombinant protein CPSIT_p8 of Chlamydia psittaci. Appl. Microbiol. Biotechnol. 2016, 100, 6385–6393. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, H.D.; Kromhout, J.; Schachter, J. Purification and partial characterization of the major outer membrane protein of Chlamydia trachomatis. Infect. Immun. 1981, 31, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Tifrea, D.F.; Follmann, F.; Andersen, P.; de la Maza, L.M. The cationic liposomal adjuvants CAF01 and CAF09 formulated with the major outer membrane protein elicit robust protection in mice against a Chlamydia muridarum respiratory challenge. Vaccine 2017, 35, 1705–1711. [Google Scholar] [CrossRef]
- Pal, S.; Fielder, T.J.; Peterson, E.M.; De La Maza, L.M. Protection against infertility in a BALB/c mouse salpingitis model by intranasal immunization with the mouse pneumonitis biovar of Chlamydia trachomatis. Infect. Immun. 1994, 62, 3354–3362. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Pal, S.; Tifrea, D.; Jia, Z.; de la Maza, L.M. A vaccine formulated with a combination of TLR-2 and TLR-9 adjuvants and the recombinant major outer membrane protein elicits a robust immune response and significant protection against a Chlamydia muridarum challenge. Microbes Infect. 2014, 16, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Jain, P.; Pal, S.; Tifrea, D.; Sun, G.; Teng, A.A.; Liang, X.; Felgner, P.L.; de la Maza, L.M. Assessment of the role in protection and pathogenesis of the Chlamydia muridarum V-type ATP synthase subunit A (AtpA) (TC0582). Microbes Infect. 2014, 16, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Tifrea, D.F.; Pal, S.; Popot, J.-L.; Cocco, M.J.; de la Maza, L.M. Increased Immunoaccessibility of MOMP Epitopes in a Vaccine Formulated with Amphipols May Account for the Very Robust Protection Elicited against a Vaginal Challenge with Chlamydia muridarum. J. Immunol. 2014, 192, 5201–5213. [Google Scholar] [CrossRef] [Green Version]
- Peterson, E.M.; Zhong, G.M.; Carlson, E.; De La Maza, L.M. Protective role of magnesium in the neutralization by antibodies of Chlamydia trachomatis infectivity. Infect. Immun. 1988, 56, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Bravo, J.; Peterson, E.M.; de la Maza, L.M. Protection of Wild-Type and Severe Combined Immunodeficiency Mice against an Intranasal Challenge by Passive Immunization with Monoclonal Antibodies to the Chlamydia trachomatis Mouse Pneumonitis Major Outer Membrane Protein. Infect. Immun. 2008, 76, 5581–5587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Cheng, X.; Peterson, E.M.; de la Maza, L.M. Mapping of a surface-exposed B-cell epitope to the variable sequent 3 of the major outer-membrane protein of Chlamydia trachomatis. J. Gen. Microbiol. 1993, 139, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tifrea, D.F.; Sun, G.; Pal, S.; Zardeneta, G.; Cocco, M.J.; Popot, J.-L.; de la Maza, L.M. Amphipols stabilize the Chlamydia major outer membrane protein and enhance its protective ability as a vaccine. Vaccine 2011, 29, 4623–4631. [Google Scholar] [CrossRef] [Green Version]
- Naglak, E.K.; Morrison, S.G.; Morrison, R.P. Neutrophils Are Central to Antibody-Mediated Protection against Genital Chlamydia. Infect. Immun. 2017, 85, e00409-17. [Google Scholar] [CrossRef] [Green Version]
- Igietseme, J.U.; Magee, D.M.; Williams, D.M.; Rank, R.G. Role for CD8+ T cells in antichlamydial immunity defined by Chlamydia-specific T-lymphocyte clones. Infect. Immun. 1994, 62, 5195–5197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, M.F.; Wilson, C.B.; Bevan, M.J.; Starnbach, M.N. Gamma interferon production by cytotoxic T lymphocytes is required for resolution of Chlamydia trachomatis infection. Infect. Immun. 1998, 66, 5457–5461. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Jain, P.; Bettahi, I.; Pal, S.; Tifrea, D.; de la Maza, L.M. A TLR2 agonist is a more effective adjuvant for a Chlamydia major outer membrane protein vaccine than ligands to other TLR and NOD receptors. Vaccine 2011, 29, 6641–6649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Luke, C.J.; Barbour, A.G.; Peterson, E.M.; de la Maza, L.M. Immunization with the Chlamydia trachomatis major outer membrane protein, using the outer surface protein A of Borrelia burgdorferi as an adjuvant, can induce protection against a chlamydial genital challenge. Vaccine 2003, 21, 1455–1465. [Google Scholar] [CrossRef]
- Pal, S.; Cruz-Fisher, M.I.; Cheng, C.; Carmichael, J.R.; Tifrea, D.F.; Tatarenkova, O.; de la Maza, L.M. Vaccination with the recombinant major outer membrane protein elicits long-term protection in mice against vaginal shedding and infertility following a Chlamydia muridarum genital challenge. NPJ Vaccines 2020, 5, 90. [Google Scholar] [CrossRef]
- Pal, S.; Favaroni, A.; Tifrea, D.F.; Hanisch, P.T.; Luczak, S.E.; Hegemann, J.H.; de la Maza, L.M. Comparison of the nine polymorphic membrane proteins of Chlamydia trachomatis for their ability to induce protective immune responses in mice against a C. muridarum challenge. Vaccine 2017, 35, 2543–2549. [Google Scholar] [CrossRef] [Green Version]
- Armitage, C.W.; O’Meara, C.P.; Harvie, M.C.G.; Timms, P.; Blumberg, R.S.; Beagley, K.W. Divergent outcomes following transcytosis of IgG targeting intracellular and extracellular chlamydial antigens. Immunol. Cell Biol. 2014, 92, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ausar, S.F.; Tifrea, D.F.; Cheng, C.; Gallichan, S.; Sanchez, V.; de la Maza, L.M.; Visan, L. Protection of outbred mice against a vaginal challenge by a Chlamydia trachomatis serovar E recombinant major outer membrane protein vaccine is dependent on phosphate substitution in the adjuvant. Hum. Vaccines Immunother. 2020, 16, 2537–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, C.V.; Zhang, X.; Giovannone, N.; Sennott, E.L.; Starnbach, M.N. Protective Immunity against Chlamydia trachomatis Can Engage Both CD4+ and CD8+ T Cells and Bridge the Respiratory and Genital Mucosae. J. Immunol. 2015, 194, 2319–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkey, M.R.; Nguyen, D.H.; Essilfie, A.T.; Kim, R.Y.; Hatchwell, L.M.; Collison, A.M.; Yagita, H.; Foster, P.S.; Horvat, J.C.; Mattes, J.; et al. Tumor necrosis factor-related apoptosis-inducing ligand translates neonatal respiratory infection into chronic lung disease. Mucosal Immunol. 2014, 7, 478–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Chen, Q.; Moore, J.; Kolls, J.K.; Halperin, S.; Wang, J. Critical role of the interleukin-17/interleukin-17 receptor axis in regulating host susceptibility to respiratory infection with Chlamydia species. Infect. Immun. 2009, 77, 5059–5070. [Google Scholar] [CrossRef] [Green Version]
- Tifrea, D.F.; Pal, S.; Fairman, J.; Massari, P.; de la Maza, L.M. Protection against a chlamydial respiratory challenge by a chimeric vaccine formulated with the Chlamydia muridarum major outer membrane protein variable domains using the Neisseria lactamica porin B as a scaffold. NPJ Vaccines 2020, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Cruz-Fisher, M.I.; Tifrea, D.; Pal, S.; Wizel, B.; de la Maza, L.M. Induction of protection in mice against a respiratory challenge by a vaccine formulated with the Chlamydia major outer membrane protein adjuvanted with IC31(R). Vaccine 2011, 29, 2437–2443. [Google Scholar] [CrossRef]
- Meoni, E.; Faenzi, E.; Frigimelica, E.; Zedda, L.; Skibinski, D.; Giovinazzi, S.; Bonci, A.; Petracca, R.; Bartolini, E.; Galli, G.; et al. CT043, a protective antigen that induces a CD4+ Th1 response during Chlamydia trachomatis infection in mice and humans. Infect. Immun. 2009, 77, 4168–4176. [Google Scholar] [CrossRef] [Green Version]
- Finco, O.; Frigimelica, E.; Buricchi, F.; Petracca, R.; Galli, G.; Faenzi, E.; Meoni, E.; Bonci, A.; Agnusdei, M.; Nardelli, F.; et al. Approach to discover T- and B-cell antigens of intracellular pathogens applied to the design of Chlamydia trachomatis vaccines. Proc. Natl. Acad. Sci. USA 2011, 108, 9969–9974. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Mirzakhanyan, Y.; Gershon, P.; Tifrea, D.F.; de la Maza, L.M. Induction of protection in mice against a respiratory challenge by a vaccine formulated with exosomes isolated from Chlamydia muridarum infected cells. NPJ Vaccines 2020, 5, 87. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Collignon, C.; Bourguignon, P.; Wouters, S.; Fierens, K.; Fochesato, M.; Dendouga, N.; Langlet, C.; Malissen, B.; Lambrecht, B.N.; et al. Enhancement of Adaptive Immunity by the Human Vaccine Adjuvant AS01 Depends on Activated Dendritic Cells. J. Immunol. 2014, 193, 1920–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, S.; Slepenkin, A.; Felgner, J.; Huw Davies, D.; Felgner, P.; de la Maza, L.M. Evaluation of Four Adjuvant Combinations, IVAX-1, IVAX-2, CpG-1826+Montanide ISA 720 VG and CpG-1018+Montanide ISA 720 VG, for Safety and for Their Ability to Elicit Protective Immune Responses in Mice against a Respiratory Challenge with Chlamydia muridarum. Pathogens 2023, 12, 863. https://doi.org/10.3390/pathogens12070863
Pal S, Slepenkin A, Felgner J, Huw Davies D, Felgner P, de la Maza LM. Evaluation of Four Adjuvant Combinations, IVAX-1, IVAX-2, CpG-1826+Montanide ISA 720 VG and CpG-1018+Montanide ISA 720 VG, for Safety and for Their Ability to Elicit Protective Immune Responses in Mice against a Respiratory Challenge with Chlamydia muridarum. Pathogens. 2023; 12(7):863. https://doi.org/10.3390/pathogens12070863
Chicago/Turabian StylePal, Sukumar, Anatoli Slepenkin, Jiin Felgner, D. Huw Davies, Philip Felgner, and Luis M. de la Maza. 2023. "Evaluation of Four Adjuvant Combinations, IVAX-1, IVAX-2, CpG-1826+Montanide ISA 720 VG and CpG-1018+Montanide ISA 720 VG, for Safety and for Their Ability to Elicit Protective Immune Responses in Mice against a Respiratory Challenge with Chlamydia muridarum" Pathogens 12, no. 7: 863. https://doi.org/10.3390/pathogens12070863