
Design and Optimization of a Monkeypox virus Specific Serological Assay

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. ParticipantSelection
2.1.1. 2003 United States Mpox Outbreak Participants
2.1.2. VARV Outbreak Participants
2.1.3. Vaccine Study Participants
2.1.4. DRC Surveillance Participants
2.1.5. Ghanian Study Participants
2.2. IRB
2.3. Peptide Design and Synthesis
2.4. OPXV ELISA
2.5. MPXV Peptide ELISA Protocol and Optimization
2.6. Statistical Analysis
3. Results
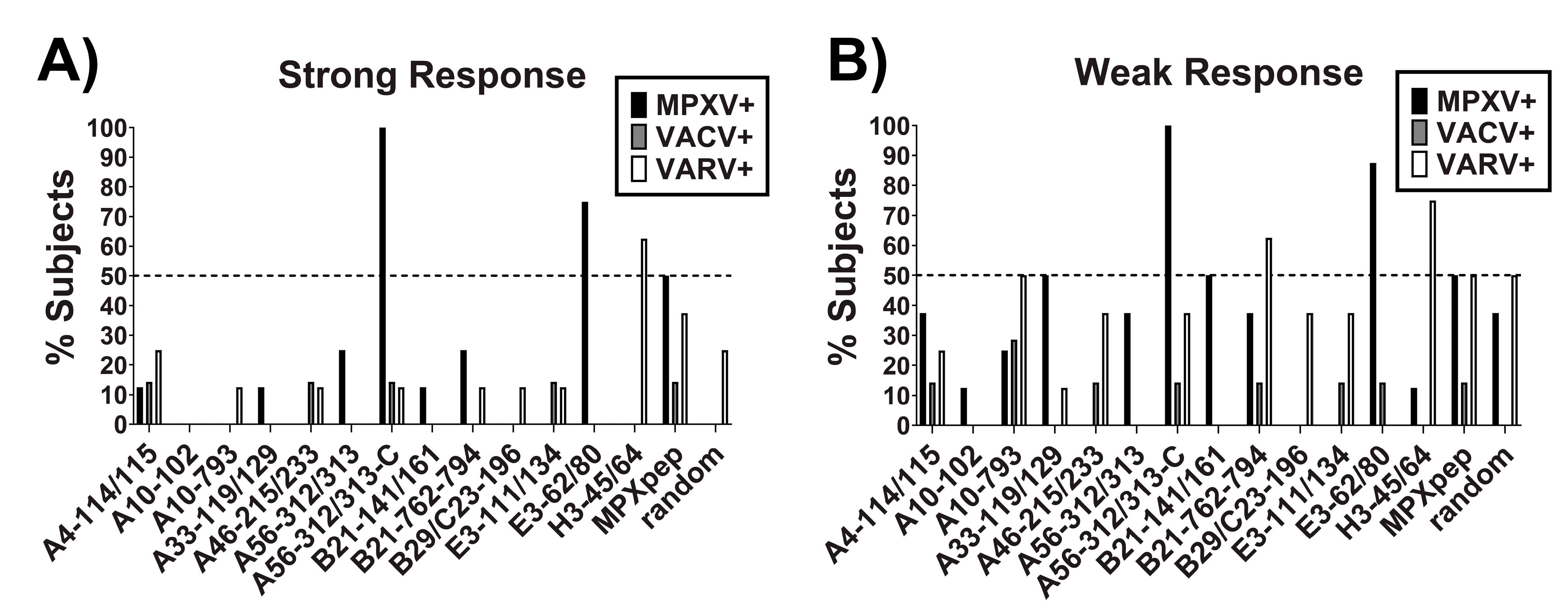
3.1. Design and Validation of MPXV Specific Peptides
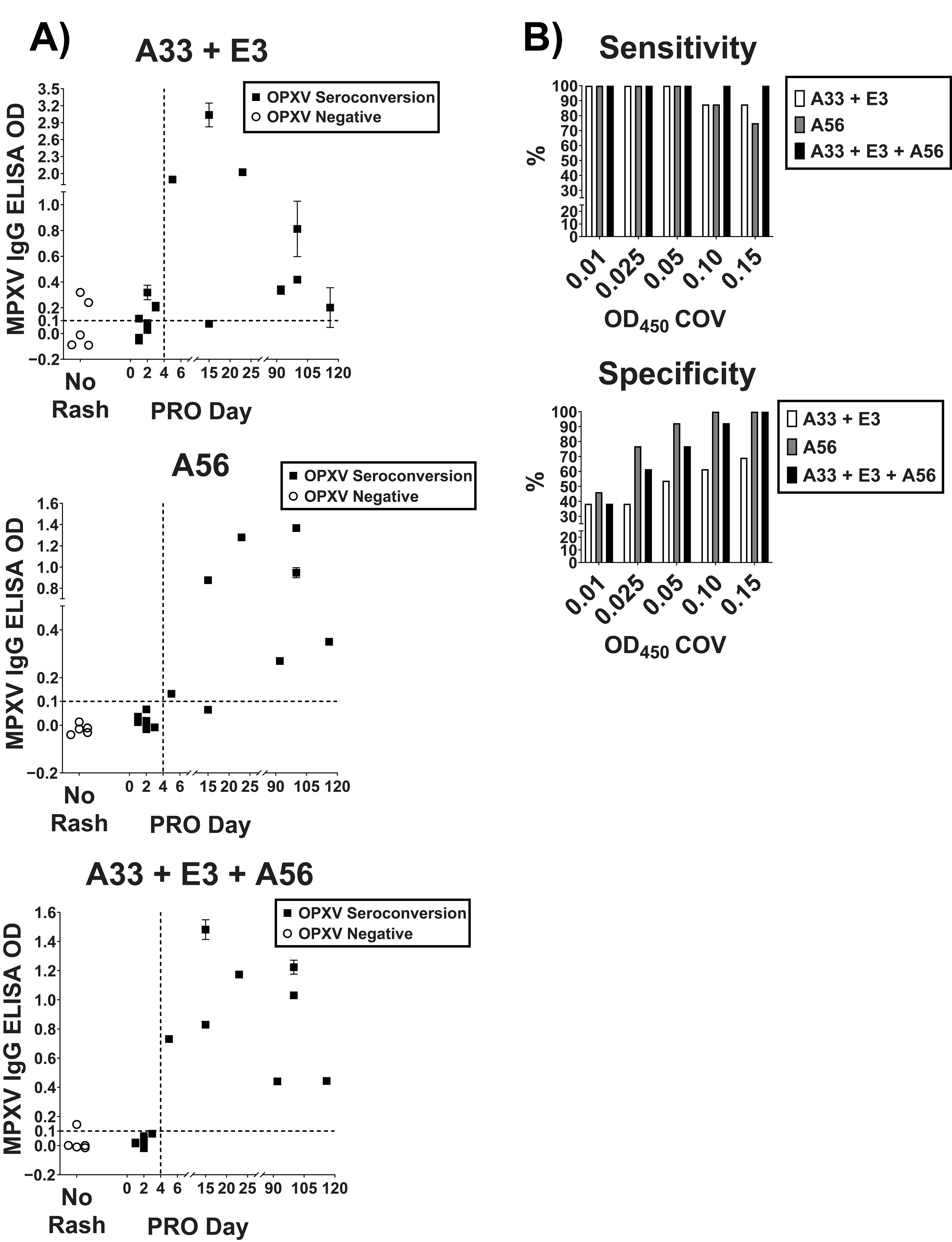
3.2. Validation and Optimization of MPXV Specific Peptide-Based ELISA
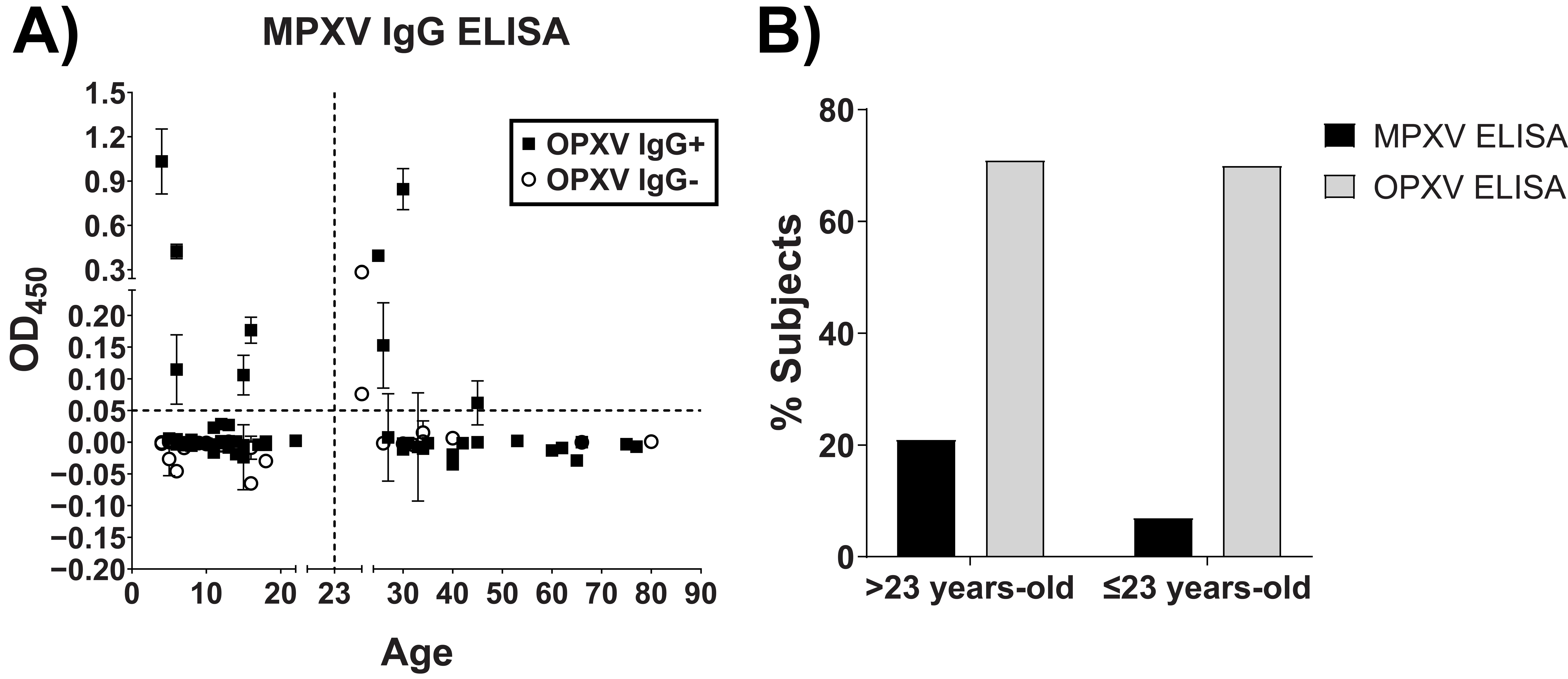
3.3. Retrospective Analysis of a Population with Unknown MPXV Exposure
4. Discussion
5. Disclaimer
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ladnyj, I.D.; Ziegler, P.; Kima, E. A human infection caused by monkeypox virus in Basankusu Territory, Democratic Republic of the Congo. Bull. World Health Organ. 1972, 46, 593–597. [Google Scholar] [PubMed]
- Di Giulio, D.B.; Eckburg, P.B. Human monkeypox: An emerging zoonosis. Lancet Infect. Dis. 2004, 4, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Rimoin, A.W.; Mulembakani, P.M.; Johnston, S.C.; Lloyd Smith, J.O.; Kisalu, N.K.; Kinkela, T.L.; Blumberg, S.; Thomassen, H.A.; Pike, B.L.; Fair, J.N.; et al. Major increase in human monkeypox incidence 30 years after smallpox vaccination campaigns cease in the Democratic Republic of Congo. Proc. Natl. Acad. Sci. USA 2010, 107, 16262–16267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Multistate outbreak of monkeypox--Illinois, Indiana, and Wisconsin, 2003. MMWR Morb. Mortal. Wkly. Rep. 2003, 52, 537–540. [Google Scholar]
- Reed, K.D.; Melski, J.W.; Graham, M.B.; Regnery, R.L.; Sotir, M.J.; Wegner, M.V.; Kazmierczak, J.J.; Stratman, E.J.; Li, Y.; Fairley, J.A.; et al. The detection of monkeypox in humans in the Western Hemisphere. N. Engl. J. Med. 2004, 350, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, J.P.; Barkati, S.; Walmsley, S.; Rockstroh, J.; Antinori, A.; Harrison, L.B.; Palich, R.; Nori, A.; Reeves, I.; Habibi, M.S.; et al. Monkeypox Virus Infection in Humans across 16 Countries—April-June 2022. N. Engl. J. Med. 2022, 387, 679–691. [Google Scholar] [CrossRef]
- CDC. 2022 Monkeypox Outbreak Global Map. Available online: https://www.cdc.gov/poxvirus/monkeypox/response/2022/world-map.html (accessed on 9 June 2022).
- WHO. 2022 Monkeypox Outbreak: Global Trends. Available online: https://worldhealthorg.shinyapps.io/mpx_global/ (accessed on 9 June 2022).
- Minhaj, F.S.; Ogale, Y.P.; Whitehill, F.; Schultz, J.; Foote, M.; Davidson, W.; Hughes, C.M.; Wilkins, K.; Bachmann, L.; Chatelain, R.; et al. Monkeypox Outbreak—Nine States, May 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 764–769. [Google Scholar] [CrossRef]
- Nolen, L.D.; Osadebe, L.; Katomba, J.; Likofata, J.; Mukadi, D.; Monroe, B.; Doty, J.; Hughes, C.M.; Kabamba, J.; Malekani, J.; et al. Extended Human-To-Human Transmission during a Monkeypox Outbreak in the Democratic Republic of the Congo. Emerg. Infect. Dis. 2016, 22, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Philpott, D.; Hughes, C.M.; Alroy, K.A.; Kerins, J.L.; Pavlick, J.; Asbel, L.; Crawley, A.; Newman, A.P.; Spencer, H.; Feldpausch, A.; et al. Epidemiologic and Clinical Characteristics of Monkeypox Cases—United States, 17 May–22 July 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 1018–1022. [Google Scholar] [CrossRef]
- Antinori, A.; Mazzotta, V.; Vita, S.; Carletti, F.; Tacconi, D.; Lapini, L.E.; D’Abramo, A.; Cicalini, S.; Lapa, D.; Pittalis, S.; et al. Epidemiological, clinical and virological characteristics of four cases of monkeypox support transmission through sexual contact, Italy, May 2022. Euro Surveill. 2022, 27, 2200421. [Google Scholar] [CrossRef]
- Vusirikala, A.; Charles, H.; Balasegaram, S.; Macdonald, N.; Kumar, D.; Barker-Burnside, C.; Cumiskey, K.; Dickinson, M.; Watson, M.; Olufon, O.; et al. Epidemiology of Early Monkeypox Virus Transmission in Sexual Networks of Gay and Bisexual Men, England, 2022. Emerg. Infect. Dis. 2022, 28, 2082–2086. [Google Scholar] [CrossRef]
- Fine, P.E.; Jezek, Z.; Grab, B.; Dixon, H. The transmission potential of monkeypox virus in human populations. Int. J. Epidemiol. 1988, 17, 643–650. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Damon, I.K. Outbreaks of human monkeypox after cessation of smallpox vaccination. Trends Microbiol. 2012, 20, 80–87. [Google Scholar] [CrossRef]
- Karem, K.L.; Reynolds, M.; Braden, Z.; Lou, G.; Bernard, N.; Patton, J.; Damon, I.K. characterization of acute-phase humoral immunity to monkeypox: Use of immunoglobulin M enzyme-linked immunosorbent assay for detection of monkeypox infection during the 2003 North American outbreak. Clin. Diagn. Lab. Immunol. 2005, 12, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.E.; Hammarlund, E.; Slifka, M.K. Optimization of peptide-based ELISA for serological diagnostics: A retrospective study of human monkeypox infection. Vector Borne Zoonotic Dis. 2012, 12, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.E.; Slifka, M.K. Retrospective analysis of monkeypox infection. Emerg. Infect. Dis. 2008, 14, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Shchelkunov, S.N.; Totmenin, A.V.; Babkin, I.V.; Safronov, P.F.; Ryazankina, O.I.; Petrov, N.A.; Gutorov, V.V.; Uvarova, E.A.; Mikheev, M.V.; Sisler, J.R.; et al. Human monkeypox and smallpox viruses: Genomic comparison. FEBS Lett. 2001, 509, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gispen, R.; Brand-Saathof, B.B.; Hekker, A.C. Monkeypox-specific antibodies in human and simian sera from the Ivory Coast and Nigeria. Bull. World Health Organ. 1976, 53, 355–360. [Google Scholar] [PubMed]
- Jezek, Z.; Nakano, J.H.; Arita, I.; Mutombo, M.; Szczeniowski, M.; Dunn, C. Serological survey for human monkeypox infections in a selected population in Zaire. J. Trop. Med. Hyg. 1987, 90, 31–38. [Google Scholar]
- Marennikova, S.S.; Malceva, N.N.; Habahpaseva, N.A. ELISA--a simple test for detecting and differentiating antibodies to closely related orthopoxviruses. Bull. World Health Organ. 1981, 59, 365–369. [Google Scholar]
- Davies, D.H.; McCausland, M.M.; Valdez, C.; Huynh, D.; Hernandez, J.E.; Mu, Y.; Hirst, S.; Villarreal, L.; Felgner, P.L.; Crotty, S. Vaccinia virus H3L envelope protein is a major target of neutralizing antibodies in humans and elicits protection against lethal challenge in mice. J. Virol. 2005, 79, 11724–11733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogg, C.N.; Americo, J.L.; Earl, P.L.; Resch, W.; Aldaz-Carroll, L.; Eisenberg, R.J.; Cohen, G.H.; Moss, B. Disparity between levels of in vitro neutralization of vaccinia virus by antibody to the A27 protein and protection of mice against intranasal challenge. J. Virol. 2008, 82, 8022–8029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heraud, J.M.; Edghill-Smith, Y.; Ayala, V.; Kalisz, I.; Parrino, J.; Kalyanaraman, V.S.; Manischewitz, J.; King, L.R.; Hryniewicz, A.; Trindade, C.J.; et al. Subunit recombinant vaccine protects against monkeypox. J. Immunol. 2006, 177, 2552–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, J.W.; Ferro, A.M.; Golden, J.W.; Silvera, P.; Dudek, J.; Alterson, K.; Custer, M.; Rivers, B.; Morris, J.; Owens, G.; et al. Molecular smallpox vaccine delivered by alphavirus replicons elicits protective immunity in mice and non-human primates. Vaccine 2009, 28, 494–511. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, D.R.; Goudsmit, J.; Holterman, L.; Ewald, B.A.; Denholtz, M.; Devoy, C.; Giri, A.; Grandpre, L.E.; Heraud, J.M.; Franchini, G.; et al. Differential antigen requirements for protection against systemic and intranasal vaccinia virus challenges in mice. J. Virol. 2008, 82, 6829–6837. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Meng, X.; Yan, B.; Crotty, S.; Deng, J.; Xiang, Y. An epitope conserved in orthopoxvirus A13 envelope protein is the target of neutralizing and protective antibodies. Virology 2011, 418, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.H.; Molina, D.M.; Wrammert, J.; Miller, J.; Hirst, S.; Mu, Y.; Pablo, J.; Unal, B.; Nakajima-Sasaki, R.; Liang, X.; et al. Proteome-wide analysis of the serological response to vaccinia and smallpox. Proteomics 2007, 7, 1678–1686. [Google Scholar] [CrossRef]
- Hughes, L.J.; Goldstein, J.; Pohl, J.; Hooper, J.W.; Lee Pitts, R.; Townsend, M.B.; Bagarozzi, D.; Damon, I.K.; Karem, K.L. A highly specific monoclonal antibody against monkeypox virus detects the heparin binding domain of A27. Virology 2014, 464–465, 264–273. [Google Scholar] [CrossRef]
- Pittman, P.R.; Martin, J.W.; Kingebeni, P.M.; Tamfum, J.-J.M.; Wan, Q.; Reynolds, M.G.; Quinn, X.; Norris, S.; Townsend, M.B.; Satheshkumar, P.S.; et al. Clinical characterization of human monkeypox infections in the Democratic Republic of the Congo. medRxiv 2022. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Carroll, D.S.; Olson, V.A.; Hughes, C.; Galley, J.; Likos, A.; Montgomery, J.M.; Suu-Ire, R.; Kwasi, M.O.; Jeffrey Root, J.; et al. A silent enzootic of an orthopoxvirus in Ghana, West Africa: Evidence for multi-species involvement in the absence of widespread human disease. Am. J. Trop. Med. Hyg. 2010, 82, 746–754. [Google Scholar] [CrossRef] [Green Version]
- 45 CFR 46; Common Rule. HHS: Washington, DC, USA, 2018.
- 21 CFR 56; Institutional Review Boards. HHS: Washington, DC, USA, 2017.
- 42 U.S.C. §241(d); The Public Health And Welfare. United States Code: Washington, DC, USA, 2012.
- 5 U.S.C. §552a; Government Organization And Employees. United States Code: Washington, DC, USA, 2015.
- 44 U.S.C. §3501; Public Printing And Documents. United States Code: Washington, DC, USA, 2012.
- Townsend, M.B.; Keckler, M.S.; Patel, N.; Davies, D.H.; Felgner, P.; Damon, I.K.; Karem, K.L. Humoral immunity to smallpox vaccines and monkeypox virus challenge: Proteomic assessment and clinical correlations. J. Virol. 2013, 87, 900–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, E.M. Validation of laboratory-developed molecular assays for infectious diseases. Clin. Microbiol. Rev. 2010, 23, 550–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisht, H.; Weisberg, A.S.; Moss, B. Vaccinia virus l1 protein is required for cell entry and membrane fusion. J. Virol. 2008, 82, 8687–8694. [Google Scholar] [CrossRef] [Green Version]
- Hammarlund, E.; Lewis, M.W.; Carter, S.V.; Amanna, I.; Hansen, S.G.; Strelow, L.I.; Wong, S.W.; Yoshihara, P.; Hanifin, J.M.; Slifka, M.K. Multiple diagnostic techniques identify previously vaccinated individuals with protective immunity against monkeypox. Nat. Med. 2005, 11, 1005–1011. [Google Scholar] [CrossRef]
- Chen, N.; Li, G.; Liszewski, M.K.; Atkinson, J.P.; Jahrling, P.B.; Feng, Z.; Schriewer, J.; Buck, C.; Wang, C.; Lefkowitz, E.J.; et al. Virulence differences between monkeypox virus isolates from West Africa and the Congo basin. Virology 2005, 340, 46–63. [Google Scholar] [CrossRef] [Green Version]
- Likos, A.M.; Sammons, S.A.; Olson, V.A.; Frace, A.M.; Li, Y.; Olsen-Rasmussen, M.; Davidson, W.; Galloway, R.; Khristova, M.L.; Reynolds, M.G.; et al. A tale of two clades: Monkeypox viruses. J. Gen. Virol. 2005, 86, 2661–2672. [Google Scholar] [CrossRef]
- Senkevich, T.G.; Yutin, N.; Wolf, Y.I.; Koonin, E.V.; Moss, B. Ancient Gene Capture and Recent Gene Loss Shape the Evolution of Orthopoxvirus-Host Interaction Genes. mBio 2021, 12, e0149521. [Google Scholar] [CrossRef]
- Langedijk, J.P.; Brandenburg, A.H.; Middel, W.G.; Osterhaus, A.; Meloen, R.H.; van Oirschot, J.T. A subtype-specific peptide-based enzyme immunoassay for detection of antibodies to the G protein of human respiratory syncytial virus is more sensitive than routine serological tests. J. Clin. Microbiol. 1997, 35, 1656–1660. [Google Scholar] [CrossRef] [Green Version]
- Morre, S.A.; Munk, C.; Persson, K.; Kruger-Kjaer, S.; van Dijk, R.; Meijer, C.J.; van Den Brule, A.J. Comparison of three commercially available peptide-based immunoglobulin G (IgG) and IgA assays to microimmunofluorescence assay for detection of Chlamydia trachomatis antibodies. J. Clin. Microbiol. 2002, 40, 584–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakowska, D.; Colon, I.; Remington, J.S.; Grigg, M.; Golab, E.; Wilczynski, J.; Sibley, L.D. Genotyping of Toxoplasma gondii by multiplex PCR and peptide-based serological testing of samples from infants in Poland diagnosed with congenital toxoplasmosis. J. Clin. Microbiol. 2006, 44, 1382–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.T.; Steere, A.C.; Marques, A.R.; Johnson, B.J.; Miller, J.N.; Philipp, M.T. Sensitive and specific serodiagnosis of Lyme disease by enzyme-linked immunosorbent assay with a peptide based on an immunodominant conserved region of Borrelia burgdorferi vlsE. J. Clin. Microbiol. 1999, 37, 3990–3996. [Google Scholar] [CrossRef] [Green Version]
- Oladepo, D.K.; Klapper, P.E.; Marsden, H.S. Peptide based enzyme-linked immunoassays for detection of anti-HSV-2 IgG in human sera. J. Virol. Methods 2000, 87, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Lee, M.K.; Kim, S.Y.; Jang, S.Y.; Hahm, K.S. The use of multiple antigenic peptide (MAP) in the immunodiagnosis of human immunodeficiency virus infection. Biochem. Mol. Biol. Int. 1997, 43, 713–721. [Google Scholar] [CrossRef]
- Shchelkunov, S.N.; Totmenin, A.V.; Safronov, P.F.; Mikheev, M.V.; Gutorov, V.V.; Ryazankina, O.I.; Petrov, N.A.; Babkin, I.V.; Uvarova, E.A.; Sandakhchiev, L.S.; et al. Analysis of the monkeypox virus genome. Virology 2002, 297, 172–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Baxby, D.; Bennett, M.; Getty, B. Human cowpox 1969-93: A review based on 54 cases. Br. J. Dermatol. 1994, 131, 598–607. [Google Scholar] [CrossRef]
- Pelkonen, P.M.; Tarvainen, K.; Hynninen, A.; Kallio, E.R.; Henttonen, K.; Palva, A.; Vaheri, A.; Vapalahti, O. Cowpox with severe generalized eruption, Finland. Emerg. Infect. Dis. 2003, 9, 1458–1461. [Google Scholar] [CrossRef]
- McCollum, A.M.; Damon, I.K. Human monkeypox. Clin. Infect. Dis. 2014, 58, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Amanna, I.J.; Carlson, N.E.; Slifka, M.K. Duration of humoral immunity to common viral and vaccine antigens. N. Engl. J. Med. 2007, 357, 1903–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotty, S.; Felgner, P.; Davies, H.; Glidewell, J.; Villarreal, L.; Ahmed, R. Cutting edge: Long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003, 171, 4969–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammarlund, E.; Lewis, M.W.; Hansen, S.G.; Strelow, L.I.; Nelson, J.A.; Sexton, G.J.; Hanifin, J.M.; Slifka, M.K. Duration of antiviral immunity after smallpox vaccination. Nat. Med. 2003, 9, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Putz, M.M.; Alberini, I.; Midgley, C.M.; Manini, I.; Montomoli, E.; Smith, G.L. Prevalence of antibodies to Vaccinia virus after smallpox vaccination in Italy. J. Gen. Virol. 2005, 86 Pt 11, 2955–2960. [Google Scholar] [CrossRef]
- Taub, D.D.; Ershler, W.B.; Janowski, M.; Artz, A.; Key, M.L.; McKelvey, J.; Muller, D.; Moss, B.; Ferrucci, L.; Duffey, P.L.; et al. Immunity from smallpox vaccine persists for decades: A longitudinal study. Am. J. Med. 2008, 121, 1058–1064. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OPXV Strain | Place of Isolation | Year | Clinical Isolate | Host | Accession Number |
|---|---|---|---|---|---|
| CPXV-AUS1999-867 | Texing, Austria | 1999 | Local lesions | Cat | HQ407377 |
| CPXV-BR | Brighton, United Kingdom | 1937 | Local lesions | Human | NC_003663 |
| CPXV-GER1980 | Hameln, Germany | 1980 | Local lesions | Elephant | HQ420895 |
| CPXV-GER1998-2 | Eckental, Germany | 1998 | Local lesions | Human | HQ420897 |
| CPXV-GER2002-MKY | Göttingen, Germany | 2002 | Fatal generalization | Marmoset | HQ420898 |
| CPXV-GER1991 | Munich, Germany | 1991 | Local lesions | Human | DQ437593 |
| CPXV-GRI | Moscow, Russia | 1990 | Local lesions | Human | X 94355 |
| CPXV-GER1990 | Bonn, Germany | 1990 | Fatal generalization | Human | HQ420896 |
| MPXV-COG2003-358 | Impfondo, Republic of the Congo | 2003 | Local lesions | Human | DQ011154; |
| MPXV-COP58 | Copenhagen, Denmark | 1958 | Local lesions | Monkey | AY753185 |
| MPXV-USA2003-039 | Wisconsin, USA | 2003 | Local lesions | Human | DQ011157 |
| MPXV-WR267 | Washington, DC, USA | 1962 | Local lesions | Monkey | AY603973 |
| MPXV-ZAR | Kasai Oriental, Zaire | 1996 | Local lesions | Human | NC_003310 |
| MPXV-ZAR-1979-005 | Equateur, Zaire | 1978 | Local lesions | Human | DQ011155 |
| VACV-ACAM2000 | New York, USA | 2004 | Cell culture | Cow (Dryvax) | AY313847 |
| VACV-COP | New York, USA | 1913 | Cell culture | Cow (Dryvax) | M35027 |
| VACV-MVA | Munich, Germany | 1971 | Chicken embryo fibroblasts culture | - | DQ983236 |
| VARV-BEN68 | Benin | 1968 | Local lesions | Human | DQ441416 |
| VARV-BGD-Banu | Bangladesh | 1975 | Local lesions | Human | DQ437581 |
| VARV-JPV51-hrpr | Japan | 1951 | Local lesions | Human | DQ441430 |
| Peptide | Protein | Sequence | Substitution(s) | Length |
|---|---|---|---|---|
| A4-114/115 | A4 (OPG130) | Biotin-β-Ala-PTPAILLPSSTAPVLKPRQQTNT | T114V, P115L | 23 |
| A10-102 | A10 (OPG136) | Biotin-β-Ala-NAGNIDIINHPINISSETNPIIN | T102H | 23 |
| A10-793 | A10 (OPG136) | Biotin-β-Ala-TIERIFNAKVCDDVKASMLEKY | G793C | 22 |
| A33-119/129 | A33 (OPG161) | Biotin-β-Ala-GSCYILHSDYKSFEDAKANCAAESS | Q119K, L120S, S122E, T129A | 25 |
| A46-215/223 | A46 (OPG176) | Biotin-β-Ala-LRGHTDSIEDEFDHFEDDDSST | E215D, Y223H | 22 |
| A56-312/313 | A56 (OPG185) | Biotin-β-Ala-SAVAIFCITYYICNKHPRKYKTENKV | R312H, S313P | 26 |
| A56-312/313-C * | A56 (OPG185) | Biotin-β-Ala-SAVAIFCITYYICNKHPRKYKTENKV | R312H, S313P | 26 |
| B21-141/161 | B21 (OPG211) | Biotin-β-Ala-TVITTEELQVTPTYAPVTTPLPTSAVPYDQRS | K141Q, S145T, P146Y, N149del, T152P, S161A | 32 |
| B21-762/794 | B21 (OPG211) | Biotin-β-Ala-GLQSPNPPLRNPLPQHDDYSPPQVHRPPP | N762del, P763del, P764del, P765del, Y766del, R767del, Q768P, R771Q, G772H, Y777S, S794P | 29 |
| B29/C23-196 | B29/C23 (OPG1) | Biotin-β-Ala-GSNISHKKVSYKDIIGSTIVDTK | E196K | 23 |
| E3-62/80 | E3 (OPG65) | Biotin-β-Ala-SSDDTPPRWSTTMDADTRPTDSDADAIIDD | I62T, F66S, M67T, T70M, E71D, K74T, P75R, D76P, A77T, A79S, M80D | 30 |
| E3-111/134 | E3 (OPG65) | Biotin-β-Ala-VIPVKKIIYWKGVNPVTVINEYCQITRRRDWS | A111V, D116Y, D119G, A120V, I125V, K134R | 32 |
| H3-45/64 | H3 (OPG108) | Biotin-β-Ala-VKDNEVMQEKRDVVIVNDDPDHYKDYVF | P45Q, N49D, K54N, A64V | 28 |
| MPXpep | A27 (OPG154) | Biotin-β-Ala-TEFFSTKAAKNPETKREAIVKAYGDDNEETLKQ | K27N, A30T, D39Y | 34 |
| Random * | - | Biotin-β-Ala- VTIKEYTATQRKLNFNEKDKESPEAADKTAEGF | - | 33 |
| Age (years) | OPXV IgG ELISA | MPXV Peptide IgG ELISA | ||
|---|---|---|---|---|
| No. (%) Positive | No. (%) Negative | No. (%) Positive | No. (%) Negative | |
| ≤23 | 39 (70%) | 17 (30%) | 4 (7%) | 52 (93%) |
| >23 | 24 (71%) | 10 (29%) | 7 (21%) | 27 (82%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taha, T.Y.; Townsend, M.B.; Pohl, J.; Karem, K.L.; Damon, I.K.; Mbala Kingebeni, P.; Muyembe Tamfum, J.-J.; Martin, J.W.; Pittman, P.R.; Huggins, J.W.; et al. Design and Optimization of a Monkeypox virus Specific Serological Assay. Pathogens 2023, 12, 396. https://doi.org/10.3390/pathogens12030396
Taha TY, Townsend MB, Pohl J, Karem KL, Damon IK, Mbala Kingebeni P, Muyembe Tamfum J-J, Martin JW, Pittman PR, Huggins JW, et al. Design and Optimization of a Monkeypox virus Specific Serological Assay. Pathogens. 2023; 12(3):396. https://doi.org/10.3390/pathogens12030396
Chicago/Turabian StyleTaha, Taha Y., Michael B. Townsend, Jan Pohl, Kevin L. Karem, Inger K. Damon, Placide Mbala Kingebeni, Jean-Jacques Muyembe Tamfum, James W. Martin, Phillip R. Pittman, John W. Huggins, and et al. 2023. "Design and Optimization of a Monkeypox virus Specific Serological Assay" Pathogens 12, no. 3: 396. https://doi.org/10.3390/pathogens12030396
APA StyleTaha, T. Y., Townsend, M. B., Pohl, J., Karem, K. L., Damon, I. K., Mbala Kingebeni, P., Muyembe Tamfum, J.-J., Martin, J. W., Pittman, P. R., Huggins, J. W., Satheshkumar, P. S., Bagarozzi, D. A., Jr., Reynolds, M. G., & Hughes, L. J. (2023). Design and Optimization of a Monkeypox virus Specific Serological Assay. Pathogens, 12(3), 396. https://doi.org/10.3390/pathogens12030396