

Composition of Culturable Microorganisms in Dusts Collected from Sport Facilities in Finland during the COVID-19 Pandemic

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Culturable Bacteria and Fungi in Samples of Sedimented Airborne Dusts Collected from Sport Facilities during the COVID-19 Pandemic
2.2. Numbers of Dust Particles and Fungi versus Bacterial Colony Counts in the Sedimented Airborne Dusts
2.3. Tracking Diversity of Fungal Colonies from the Samples of Settled Dusts Collected from Sport Facilities—Identification of the Isolates to Genus, Section and Species Levels
2.4. Numbers of Persons Visiting the Different Locations, Density of Particles and Microbial Colony Counts in the Settled Dusts
2.5. Summary of Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling Design
5.2. The Study Buildings
5.3. Collection of Sedimented Airborne Dust by a Standardised Method
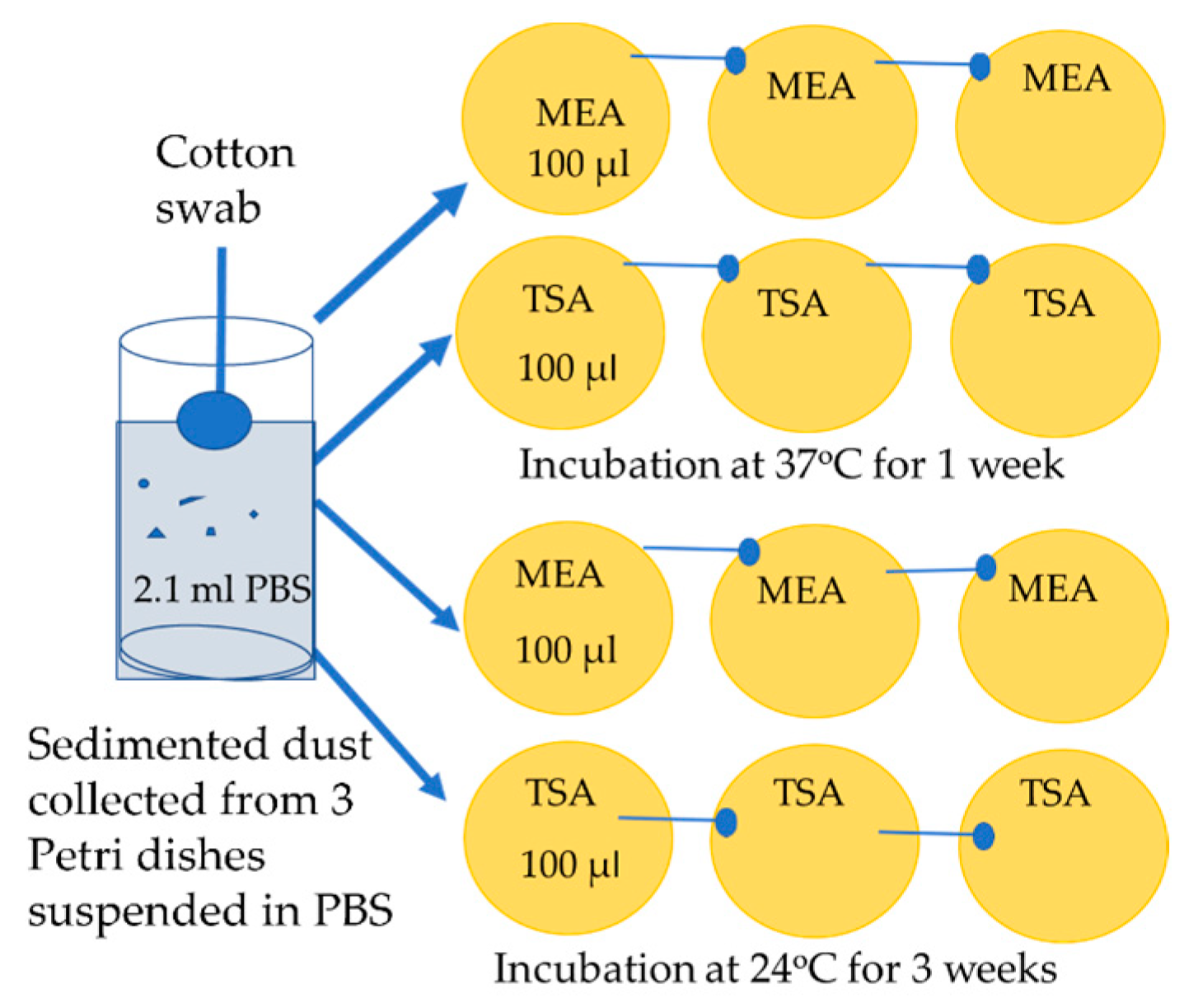
5.4. Cultivation of Dust Samples, Enumeration of Bacterial and Fungal Colonies, Isolation of Pure Cultures and Testing for Pathogenic Potential
5.5. Tracking Diversity of Fungal Isolates, Assignation to Genera and Sections, Testing for Pathogenic Potential, and Identification to the Species Level
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martini, M.; Gazzaniga, V.; Behzadifar, M.; Bragazzi, N.L.; Barberis, I. The history of tuberculosis: The social role of sanatoria for the treatment of tuberculosis in Italy between the end of the 19th century and the middle of the 20th. J. Prev. Med. Hyg. 2018, 59, E323–E327. [Google Scholar] [CrossRef] [PubMed]
- De Los Santos, E. Flashback: In the Fight Against Tuberculosis, Open Air Schools in Chicago Took an Unorthodox Approach: Keep Kids Outside, Even in Winter. Chicago Tribune. 1 May 2020. Available online: https://www.chicagotribune.com/history/ct-opinion-flashback-open-air-schools-tuberculosis-20200501-phyphmoe4vhhnelpjj2sck7fmi-story.html (accessed on 15 February 2023).
- Sipinen, J. Terveys on Kallein Aarteesi-Ihannekansalainen 1930-Luvun Suomalaisessa Terveysvalistuksessa. Suomi 2016, 3, 1–43. Available online: https://hybrislehti.net/terveys-on-kallein-aarteesi-ihannekansalainen-1930luvun-suomalaisessa-terveysvalistuksessa (accessed on 15 February 2023).
- Haahtela, T. Biodiversity for resilience—What is needed for allergic children. Pediatr. Allerg. Immunol. 2022, 33, e13779. [Google Scholar] [CrossRef] [PubMed]
- Pascal, M.; Corso, M.; Chanel, O.; Declercq, C.; Badaloni, C.; Cesaroni, G.; Henschel, S.; Meister, K.; Haluza, D.; Martin-Olmedo, P.; et al. Assessing the public health impacts of urban air pollution in 25 European cities: Results of the Aphekom project. Sci. Total Environ. 2013, 449, 390–400. [Google Scholar] [CrossRef] [PubMed]
- IARC. Press Release No. 221. Outdoor Air Pollution a Leading Environmental Cause of Cancer Deaths. 2013. Available online: https://www.iarc.who.int/wp-content/uploads/2018/07/pr221_E.pdf (accessed on 15 February 2023).
- LIPAS Database (LIPAS-Tietokanta). Computer Based Information System of Sports Sites and Facilities. University of Jyväskylä. 2020. Available online: https://www.lipas.fi/liikuntapaikat (accessed on 15 February 2023).
- WHO (World Health Organization). Physical Activity Strategy for the WHO European Region 2016–2025; WHO Regional Office for Europe: Copenhagen, Denmark, 2016; ISBN 978-928-905-147-7. [Google Scholar]
- Koivisto, T. Air Contaminants in Different Indoor Sports Facilities. Master’s Thesis, Aalto University, Aalto, Finland, 2021. Available online: https://aaltodoc.aalto.fi/handle/123456789/110685 (accessed on 15 February 2023).
- Salonen, H.; Salthammer, T.; Morawska, L. Human exposure to air contaminants in sports environments. Indoor Air 2020, 6, 1109–1129. [Google Scholar] [CrossRef]
- Ramos, C.A.; Viegas, C.; Cabo Verde, S.; Wolterbeek, T.H.; Almeida, S.M. Characterizing the fungal and bacterial microflora and concentrations in fitness centers. Indoor Built Environ. 2015, 25, 872–882. [Google Scholar] [CrossRef]
- Ramos, C.A.; Reis, J.F.; Almeida, T.; Wolterbeek, T.H.; Almeida, S.M. Estimating the inhaled dose of pollutants during indoor physical activity. Sci. Total Environ. 2015, 527–528, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Ramos, C.A.; Wolterbeek, T.H.; Almeida, S.M. Exposure to indoor air pollutants during physical activity in fitness centers. Build. Environ. 2014, 82, 349–360. [Google Scholar] [CrossRef]
- Boonrattanakij, N.; Yomchinda, S.; Lin, F.; Luzvisminda, M.; Bellotindos, L.; Lu, M.-C. Investigation and disinfection of bacteria and fungi in sports fitness centers. Environ. Sci. Pollut. Res. 2021, 28, 52576–52586. [Google Scholar] [CrossRef]
- Zheng, G.; Webster, T.; Salamova, A. Quaternary ammonium compounds: Bioaccumulation potentials in humans and levels in blood before and during the COVID-19 pandemic. Environ. Sci. Technol. 2021, 55, 14689–14698. [Google Scholar] [CrossRef]
- Dewey, H.; Jones, J.; Keating, M.; Budhathoki-Upret, J. Increased use of disinfectants during the COVID-19 pandemic and its potential impacts on health and safety. ACS Chem. Health Saf. 2022, 29, 27–38. [Google Scholar] [CrossRef]
- Koronavirus: Uusi Ohje Siivoukseen Suomalaisille Työpaikoille. Available online: https://www.uusisuomi.fi›Uutiset›Työterveys (accessed on 15 February 2023).
- Baddley, J.W.; Thompson, G.R.; Chen, S.C.A.; White, P.L.; Johnson, M.D.; Nguyen, M.H.; Schwartz, I.S.; Spec, A.; Ostrosky-Zeichner, L.; Jackson, B.R.; et al. Coronavirus Disease 2019-associated invasive fungal infection. Open Forum Infect. Dis. 2021, 8, ofab5. [Google Scholar] [CrossRef]
- Fortarezza, F.; Boscolo, A.; Pezzuto, F.; Lunardi, F.; Jesús Acosta, M.; Giraudo, C.; Del Vecchio, C.; Sella, N.; Tiberio, I.; Godi, I.; et al. Proven COVID-19-associated pulmonary aspergillosis in patients with severe respiratory failure. Mycoses 2021, 64, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, R.; Joshi, S.R.; Misra, A. Mucormycosis in COVID-19: Mucormycosis in COVID-19: A systematic review of cases reported worldwide and in India. Diabetes Metab. Syndr. 2021, 15, 102146. [Google Scholar] [CrossRef] [PubMed]
- Monika, P.; Chandraprabha, M.N. Risks of mucormycosis in the current Covid-19 pandemic: A clinical challenge in both immunocompromised and immunocompetent patients. Mol. Biol. Rep. 2022, 49, 4977–4988. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Varga, A.; Mikkola, R.; Vornanen-Winqvist, C.; Salo, J.; Kredics, L.; Kocsubé, S.; Salonen, H. Aspergillus was the dominant genus found during diversity tracking of cultivable potentially pathogenic indoor fungal isolates. Pathogens 2022, 11, 1171. [Google Scholar] [CrossRef] [PubMed]
- Viegas, C.; Alves, C.; Carolino, E.; Rosado, L.; Santos, C.S. Prevalence of fungi in indoor air with reference to gymnasiums with swimming pools. Indoor Built Environ. 2010, 19, 555–561. [Google Scholar] [CrossRef]
- Liang, Z.; Dong, C.B.; Liang, H.; Zhen, Y.X.; Zhou, R.L.; Han, Y.F.; Liang, Z.Q. A microbiome study reveals the potential relationship between the bacterial diversity of a gymnastics hall and human health. Sci. Rep. 2022, 12, 5663. [Google Scholar] [CrossRef]
- Ekanayake, A.; Rajapaksha, A.U.; Hewawasam, C.; Anand, U.; Bontempi, E.; Kurwadkar, S.; Biswas, J.K.; Vithanage, M. Environmental challenges of COVID-19 pandemic: Resilience and sustainability—A review. Environ. Res. 2022, 216, 114496. [Google Scholar] [CrossRef]
- Guo, J.; Liao, M.; He, B.; Liu, J.; Hu, X.; Yan, D.; Wang, J. Impact of the COVID-19 pandemic on household disinfectant consumption behaviors and related environmental concerns: A questionnaire-based survey in China. J. Environ. Chem. Eng. 2021, 9, 106168. [Google Scholar] [CrossRef]
- Lou, J.; Wang, W.; Lu, H.; Wang, L.; Zhu, L. Increased disinfection byproducts in the air resulting from intensified disinfection during the COVID-19 pandemic. J. Hazard. Mater. 2021, 418, 126249. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.I.; Tian, Y.; Taylor, J.W.; Bruns, T.D.; Hyvarinen, A.; Taubel, M. Passive dust collectors for assessing airborne microbial material. Microbiome 2015, 3, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, J.B.; Keady, P.B.; Brewer, T.E.; Clements, N.; Morgan, E.E.; Awerbuch, J.; Miller, S.L.; Fierer, N. Impacts of flood damage on airborne bacteria and fungi in homes after the 2013 Colorado front range flood. Environ. Sci. Technol. 2015, 49, 2675–2684. [Google Scholar] [CrossRef]
- Madsen, A.M.; Matthiesen, C.B.; Frederiksen, M.W.; Frederiksen, M.; Frankel, M.; Spilak, M.; Gunnarsen, L.; Timm, M. Sampling, extraction and measurement of bacteria, endotoxin, fungi and inflammatory potential of settling indoor dust. J. Environ. Monitor. 2012, 14, 3230–3239. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Biocidal resistance in clinically relevant microbial species: A major public health risk. Pathogens 2021, 10, 598. [Google Scholar] [CrossRef]
- Barabote, R.D.; Thekkiniath, J.; Strauss, R.E.; Vediyappan, G.; Fralick, J.A.; Francisco, M.J.S. Xenobiotic efflux in bacteria and fungi: A genomics update. Adv. Enzymol. Relat. Areas Mol. Biol. 2011, 77, 237–306. [Google Scholar]
- Andersen, B.; Frisvad, J.C.; Dunn, R.R.; Thrane, U. A pilot study on baseline fungi and moisture indicator fungi in Danish homes. J. Fungi 2021, 7, 71. [Google Scholar] [CrossRef]
- Stewart, S.L.; Grinshpun, S.A.; Willeke, K.; Terzieva, S.; Ulevicius, V.; Donnelly, J. Effect of impact stress on microbial recovery on an agar surface. Appl. Environ. Microbiol. 1995, 61, 1232–1239. [Google Scholar] [CrossRef] [Green Version]
- Cornet, M.; Gaillardin, C. pH signaling in human fungal pathogens: A new target for antifungal strategies. Eukaryot. Cell 2014, 13, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Davis, D. How human pathogenic fungi sense and adapt to pH: The link to virulence. Curr. Opin. Microbiol. 2009, 12, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+-permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Hautaniemi, M.; Salkinoja-Salonen, M.S. Toxic indole alkaloids avrainvillamide and stephacidin B produced by a biocide tolerant indoor mold Aspergillus westerdijkiae. Toxicon 2015, 99, 58–67. [Google Scholar] [CrossRef]
- Raksha; Singh, G.; Urhekar, A.D. Virulence factors detection in Aspergillus isolates from clinical and environmental samples. J. Clin. Diagn. Res. 2017, 11, DC13–DC18. [Google Scholar]
- Góralska, K.; Błaszkowska, J.; Dzikowiec, M. The occurrence of potentially pathogenic filamentous fungi in recreational surface water as a public health risk. J. Water Health 2020, 18, 127–144. [Google Scholar] [CrossRef]
- German Collection of Microorganisms and Cell Cultures GmbH: Catalogue (dsmz.de). Available online: https//dsmz.de (accessed on 15 February 2023).
- WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2022.
- Ruutu, P.; Valtonen, V.; Tiitanen, L.; Elonen, E.; Volin, L.; Veijalainen, P.; Ruutu, T. An outbreak of invasive aspergillosis in a haematologic unit. Scand J. Infect. Dis. 1987, 19, 347–351. [Google Scholar] [CrossRef]
- Ichai, P.; Saliba, F.; Baune, P.; Daoud, A.; Coilly, A.; Samuel, D. Impact of negative air pressure in ICU rooms on the risk of pulmonary aspergillosis in COVID-19 patients. Crit. Care 2020, 24, 538. [Google Scholar] [CrossRef]
- Dylag, M.; Spychała, K.; Zielinski, J.; Łagowski, D.; Gnat, S. Update on Stachybotrys chartarum—Black mold perceived as toxigenic and potentially pathogenic to humans. Biology 2022, 11, 352. [Google Scholar] [CrossRef]
- Selway, C.A.; Mills, J.G.; Weinstein, P.; Skelly, C.; Yadav, S.; Lowe, A.; Breed, M.F.; Weyrich, L.S. Transfer of environmental microbes to the skin and respiratory tract of humans after urban green space exposure. Environ. Int. 2020, 145, 106084. [Google Scholar] [CrossRef]
- Roslund, M.I.; Puhakka, R.; Grönroos, M.; Nurminen, N.; Oikarinen, S.; Gazali, A.M.; Cinek, O.; Kramná, L.; Siter, N.; Vari, H.K.; et al. Biodiversity intervention enhances immune regulation and health-associated commensal microbiota among daycare children. Sci. Adv. 2020, 6, eaba2578. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.; Bloomfield, S. Microbial exposures that establish immunoregulation are compatible with targeted hygiene. J. Allerg Clin. Immunol. 2021, 148, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Soininen, L.; Roslund, M.I.; Nurminen, N.; Puhakka, R.; Laitinen, O.H.; Hyöty, H.; Sinkkonen, A. Indoor green wall affects health-associated commensal skin microbiota and enhances immune regulation: A randomized trial among urban office workers. Sci Rep. 2022, 12, 6518. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. (Eds.) Introduction to Food and Air-Borne Fungi, 6th ed.; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002. [Google Scholar]
- Druzhinina, I.S.; Komon-Zelazowsa, M.; Kredics, L.; Hatvani, L.; Antal, Z.; Belayneh, T.; Kubicek, C.P. Alternative reproductive strategies of Hypocrea orientalis and genetically close but clonal Trichoderma longibrachiatum, both capable of causing invasive mycoses of humans. Microbiology 2008, 154, 3447–3459. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Hong, S.; Go, S.; Shin, H.; Frisvad, J.; Samson, R. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 2005, 97, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [Green Version]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Total Bacteria | Total Fungi | Microbial Colony Count CFU of Fungi, CFU of Bacteria (in Brackets) | |||
|---|---|---|---|---|---|---|
| Dusts | CFU | CFU | MEA 24 °C | TSA 24 °C | MEA 37 °C | TSA 37 °C |
| Dusts from sport facilities sedimented for 4 weeks during the COVID-19 pandemic (June–July 2021) | ||||||
| SF1 | 85 | 38 | 31 (ND) | 5 (25) | 2 (ND) | ND (60) |
| SF2a | 54 | <1 | ND (ND) | ND (33) | ND (ND) | ND (24) |
| SF2b | 13 | 2 | 2 (ND) | ND (2) | ND (ND) | ND (11) |
| SF3a | 9 | 6 | 2 (ND) | 2 (3) | 2 (3) | 1 (3) |
| SF3b | 14 | 4 | ND (ND) | 3 (14) | 1 (ND) | ND (ND) |
| SF4 | 5 | 15 | 8 (ND) | 4 (4) | 2 (ND) | 1 (1) |
| SF5 | 144 | 45 | 5 (1) | 30 (79) | 1 (ND) | 9 (65) |
| SF6 | 5 | 8 | 3 (ND) | ND (ND) | 3 (ND) | 2 (5) |
| SF7 | 23 | 4 | ND (ND) | ND (15) | 4 (ND) | ND (8) |
| Reference sites | ||||||
| Urban dusts sedimented for 2 weeks in public buildings before the pandemic (December 2018–April 2019) | ||||||
| SD1a | 192 | ND | ND (ND) | ND (116) | ND (ND) | ND (76) |
| SD1b | 223 | ND | 1(ND) | ND (120) | ND (ND) | ND (103) |
| SD2a | 94 | 1 | 1 (ND) | ND (57) | ND (3) | ND (34) |
| SD2b | 132 | 1 | 1 (ND) | ND (87) | ND (ND) | ND (45) |
| SD3a | 205 | 10 | 10 (ND) | ND (110) | ND (ND) | ND (95) |
| SD3b | 184 | 7 | 4(ND) | 2 (80) | 1 (ND) | ND (104) |
| SD3c | 42 | 2 | 1 (ND) | ND (8) | 1 (ND) | ND (34) |
| Rural dust from a hay barn with moldy hay collected for 4 weeks during the pandemic in November 2021 | ||||||
| CD1 | >1000 | 191 | 161 (ND) | ND (>500) | 30 (ND) | ND (>500) |
| Rural dust from a summer house collected for 4 weeks during the pandemic in November 2021 | ||||||
| CD2 | >100 | 26 | 3 (ND) | 23 (>100) | ND (2) | ND (>100) |
| Location Dust Sample | Numbers of PM in Samples | Total Microbial CFU in Sample | Fungal CFU in Sample | Proportion of Fungal CFU in Total Microbial CFU (%) |
|---|---|---|---|---|
| Dust samples from sport facilities sedimented for 4 weeks during the COVID-19 pandemic (June–July 2021) | ||||
| SF1 | >100 | 123 | 38 | 31 |
| SF2a | >30 | 54 | ND | ND |
| SF2b | <10 | 15 | 2 | 13 |
| SF3a | <10 | 15 | 6 | 40 |
| SF3b | <10 | 20 | 4 | 20 |
| SF4 | <10 | 20 | 15 | 85 |
| SF5 | >100 | 189 | 45 | 23 |
| SF6 | <10 | 14 | 8 | 62 |
| SF7 | <10 | 27 | 4 | 15 |
| Reference sites | ||||
| Urban dust samples sedimented for 2 weeks from public buildings before the pandemic (December 2018–April 2019) | ||||
| SD1a | NI | 192 | ND | <1 |
| SD1b | NI | 223 | ND | <1 |
| SD2a | NI | 95 | 1 | 1 |
| SD2b | NI | 133 | 1 | <1 |
| SD3a | NI | 215 | 10 | 4 |
| SD3b | NI | 191 | 7 | 4 |
| SD3c | NI | 44 | 2 | 5 |
| Rural dust samples from a hay barn with moldy hay sedimented for 4 weeks during the pandemic (November 2021) | ||||
| CD1 | >1000 | >2000 | 191 | <10 |
| Rural dust from a summer house sedimented for 4 weeks during pandemic (November 2021) | ||||
| CD2 | >1000 | >1000 | 26 | <1 |
| Location | Total CFU in Sample | |||||
|---|---|---|---|---|---|---|
| Dust Sample | Bacteria | Fungi | Genus/Section | CFU in Sample | Growth at 37 °C at pH 7.2 | Potentially Pathogenic CFU in Total Fungal CFU (%) |
| SF1 | 85 | 38 | Aspergillus section Nigri | 2 | + | |
| Aspergillus section Nidulantes | 3 | - | ||||
| Aspergillus section Flavi | 1 | + | 5 | |||
| Chaetomium cochliodes | 2 | - | ||||
| Unidentified colonies including Penicillium sp. | 30 | - | ||||
| SF2a | 54 | ND | ND | ND | ND | |
| SF2b | 13 | 6 | Aspergillus section Flavi | 2 | + | 100 |
| Yeasts | 4 | + | ||||
| SF3a | 9 | 6 | Aspergillus section Flavi | 2 | + | |
| Aspegillus section Fumigati | 1 | + | 100 | |||
| Aspergillus section Nigri | 2 | + | ||||
| Paecilomyces sp. | 1 | + | ||||
| SF3b | 14 | 4 | Aspergillus section Flavi | 1 | + | |
| Paecilomyces sp. | 1 | + | 50 | |||
| Unidentified colonies | 2 | - | ||||
| SF4 | 5 | 15 | Aspergillus section Fumigati | 4 | + | |
| Aspergillus section Flavi | 7 | + | 100 | |||
| Aspergillus section Nigri | 3 | + | ||||
| SF5 | 144 | 45 | Aspergillus section Flavi | 4 | + | |
| Aspergillus section Nigri | 6 | + | 22 | |||
| Unidentified colonies including Penicillium sp. | 37 | - | ||||
| SF6 | 5 | 9 | Aspergillus section Fumigati | 3 | + | |
| Aspergillus section Nigri | 2 | + | 88 | |||
| Paecilomyces sp. | 3 | + | ||||
| Penicillium sp. | 1 | - | ||||
| SF7 | 23 | 4 | Aspergillus section Flavi | 3 | + | |
| Paecilomyces sp. | 1 | + | 100 | |||
| Control dusts samples | ||||||
| CD1 | >500 | 191 | Aspergillus section Nigri | 21 | + | |
| Aspergillus section Fumigati | 9 | + | ||||
| Aspergillus section Circumdati | 3 | - | 18 | |||
| Trichoderma sp. | 1 | - | ||||
| Unidentified colonies including Penicillium sp. | 157 | - | ||||
| CD2 | >100 | 26 | Penicillium sp. | 16 | - | <1 |
| Aspergillus section Nidulantes | 10 | - | ||||
| Sport Facility | Dust Samples Sedimented for 4 Weeks | |||||
|---|---|---|---|---|---|---|
| Location | No. of Persons | Particles PM | Total Bacteria CFU | Total Fungi CFU | Pathogenic Fungi CFU | Pathogenic Fungi CFU in Total Fungal CFU (%) |
| Sport facilities | ||||||
| SF1 | ~5000 | >100 | 85 | 38 | 2 | 5 |
| SF2a | ~4000 | >30 | 54 | ND | ND | |
| SF2b | <50 | <10 | 13 | 6 | 6 | 100 |
| SF3a | ~1000 | <10 | 3 | 6 | 6 | 100 |
| SF3b | Closed | <10 | 14 | 4 | 2 | 50 |
| SF4 | ~300 | <10 | 5 | 15 | 15 | 100 |
| SF5 | ~5000 | >100 | 144 | 45 | 10 | 22 |
| SF6 | <50 | <10 | 5 | 8 | 8 | 88 |
| SF7 | <500 | <10 | 23 | 4 | 4 | 100 |
| Control sites | ||||||
| CD1 | <2 | >1000 | >500 | 191 | 34 | 18 |
| CD2 | <2 | >1000 | >500 | 26 | ND | <1 |
| Section | Species | Strain ID | Location of Isolation | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| rpb2 | cmdA | Reference | ||||
| Fumigati | Aspergillus fumigatus | AE1 | SF3a | OP295388 | [22] | |
| Aspergillus fumigatus | AE5 | SF5 | OP295389 | [22] | ||
| Aspergillus fumigatus | AH3 | SF6 | OP295390 | [22] | ||
| Nigri | Aspergillus niger | AE4 | SF4 | OP295391 | [22] | |
| Aspergillus tubingensis | ASN | SF5 | OP295393 | [22] | ||
| Flavi | Aspergillus flavus | AGE | SF3a | OP295387 | [22] | |
| Aspergillus flavus | AE2 | SF3b | OP295385 | [22] | ||
| Aspergillus flavus | AEH | SF4 | OP295386 | [22] | ||
| Aspergillus flavus | ASpf | SF2b | OP356696 | [22] | ||
| Paecilomyces sp. | Pec5 | SF7 | OP356695 | Recent study | ||
| Paecilomyces sp. | Pec3 | SF3b | OP356694 | Recent study | ||
| Chaetomium cochliodes | CH2 | SF1 | OP356691 | OP295395 | [22] | |
| Chaetomium cochliodes | CH3 | SF1 | OP356692 | [22] | ||
| Dust Sample Site | Location | Building | Occupancy (Number of Persons) in the Sampling Site during the Sampling Period | Sampling Site | Building Described in Ref [3] Code |
|---|---|---|---|---|---|
| SF1 | Vantaa | A bedrock shelter used as sport hall | ~500 | Athletics area | 11 |
| SF2a | Helsinki | Sport hall | ~4000 | Athletics area | 7 |
| SF2b | Helsinki | <50 | Gym | 7 | |
| SF3a | Espoo | Sport hall | ~900–1000 | Ball game area | 2 |
| SF3b | Espoo | (Closed during sampling) | Gym | 2 | |
| SF4 | Espoo | A bedrock shelter used as sport hall | ~300 | Table tennis area | 5 |
| SF5 | Helsinki | Sport hall | ~5000 | Gymnastics area | 6 |
| SF6 | Helsinki | Sport hall | <50 | Gym | 8 |
| SF7 | Vantaa | Sport hall | <500 | Gym | 13 |
| SD1a | Vantaa | School | 20–30 | Classroom | |
| SD1b | Vantaa | School | 20–30 | Classroom | |
| SD2a | Espoo | School | 20–30 | Classroom | |
| SD2b | Espoo | School | 20–30 | Classroom | |
| SD3a | Helsinki | Day care | 20–30 | Playroom | |
| SD3b | Helsinki | Day care | 20–30 | Playroom | |
| SD3c | Helsinki | Day care | 20–30 | Playroom | |
| CD1 | Hautjärvi | Hay barn (store of discharged mouldy hay) | <2 | Hay barn | |
| CD2 | Joroinen | Summer house (not cleaned since 1947) | <2 | Summer house |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersson, M.; Vornanen-Winqvist, C.; Koivisto, T.; Varga, A.; Mikkola, R.; Kredics, L.; Salonen, H. Composition of Culturable Microorganisms in Dusts Collected from Sport Facilities in Finland during the COVID-19 Pandemic. Pathogens 2023, 12, 339. https://doi.org/10.3390/pathogens12020339
Andersson M, Vornanen-Winqvist C, Koivisto T, Varga A, Mikkola R, Kredics L, Salonen H. Composition of Culturable Microorganisms in Dusts Collected from Sport Facilities in Finland during the COVID-19 Pandemic. Pathogens. 2023; 12(2):339. https://doi.org/10.3390/pathogens12020339
Chicago/Turabian StyleAndersson, Maria (Aino), Camilla Vornanen-Winqvist, Tuomas Koivisto, András Varga, Raimo Mikkola, László Kredics, and Heidi Salonen. 2023. "Composition of Culturable Microorganisms in Dusts Collected from Sport Facilities in Finland during the COVID-19 Pandemic" Pathogens 12, no. 2: 339. https://doi.org/10.3390/pathogens12020339