
Molecular Detection and Genetic Diversity of Tick-Borne Pathogens in Goats from the Southern Part of Thailand


1
Department of Protozoology, Faculty of Tropical Medicine, Mahidol University, Bangkok 10400, Thailand
2
Department of Pre-Clinic and Applied Animal Science, Faculty of Veterinary Science, Mahidol University, Salaya, Nakhon Pathom 73170, Thailand
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(4), 477; https://doi.org/10.3390/pathogens11040477
Submission received: 29 March 2022
/
Revised: 11 April 2022
/
Accepted: 14 April 2022
/
Published: 15 April 2022
(This article belongs to the Special Issue Molecular Epidemiology of Tick-Borne Protozoal and Rickettsial Diseases)
Abstract
:Tick-borne hemoprotozoan and rickettsial diseases affect the health and productivity of small ruminants in tropical and subtropical regions. Despite the large population of goats in the southern part of Thailand, there is limited information on the prevalence of tick-borne pathogens. In this study, polymerase chain reaction was used to detect the presence of Theileria spp., T. ovis, T. orientalis, Babesia ovis, Anaplasma ovis, and A. marginale in 262 goats from three provinces in the southern part of Thailand. In this investigation, Theileria spp. and A. ovis were detected while T. ovis, B. ovis, and A. marginale were not detected. Overall infection rates of Theileria spp. and A. ovis were 10.3% and 1.5%, respectively. The co-infections of two parasites was observed in 1.5% of goats. Sequence analysis showed the presence of T. luwenshuni and T. orientalis in the goat samples. This study is the first to use the molecular detection of T. orientalis in Thai goats, and presents genetic characterization using the major piroplasm surface protein (MPSP) gene. In the phylogenetic analysis, the T. orientalis MPSP sequence was classified as type 7. The A. ovis major surface protein 4 (MSP4) gene sequences shared high identities and similarity with each other and clustered with isolates from other regions. This study provides information about the prevalence and genetic diversity of tick-borne pathogens in goats in the study area, and is expected to be valuable for the development of effective control measures to prevent disease in animals in Thailand.
1. Introduction
Tick-borne infections are one of the most common causes of decreases in livestock productivity, particularly in tropical and subtropical regions [1]. Theileria spp., Babesia spp. and Anaplasma spp. are important causative agents of tick-borne diseases in small ruminants, and have considerable economic impact [2,3].
Small ruminants are vulnerable to several species of Theileria including Theileria lestoquardi, T. ovis, T. luwenshuni, T. uilenbergi, T. separata, T. recondita, Theileria sp. OT3 and Theileria sp. MK [4,5,6]. Among these species, T. lestoquardi, T. luwenshuni and T. uilenbergi have been described as pathogenic, while T. ovis, T. recondita and T. separata cause benign theileriosis, a subclinical infection in goats and sheep [4,7,8]. Babesia species known to infect small ruminants include Babesia ovis, B. motasi, and B. crassa [9]. B. ovis and B. motasi are known to cause ovine babesiosis [9,10]. B. ovis is considered to be highly pathogenic in sheep, with mortality rates ranging from 30 to 50% [10].
Anaplasma ovis and Anaplasma phagocytophilum are rickettsial gram-negative organisms that cause ovine anaplasmosis. [11,12]. A. ovis is a major causative agent of ovine anaplasmosis. However, infection with A. ovis in sheep and goats is usually asymptomatic [12]. The infection is transmitted biologically and mechanically by ticks, blood-sucking insects, and contaminated blood [13].
In Thailand, goats are farmed primarily in the southern parts of the country. The goats are usually reared outdoors in mixed smallholder farms, and are therefore exposed to a range of vector-borne illness transmitted by arthropods such as ticks and blood-sucking insects. The prevalence of tick-borne pathogens (TBPs) has been investigated in various parts of Thailand [14,15,16,17], although these studies have focused on cattle and buffalo, and there is little information on infections in small ruminants. In the present study we investigated TBPs in goats from the southern part of Thailand, using polymerase chain reaction (PCR) assays. We also examined the prevalence and genetic diversity of the pathogens in goats. The phylogenetic relationships among the isolates identified in this research and those isolated from different countries were assessed.
2. Results
2.1. PCR Detection of TBPs in Goat Samples
In the present study, none of the 262 blood samples obtained were positive for B. bovis, T. ovis, or A. marginale. Theileria spp. and A. ovis were detected in the goat populations (Table 1). It was found that 13.3% (35/262) of goats from the southern part of Thailand were positive for at least one pathogen. The infection rates of TBPs in Yala, Pattani, and Narathiwat provinces were 4.9% (13/262), 4.1% (11/262) and 2.3% (6/262). Theileria spp. (10.3%, 27/262) were the most prevalent single infection, followed by A. ovis (1.5%, 4/262). Co-infections with Theileria spp. and A. ovis were found in 1.5% (4/262) of samples.
2.2. Sequencing Analysis
The 15 Theileria spp. isolates (OM802536–OM802550) were sequenced and the sequences compared with each other and with previously published sequences. Fourteen of the isolates (OM802536–OM802549) had 99.3–100% similarity to T. luwenshuni isolates from goats in Thailand (MW307318, MW307319 and MW307320) and sheep in China (KC414093 and KC414097), and another (OM802550) revealed 99.8–100% similarity to T. orientalis isolates from sheep in China (KC414098, KC 414099 and KC414100), cattle in Pakistan (MG599097) and ticks in India (MT768053) (Figure 1). The T. orientalis MPSP gene sequences had 99.6–100% similarity to T. orientalis MPSP gene isolates from various countries (AB871321, AB871336, AB562533, LC125433, and KU356867). The nucleotide sequence identity value of A. ovis MSP4 in this study (OM830304–OM830306) was 100% among the isolated sequences and 98.8–99.1% with previously published sequences (MH908943, MT344082, LC229602 and MK290834) (Figure 2).
2.3. Phylogenetic Analysis
Phylogenetic analysis of the Theileria spp. 18s rRNA gene revealed that T. luwenshuni was present in the three provinces of the southern part of Thailand. In the phylogenetic tree, the 14 sequences from Yala, Pattani and Narathiwat formed a cluster with the T. luwenshuni gene sequences previously isolated from goats in Thailand and sheep in China (Figure 3). One isolate clustered with T. orientalis isolated from sheep in China and cattle from Australia and Pakistan (Figure 3). One T. orientalis MPSP gene sequence in this study was located in the type 7 clade, and was closely related to isolates reported in cattle from Thailand, Japan and Vietnam (Figure 4).
The phylogenetic tree showed that the A. ovis MSP4 genes from three isolates were confined to the same clade and had a close relationship with sequences from goats and sheep in China, Turkey and Kenya (Figure 5).
3. Discussion
Ovine tick-borne pathogen infections affect goats and sheep worldwide, and are widely distributed in tropical and subtropical areas [18]. In Southeast Asia, the occurrence of TBPs in goat and sheep has been investigated in countries including China [19,20,21], South Korea [22], Vietnam [23] and Myanmar [24]. However, little information on their prevalence and distribution in Thailand is available. In the present study, the prevalence and genetic characteristics of A. ovis and Theileria spp. infections of goats in three provinces from the southern part of Thailand were investigated.
Ovine anaplasmosis is a tick-borne disease caused by A. ovis, and affects goats and sheep worldwide [11]. The parasite is believed to induce asymptomatic infection in small ruminants [12], but cases of clinical illness have been reported, with symptoms including anaemia, pale mucous membranes, weight loss, abortion and death [25]. Little research has been done in Thailand into the prevalence of Anaplasma spp. infection in small ruminants. In this study, A. ovis was detected in 1.5% (4/262) of the goat samples. The prevalence of infection with the parasite was lower than previously reported in goats from Sa Kaeo province, Thailand (3.6%) [26], China (15.3–63.8%) [11,21,27], Bangladesh (14.7%) [28], Pakistan (16.6%) [29] and South Korea (17.9%) [22]. Several factors might have contributed to the differences in rates of infection with A. ovis, including vector activity, climate, variation in the susceptibilities of animals from different geographical areas and sample sizes. Further studies into vectors and seasonal dynamics, and the use of a larger number of samples from different provinces will be valuable for the more precise determination of the presence of A. ovis in Thailand.
The MSP4 gene of Anaplasma spp. is widely used for genetic characterization [30]. The sequences of the MSP4 genes obtained in the current study were highly conserved among the goat isolates, and were identical to previous isolates from China [31], Tur-key [32] and Portugal [33]. Phylogenetic analysis revealed that all of the A. ovis MSP4 sequences clustered into one clade, together with sequences from China, Kenya, Portugal, Cyprus, Tunisia and Turkey, suggesting that the A. ovis isolates in the study belong to one genotype. These findings are consistent with those of a previous study that reported that low or high genetic diversity of the MSP4 gene is associated with low or high prevalence of A. ovis [34,35]. The MSP4 gene in A. ovis evolves relatively rapidly, and has produced many pseudogenes [36]. Previous studies have identified two genotypes of A. ovis MSP4 from Uganda and four from China [27,37].
Theileriosis caused by Theileria spp is an important tick-borne disease of domestic animals and causes economic losses in many countries [38]. In this study, the overall prevalence of Theileria spp. was 10.3%, a proportion which is lower than the previously detected 41.3% in goats from Sakaeo province, Thailand [39]. In the neighbouring countries, the prevalence rates of Theileria spp. in goats was 34.7–52.7% from China [19,40,41], 33.3% from Myanmar [24], 8.5% from Bangladesh [42] and 13.4% from Uganda [37].
The 18S small subunit ribosomal RNA (18S rRNA) gene is widely used in the detection of Theileria spp., and this gene has been successfully used to distinguish the previously unknown Theileria spp. [19]. Sequence analysis revealed the presence of T. luwenshuni and T. orientalis in this study. The partial T. luwenshuni 18s rRNA gene sequence of the goat isolates from the southern part of Thailand had 99.3–100% nucleotide sequence identity with those of T. luwenshuni from Rayong province in Thailand and from strains isolated in China. A previous study reported a prevalence of 5% of T. luwenshuni in the Rayong and Chonburi provinces of Thailand [43]. T. luwenshuni is widely distributed in the small ruminant population in China [19,21,31,41], but its distribution in Thai goats is unknown. The results of the phylogenetic analysis revealed that the T. luwenshuni isolates in this research were closely related to T. luwenshuni isolates from China, suggesting that the parasite may have invaded Thailand through the passage of infected tick vectors or animals from China. Further epidemiological studies, taking into account mechanical or biological vectors for the transmission of T. luwenshuni and risk factor analysis, will be necessary to understand the current distribution of the pathogen in small ruminants in Thailand.
The partial T. orientalis 18s rRNA gene sequence of the goat isolate had 99.8–100% nucleotide sequence identity with those of the China, Pakistan and India isolates. T. orientalis has been reported to have a high prevalence in cattle and buffalo in Thailand [14,15,16]. This finding could indicate that the pathogen is in circulation among the animal farms in this country. To the best of the authors’ knowledge, this is the first report of the detection of T. orientalis in Thai goats. This agent has been detected in sheep blood samples from China [19] and Vietnam [14]. Haemaphysalis spp. are biological vectors for T. orientalis [44]. However, these tick species are not frequently found on livestock in Thailand, which are mainly infested with Rhipicephalus microplus [45]. The DNA of T. orientalis has been detected in the R. microplus [14]; however, it is unclear whether this tick may act as a mechanical or biological vector for this parasite. The biting arthropods are considered to be potential vectors for the transmission of Theileria spp. [46,47]. Further studies into tick vectors and biting arthropods will be needed to clarify the transmission of T. orientalis in Thailand.
The MPSP gene has been highly useful as a marker for investigating the phylogeny of T. orientalis, and is also used for diagnostic purposes [14,15,16,48]. T. orientalis isolates from different countries have been classified into 11 types (types 1–8 and types N1–N3) based on the MPSP gene sequences [48]. Phylogenetic analysis in this study revealed that the T. orientalis MPSP sequence of the goat isolate was of type 7 and formed a cluster with T. orientalis gene sequences previously isolated from cattle in Thailand, Vietnam and Japan. Previous research on genetic diversity of T. orientalis in cattle in Thailand categorized MPSP gene isolates into five types (types 1, 3, 5, 7 and N3) [14,15,16]. However, in the current study we were unable to detect other MPSP types due to the limited number of positive samples. Therefore, a large-scale study with a much larger sample size from different provinces should be performed.
In this study, B. ovis, T. ovis and A. marginale were not detected in goat samples, suggesting that these parasites are not common in the study area. However, T. ovis was detected in biting flies (Tabanus megalops) in Nakhon Pathom province, Thailand [47]. Further studies are required to determine whether these parasites are present in small ruminants in Thailand.
In conclusion, we detected the presence of A. ovis and Theileria spp. in domestic goats from the southern part of Thailand. The prevalence of the parasites was lower than those reported in several other countries. Sequence analysis showed that at least two species of Theileria currently exist in Thai goats. Here we describe for the first time the detection of T. orientalis in goat samples. The phylogenetic analysis revealed one T. orientalis MPSP type and A. ovis MSP4 type. Our results improve the understanding of the epidemiology of TBPs in the goat population in Thailand. However, continuous surveillance and updating of the occurrence of TBPs in small ruminants in this country should be undertaken.
4. Materials and Methods
4.1. Ethical Statement
The blood samples were obtained from a project aimed at the evaluation of Toxoplasma gondii and Neospora caninum specific recombinant proteins for the diagnosis of Toxoplasmosis and Neosporosis in cattle and goats. Use of goat blood samples in this study was approved by the Ethics and Animal Care and Use Committee of the Faculty of Veterinary Science, Mahidol University (Permit Number: MUVS-2021-12-53).
4.2. Animal Samples and DNA Extraction
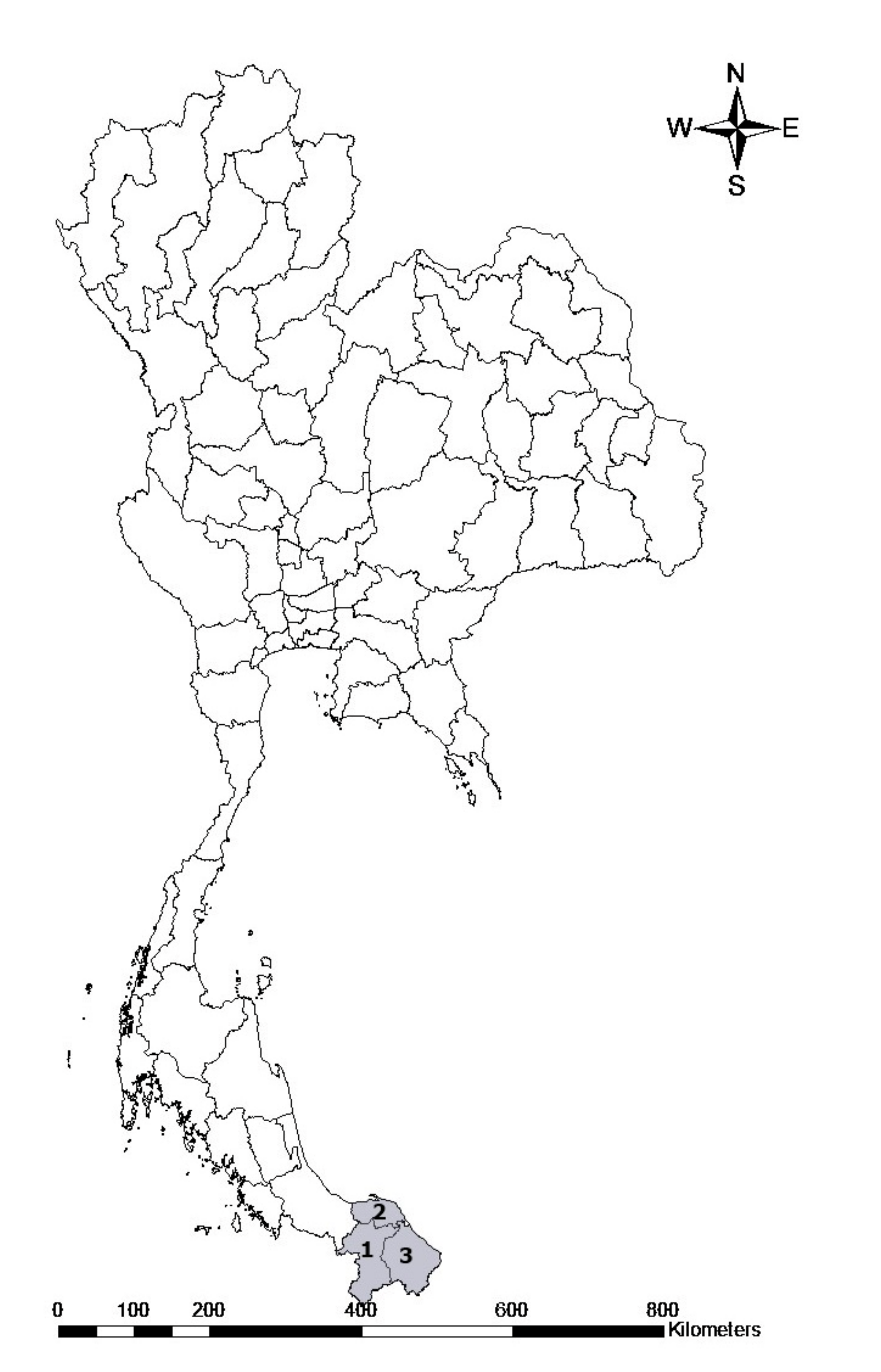
A total of 262 blood samples were collected from randomly selected herds from three provinces in the southern part of Thailand, including the Yala, Pattani and Narathiwat provinces (Figure 6). These blood samples were collected between March and April, 2021. The animals were restrained and blood was obtained from the jugular vein and transferred into 10 mL blood collection tubes containing anticoagulant. The samples were carried to the laboratory in a cooled box with ice packs and kept at 4 °C until they were used.
A total of 200 mL of blood sample was used to extract and purify genomic DNA using G-spin™ Total DNA extraction kits (iNtRON Biotechnology, Inc., Seongnam, South Korea) according to the manufacturer’s instructions. The extracted DNA was stored at −20 °C until they were used.
4.3. Molecular Detection of TBPs and DNA Sequencing
PCR assays were performed using primers for specific genes of Theileria spp., T. orientalis, T. ovis, B. ovis, A. ovis and A. marginale, as previously described [19,49,50,51,52]. Parasite detection was based on procedures previously described, as shown in Table 2.
A final volume of 25 µL was composed of 5 µL of 5X One Taq standard reaction buffer, 0.5 µL of deoxyribonucleotide triphosphates (dNTPs), 0.2 µM of each primer, and 0.125 µL of One Taq DNA polymerase (New England Biolabs, Ipswich, MA, USA), and topped up with distilled water to the final volume. DNA samples previously confirmed to be positive for Theileria spp., A. ovis and A. marginale were used as positive controls, whereas double-distilled water was used as the negative control. The positive controls of B. ovis and T. ovis were not available in our laboratory, so the correct size of PCR product was confirmed after sequencing. The PCR products were separated by gel electrophoresis on 1.5% agarose in 1 × TAE buffer and visualised using FluoroDye™ DNA Fluorescent Loading Dye (SMOBIO Technology, Hsinchu City, Taiwan) under a UV transilluminator.
Positive amplicons were snipped and extracted from agarose gel using Nucleo-Spin® Gel and PCR Clean-up (Macherey-Nagel, Düren, Germany). The extracts concentration was measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The purified product was submitted for Sanger DNA sequencing (Macrogen, Seoul, Korea). Bioedit version 7.2.6 (Tom Hall Ibis Biosciences, CA, USA) was used to analyze the nucleotide sequences, and GenBank BLASTn analysis was used to evaluate their identities and similarities [53]. The percent identities between nucleotides were computed by pairwise distances using MEGA version X software (www.megasoftware.net, accessed on 3 March 2022) [54].
4.4. Phylogenetic Analysis
The pathogen sequences in this study were compared to sequences from other regions of the world banked in genetic databases using MEGA version X software [53]. Multiple sequence alignment was conducted using the Muscle programme for each locus, and the genetic relatedness was determined using neighbour-joining or maximum likelihood. The confidence in the branching pattern of the trees was estimated using bootstrap tests with 1000 replications.
Author Contributions
Conceptualization, C.J.; methodology, R.U. and C.J.; formal analysis, C.J.; resources, R.U. and A.M.; writing—original draft preparation, R.U. and C.J.; writing—review and editing, R.U. and C.J. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Faculty of Veterinary Science, Mahidol University, Thailand.
Institutional Review Board Statement
The use of goat blood samples in this study was approved by the Ethics and Animal Care and Use Committee of the Faculty of Veterinary Science, Mahidol University (Permit Number: MUVS-2021-12-53, approval date 8 February 2022).
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to appreciate the contribution of the filed veterinarians and farmer in Yala, Pattani and Narathiwat Provincial Livestock Office for their support in the field sample collection. We would like to thank Morakot Kaewthamasorn, Faculty of Veterinary Science, Chulalongkorn University and Ketsarin Kamyingkird, Faculty of Veterinary Medicine, Kasetsart University for providing Theileria spp. and Anaplasma spp. control DNA samples.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Dantas-Torres, F.; Chomel, D.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Alessandra, T.; Santo, C. Tick-borne diseases in sheep and goats: Clinical and diagnostic aspects. Small Rumin. Res. 2012, 106, S11–S16. [Google Scholar] [CrossRef]
- Ghafar, A.; Abbas, T.; Rehman, A.; Sandhu, Z.U.; Cabezas-Cruz, A.; Jabbar, A. Systematic review of ticks and tick-borne pathogens of small ruminants in Pakistan. Pathogens 2020, 9, 937. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Luo, J.; Schnittger, L.; Lu, B.; Beyer, D.; Ma, M.; Guan, G.; Bai, Q.; Lu, C.; Ahmed, J. Phylogenetic analysis of Theileria species transmitted by Haemaphysalis qinghaiensis. Parasitol. Res. 2004, 92, 36–42. [Google Scholar] [CrossRef]
- Zaeemi, M.; Haddadzadeh, H.; Khazraiinia, P.; Kazemi, B.; Bandehpour, M. Identification of different Theileria species (Theileria lestoquardi, Theileria ovis, and Theileria annulata) in naturally infected sheep using nested PCR–RFLP. Parasitol. Res. 2010, 108, 837–843. [Google Scholar] [CrossRef]
- Bilgic, H.B.; Bakırcı, S.; Kose, O.; Unlu, A.H.; Hacılarlıoglu, S.; Eren, H.; Weir, W.; Karagenc, T. Prevalence of tick-borne haemoparasites in small ruminants in Turkey and diagnostic sensitivity of ingle-PCR and RLB. Parasites Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Altay, K.; Aktaş, M.; Dumanli, N. Theileria infections in small ruminants in the east and southeast Anatolia. Turk. Parazitol. Derg. 2007, 31, 268–271. [Google Scholar]
- Chae, J.S.; Allsopp, B.A.; Waghela, S.D.; Park, J.H.; Kakuda, T.; Sugimoto, C.; Allsopp, M.T.; Wagner, G.G.; Holman, P.J. A study of the systematics of Theileria spp. based upon small-subunit ribosomal RNA gene sequences. Parasitol. Res. 1999, 85, 877–883. [Google Scholar] [CrossRef]
- Schnittger, L.; Yin, H.; Gubbels, M.-J.; Beyer, D.; Niemann, S.; Jongejan, F.; Ahmed, J.S. Phylogeny of sheep and goat Theileria and Babesia parasites. Parasitol. Res. 2003, 91, 398–406. [Google Scholar]
- Ijaz, M.; Rehman, A.; Ali, M.M.; Umair, M.; Khalid, S.; Mehmood, K.; Hanif, A. Clinico-epidemiology and therapeutical trials on babesiosis in sheep and goats in Lahore, Pakistan. J. Anim. Plant Sci. 2013, 23, 666–669. [Google Scholar]
- Yang, J.; Li, Y.; Liu, Z.; Liu, J.; Niu, Q.; Ren, Q.; Chen, Z.; Guan, G.; Luo, J.; Yin, H. Molecular detection and characterization of Anaplasma spp. in sheep and cattle from Xinjiang, northwest China. Parasites Vectors 2015, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, A.; Rahbari, S.; Shayan, P.; Sadeghi-dehkordi, Z.; Bahonar, A. Molecular detection of Anaplasma marginale and Anaplasma ovis in sheep and goat in west highland pasture of Iran. Asian Pac. J. Trop. Biomed. 2017, 7, 455–459. [Google Scholar] [CrossRef]
- Mason, K.L.; González, M.V.; Chung, C.; Mousel, M.R.; White, S.N.; Taylor, J.B.; Scoles, G.A. Validation of an improved Anaplasma antibody competitive ELISA for detection of Anaplasma ovis antibody in domestic sheep. J. Vet. Diagn. Investig. 2017, 29, 763–766. [Google Scholar] [CrossRef] [Green Version]
- Altangerel, K.; Sivakumar, T.; Inpankaew, T.; Jittapalapong, S.; Terkawi, M.A.; Ueno, A.; Xuan, X.; Igarashi, I.; Yokoyama, N. Molecular prevalence of different genotypes of Theileria orientalis detected from cattle and water buffaloes in Thailand. J. Parasitol. 2011, 97, 1075–1079. [Google Scholar] [CrossRef]
- Jirapattharasate, C.; Moumouni, P.F.A.; Cao, S.; Iguchi, A.; Liu, M.M.; Wang, G.; Zhou, M.; Vudriko, P.; Changbunjong, T.; Sungpradit, S.; et al. Molecular epidemiology of bovine Babesia spp. and Theileria orientalis parasites in beef cattle from northern and northeastern Thailand. Parasitol. Int. 2016, 65, 62–69. [Google Scholar] [CrossRef]
- Jirapattharasate, C.; Moumouni, P.F.A.; Cao, S.; Iguchi, A.; Liu, M.M.; Wang, G.; Zhou, M.; Vudriko, P.; Efstratiou, A.; Changbunjong, T.; et al. Molecular detection and genetic diversity of bovine Babesia spp., Theileria orientalis, and Anaplasma marginale in beef cattle in Thailand. Parasitol. Res. 2017, 116, 751–762. [Google Scholar] [CrossRef]
- Saetiew, N.; Simking, P.; Inpankaew, T.; Wongpanit, K.; Kamyingkird, K.; Wongnakphet, S.; Stich, R.W.; Jittapalapong, S. Prevalence and genetic diversity of Anaplasma marginale infections in water buffaloes in Northeast Thailand. J. Trop. Med. Parasitol. 2015, 38, 9–16. [Google Scholar]
- Ros-García, A.; Barandika, J.F.; García-Pérez, A.L.; Juste, R.A.; Hurtado, A. Assessment of exposure to piroplasms in sheep grazing in communal mountain pastures by using a multiplex DNA bead-based suspension array. Parasites Vectors 2013, 6, 277. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Zhang, S.; Jia, L.; Xue, S.; Yu, L.; Kamyingkird, K.; Moumouni, P.F.A.; Moussa, A.A.; Zhou, M.; Zhang, Y.; et al. Molecular detection of Theileria species in sheep from northern China. J. Vet. Med. Sci. 2013, 75, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lv, Y.; Zhang, F.; Zhang, W.; Wang, J.; Cui, Y.; Wang, R.; Jian, F.; Zhang, L.; Ning, C. Molecular and phylogenetic analysis of Anaplasma spp. in sheep and goats from six provinces of China. J. Vet. Sci. 2016, 17, 523–529. [Google Scholar] [CrossRef]
- Li, Y.; Galon, E.M.; Guo, Q.; Rizk, M.A.; Moumouni, P.F.A.; Liu, M.; Li, J.; Ji, S.; Chahan, B.; Xuan, X. Molecular detection and identification of Babesia spp., Theileria spp., and Anaplasma spp. in sheep from border regions, Northwestern China. Front. Vet. Sci. 2020, 7, 630. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-G.; Kwon, O.-D.; Kwak, D. Molecular and Phylogenetic Analysis of Tick-Borne Pathogens in Ticks Parasitizing Native Korean Goats (Capra hircus coreanae) in South Korea. Pathogens 2020, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khukhuu, A.; Lan, D.T.; Long, P.T.; Ueno, A.; Li, Y.; Luo, Y.; Macedo, A.C.; Matsumoto, K.; Inokuma, H.; Kawazu, S.; et al. Molecular epidemiological survey of Theileria orientalis in Thua Thien Hue Province, Vietnam. J. Vet. Med. Sci. 2011, 73, 701–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bawm, S.; Kakisaka, K.; Thu, M.J.; Chel, H.M.; Oo, Y.M.N.; Soe, N.C.; Win, S.Y.; Htun, L.L.; Win, M.M.; Suzuki, H.; et al. First molecular detection of Theileria luwenshuni from goats in Myanmar. Parasitol. Res. 2018, 117, 3361–3364. [Google Scholar] [CrossRef]
- Yasini, S.P.; Khaki, Z.; Rahbari, S.; Kazemi, B.; Amoli, J.S.; Gharabaghi, A.; Jalali, S.M. Hematologic and clinical aspects of experimental ovine anaplasmosis caused by Anaplasma ovis in Iran. Iran. J. Parasitol. 2012, 7, 91–98. [Google Scholar]
- Kaewhom, P.; Srikijkasemwat, K. Molecular identification of Anaplasma ovis and Anaplasma marginale in sheep using PCR–RFLP method. Int. J. Agric. Technol. 2021, 17, 899–908. [Google Scholar]
- Liu, Z.; Ma, M.; Wang, Z.; Wang, J.; Peng, Y.; Li, Y.; Guan, G.; Luo, J.; Yin, H. Molecular survey and genetic identification of Anaplasma species in goats from Central and Southern China. Appl. Environ. Microbiol. 2012, 78, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Md Faruque, R.; Md Rahman, M.; Chowdhury, M.U.E. Epidemiology and molecular detection of Anaplasma spp. in goats from Chattogram district, Bangladesh. Vet. Med. Sci. 2022, 1–10. [Google Scholar] [CrossRef]
- Niaz, S.; Ur Rahman, Z.; Ali, I.; Cossío-Bayúgar, R.; Amaro-Estrada, I.; Alanazi, A.D.; Khattak, I.; Zeb, J.; Nasreen, N.; Khan, A. Molecular prevalence, characterization and associated risk factors of Anaplasma spp. and Theileria spp. in small ruminants in Northern Pakistan. Parasite 2021, 28, 3. [Google Scholar] [CrossRef]
- De la Fuente, J.; Lew, A.; Lutz, H.; Meli, M.L.; Hofmann-Lehmann, R.; Shkap, V.; Molad, T.; Mangold, A.J.; Almazán, C.; Naranjo, V.; et al. Genetic diversity of Anaplasma species major surface proteins and implications for anaplasmosis serodiagnosis and vaccine development. Anim. Health Res. Rev. 2005, 6, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ma, L.; Moumouni, P.F.A.; Jian, Y.; Wang, G.; Zhang, X.; Li, X.; Wang, G.; Lee, S.H.; Galon, E.M.; et al. Molecular survey and characterization of tick-borne pathogens in sheep from Qinghai, China. Small Rumin. Res. 2019, 175, 23–30. [Google Scholar] [CrossRef]
- Benedicto, B.; Ceylan, O.; Moumouni, P.F.A.; Lee, S.H.; Tumwebaze, M.A.; Li, J.; Galon, E.M.; Liu, M.; Li, Y.; Ji, S.; et al. Molecular detection and assessment of risk factors for tick-borne diseases in sheep and goats from Turkey. Acta Parasitol. 2020, 65, 723–732. [Google Scholar] [CrossRef]
- Pereira, A.; Parreira, R.; Cotão, A.J.; Nunes, M.; Vieira, M.L.; Azevedo, F.; Campino, L.; Maia, C. Tick-borne bacteria and protozoa detected in ticks collected from domestic animals and wildlife in central and southern Portugal. Ticks Tick Borne Dis. 2018, 9, 225–234. [Google Scholar] [CrossRef]
- De la Fuente, J.; Atkinson, M.W.; Naranjo, V.; Fernández de Mera, I.G.; Mangold, A.J.; Keating, K.A.; Kocan, K.M. Sequence analysis of the msp4 gene of Anaplasma ovis strains. Vet. Microbiol. 2007, 119, 375–381. [Google Scholar] [CrossRef]
- Han, R.; Yang, J.; Liu, Z.; Gao, S.; Niu, Q.; Hassan, M.A.; Luo, J.; Yin, H. Characterization of Anaplasma ovis strains using the major surface protein 1a repeat sequences. Parasit Vectors 2017, 10, 447. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Peasley, A.M.; Yang, J.; Li, Y.; Guan, G.; Luo, J.; Yin, H.; Brayton, K.A. The Anaplasma ovis genome reveals a high proportion of pseudogenes. BMC Genom. 2019, 20, 69. [Google Scholar] [CrossRef] [Green Version]
- Tumwebaze, M.A.; Byamukama, B.; Tayebwa, D.S.; Byaruhanga, J.; Angwe, M.K.; Galon, E.M.; Liu, M.; Lee, S.H.; Ringo, A.E.; Moumouni, P.F.A.; et al. First molecular detection of Babesia ovis, Theileria spp., Anaplasma spp., and Ehrlichia ruminantium in goats from Western Uganda. Pathogens 2020, 9, 895. [Google Scholar] [CrossRef]
- Ben, J.; Mans, R.P.; Latif, A.A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. 2015, 4, 104–118. [Google Scholar]
- Kaewhom, P.; Thitasarn, W. The prevalence of Theileria spp. of goat in Watthana Nakhon district, Sakaeo province. J. Mahanakorn Vet. Med. 2017, 12, 57–66. [Google Scholar]
- Yu, Z.Q.; Song, J.K.; Zhang, H.J.; Liu, T.L.; Fan, X.C.; Zhao, G.H. Molecular characterization of Theileria spp. In goats from Shaanxi province, Northwestern China. J. Parasitol. 2018, 104, 726–731. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.H.; Upadhyay, A.; Zhao, J.G.; Huang, L.Y.; Liao, C.H.; Han, Q. Identification of Theileria spp. and investigation of hematological profiles of their infections in goats in Hainan Island, China. Parasite 2022, 29, 13. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Rudra, P.G.; Singha, S.; Das, T.; Gebrekidan, H.; Uddin, B.; Chowdhury, M.Y.E. Molecular Epidemiology and Characterization of Theileria in Goats. Protist 2021, 2, 125804. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.L.C.; Nugraheni, Y.R.; Tiawsirisup, S.; Saiwichai, T.; Thiptara, A.; Kaewthamasorn, M. Development of a novel multiplex PCR assay for the detection and differentiation of Plasmodium caprae from Theileria luwenshuni and Babesia spp. in goats. Acta Trop. 2021, 220, 105957. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K. A review of the taxonomy of Theileria sergenti/buffeli/orientalis group parasite in calltle. J. Protozool. Res. 1992, 2, 87–96. [Google Scholar]
- Ahantarig, A.; Trinachartvanit, W.; Milne, J.R. Tick-borne pathogens and diseases of animals and humans in Thailand. Southeast Asian J. Trop. Med. Public Health 2008, 39, 1015–1032. [Google Scholar]
- Hammer, J.F.; Jenkins, C.; Bogema, D.; Emery, D. Mechanical transfer of Theileria orientalis: Possible roles of biting arthropods, colostrum and husbandry practices in disease transmission. Parasite Vector. 2016, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Jirapattharasate, C.; Changbunjong, T.; Sedwisai, P.; Weluwanarak, T. Molecular detection of piroplasms in haematophagus flies in the Nakhon Pathom and Kanchanaburi Provinces, Thailand. Vet. Integr. Sci. 2018, 16, 123–133. [Google Scholar]
- Sivakumar, T.; Hayashida, K.; Sugimoto, C.; Yokoyama, N. Evolution and genetic diversity of Theileria. Infect. Genet. Evol. 2014, 27, 250–263. [Google Scholar] [CrossRef] [Green Version]
- Torina, A.; Agnone, A.; Blanda, V.; Alongi, A.; D’Agostino, R.; Caracappa, S.; Marino, A.M.; Di Marco, V.; De La Fuente, J. Development and validation of two PCR tests for the detection of and differentiation between Anaplasma ovis and Anaplasma marginale. Ticks Tick Borne Dis. 2012, 3, 283–287. [Google Scholar] [CrossRef]
- Aktaş, M.; Altay, K.; Dumanlı, N. Development of a polymerase chain reaction method for diagnosis of Babesia ovis infection in sheep and goats. Vet. Parasitol. 2005, 133, 277–281. [Google Scholar] [CrossRef]
- Altay, K.; Dumanli, N.; Holman, P.J.; Aktaş, M. Detection of Theileria ovis in naturally infected sheep by nested PCR. Vet. Parasitol. 2005, 127, 99–104. [Google Scholar] [CrossRef]
- Ota, N.; Mizuno, D.; Kuboki, N.; Igarashi, I.; Nakamura, Y.; Yamashina, H.; Hanzaike, T.; Fujii, K.; Onoe, S.; Hata, H.; et al. Epidemiological survey of Theileria orientalis infection in grazing cattle in the eastern part of Hokkaido, Japan. J. Vet. Med. Sci. 2009, 71, 937–944. [Google Scholar] [CrossRef] [Green Version]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
Figure 1.
Nucleotide sequence alignment results of T. orientalis 18S rRNA gene isolate from goat in this study (OM802550) and database sequences.
Figure 1.
Nucleotide sequence alignment results of T. orientalis 18S rRNA gene isolate from goat in this study (OM802550) and database sequences.

Figure 2.
Nucleotide sequence alignment results of MSP4 gene of A. ovis isolates from goats (OM830304–OM830306) in this study and database sequences.
Figure 2.
Nucleotide sequence alignment results of MSP4 gene of A. ovis isolates from goats (OM830304–OM830306) in this study and database sequences.
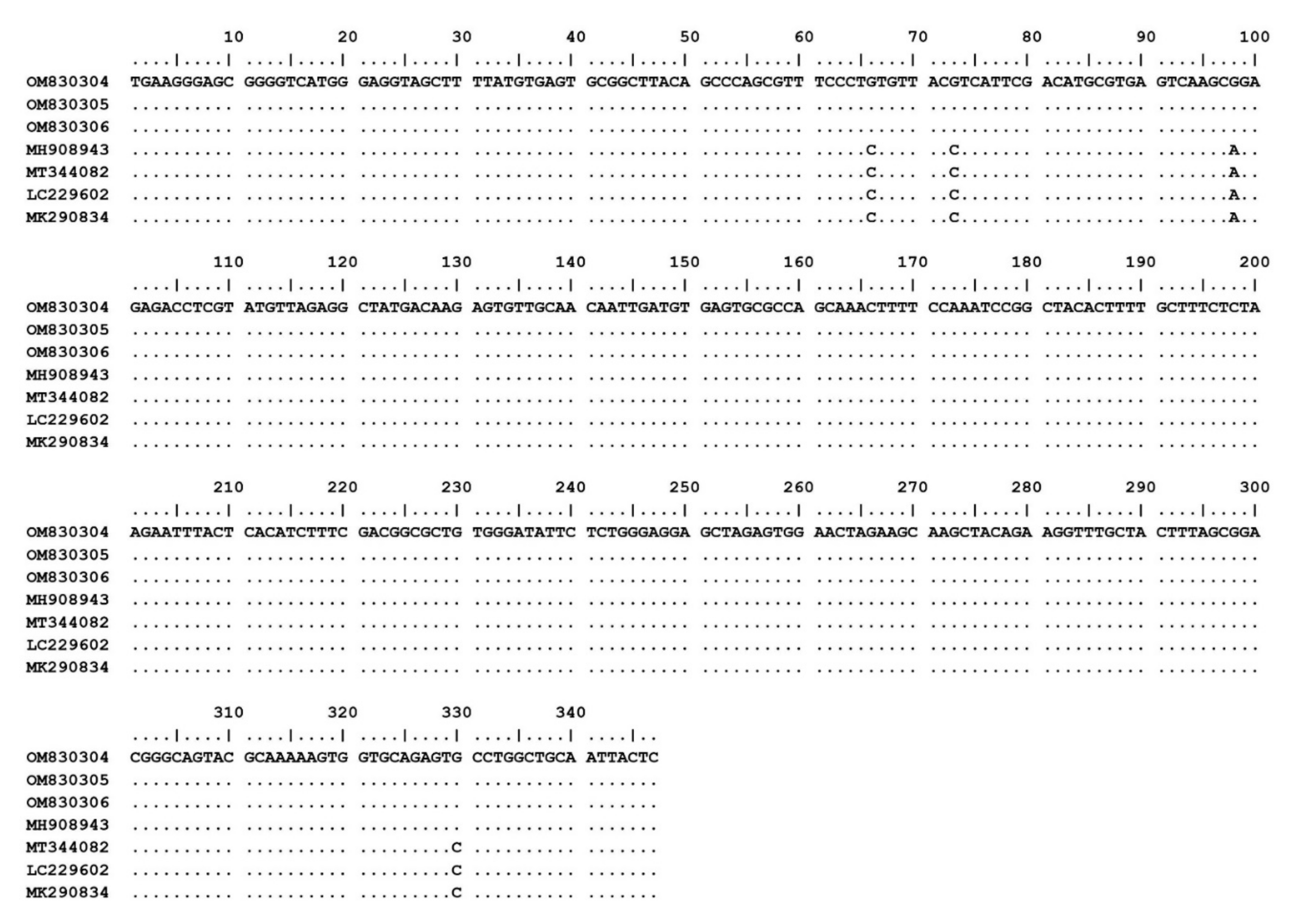
Figure 3.
The phylogenetic tree of 18S rRNA gene partial sequences of T. luwenshuni and T. orientalis in this study (boldface letters) and representative Theileria species. The tree was constructed using maximum likelihood with the Kimura-2 method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. B. crassa and B. motasi 18S rRNA gene partial sequences were used as outgroups.
Figure 3.
The phylogenetic tree of 18S rRNA gene partial sequences of T. luwenshuni and T. orientalis in this study (boldface letters) and representative Theileria species. The tree was constructed using maximum likelihood with the Kimura-2 method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. B. crassa and B. motasi 18S rRNA gene partial sequences were used as outgroups.
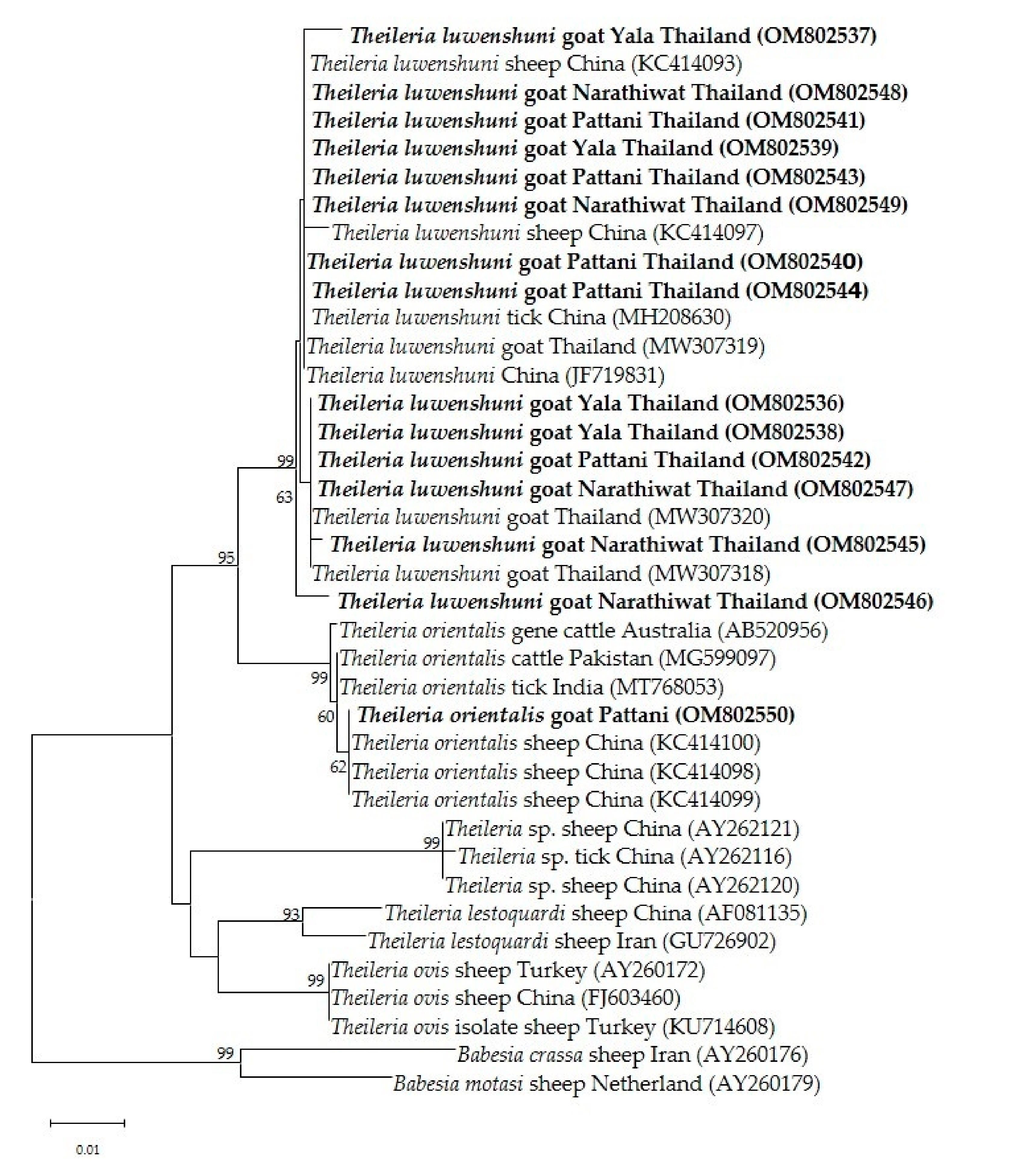
Figure 4.
The phylogenetic tree of T. orientalis MPSP gene sequence in this study (boldface letters) and from global isolates. The tree was constructed using the neighbor-joining method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. The Tams1 gene of T. annulate and T. parva were used as outgroups.
Figure 4.
The phylogenetic tree of T. orientalis MPSP gene sequence in this study (boldface letters) and from global isolates. The tree was constructed using the neighbor-joining method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. The Tams1 gene of T. annulate and T. parva were used as outgroups.
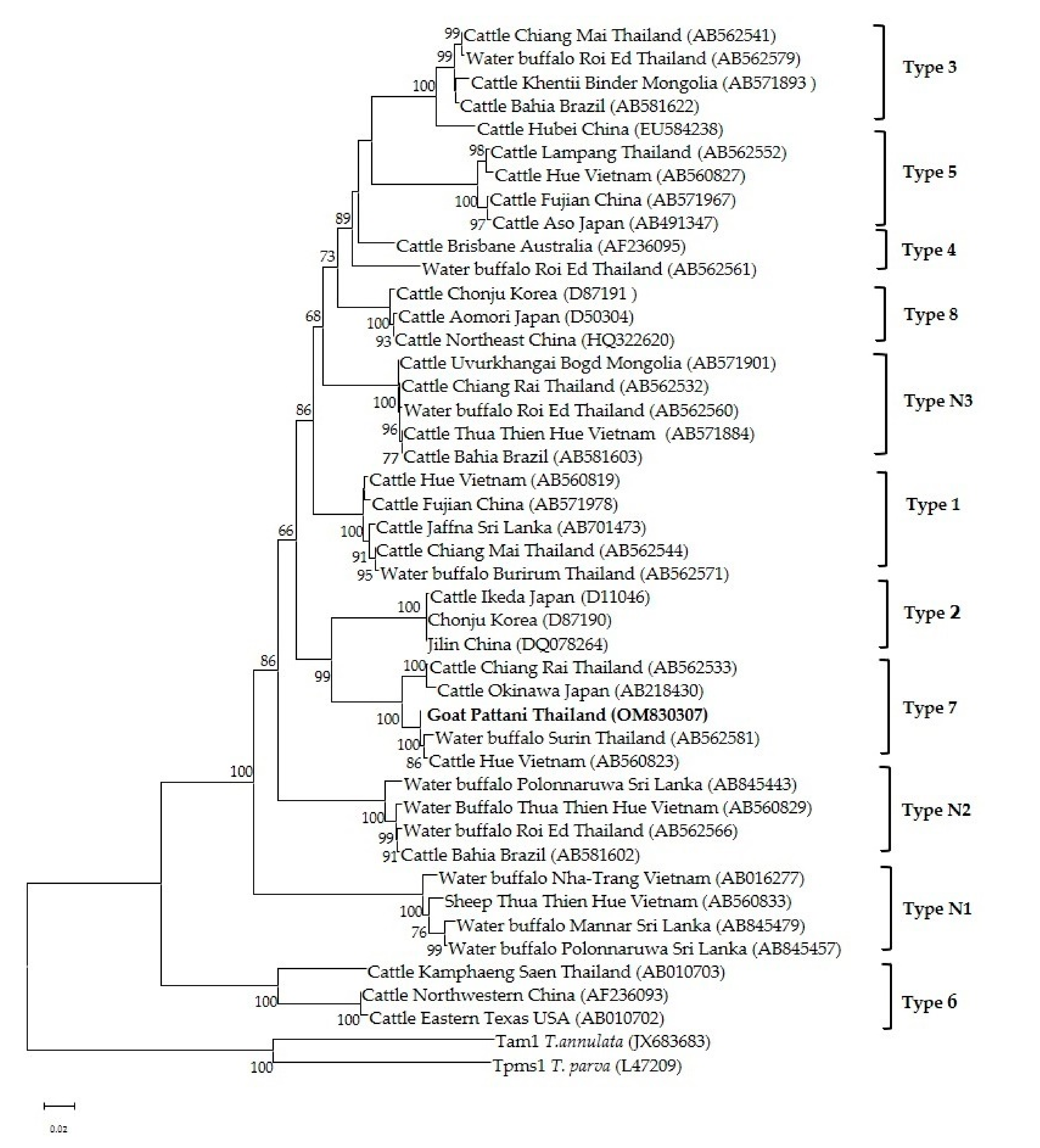
Figure 5.
Phylogenetic relationship of A. ovis MSP4 sequences from goats in this study (boldface letters) and reference sequences from Genbank database. The tree was constructed using maximum likelihood with Kimura-2 method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. The sequences of A. marginale and A. centrale MSP4 gene were used as outgroups.
Figure 5.
Phylogenetic relationship of A. ovis MSP4 sequences from goats in this study (boldface letters) and reference sequences from Genbank database. The tree was constructed using maximum likelihood with Kimura-2 method with 1000 bootstrap replications. Bootstrap values are indicated at each node and the numbers over 60 are shown in the tree. The sequences of A. marginale and A. centrale MSP4 gene were used as outgroups.

Figure 6.
Map of sampling areas in the Sothern part of Thailand. The numbers represent the provinces where the goat samples were collected: 1. Yala, 2. Pattani, and 3. Narathiwat.
Figure 6.
Map of sampling areas in the Sothern part of Thailand. The numbers represent the provinces where the goat samples were collected: 1. Yala, 2. Pattani, and 3. Narathiwat.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The PCR screening results for Theileria spp. ana A. ovis infections in goats the from the southern region.
Table 1.
The PCR screening results for Theileria spp. ana A. ovis infections in goats the from the southern region.
| Province | No. of Tested Goats | A. ovis (%) | Theileria spp. (%) | Mixed Infections (%) | Isolated Number of T. luwenshuni | Isolated Number of T. orientalis |
|---|---|---|---|---|---|---|
| Yala | 100 | 3 (1.1) | 11 (4.2) | 2 (0.7) | 4 | 0 |
| Pattani | 100 | 1(0.4) | 10 (3.8) | 1 * (0.4) | 5 | 1 |
| Narathiwat | 62 | 0 | 6 (2.3) | 1 * (0.4) | 5 | 0 |
| Total | 262 | 4 (1.5) | 27 (10.3) | 4 (1.5) | 14 | 1 |
* Co-infections between T. luwenshuni and A. ovis.
Table 2.
List of target genes and primers used for PCR assays.
| Target Gene | Assay | Primer Sequences (5′→3′) | Fragment (bp) | Annealing Temp (°C) | Reference | |
|---|---|---|---|---|---|---|
| Forward | Reverse | |||||
| A. ovis (MSP4) | PCR | TGAAGGGAGCGGGGTCATGGG | GAGTAATTGCAGCCAGGCACTCT | 347 | 62 | [49] |
| A. marginale (MSP4) | PCR | CTGAAGGGGGAGTAATGGG | GGTAATAGCTGCCAGAGATTCC | 344 | 60 | [49] |
| B. ovis (18S rRNA) | PCR | TGGGCAGGACCTTGGTTCTTCT | CCGCGTAGCGCCGGCTAAATA | 549 | 62 | [50] |
| T. ovis (18S rRNA) | PCR | TCGAGACCTTCGGGT | TCCGGACATTGTAAAACAAA | 520 | 60 | [51] |
| T. orientalis (MPSP) | PCR | CTTTGCCTAGGATACTTCCT | ACGGCAAGTGGTGAGAACT | 776 | 58 | [52] |
| Theileria spp. (18S rRNA) | PCR nPCR | GAAACGGCTACCACATCT TTAAACCTCTTCCAGAGT | AGTTTCCCCGTGTTGAGT TCAGCCTTGCGACCATAC | 778 581 | 55 55 | [19] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Udonsom, R.; Mahittikorn, A.; Jirapattharasate, C. Molecular Detection and Genetic Diversity of Tick-Borne Pathogens in Goats from the Southern Part of Thailand. Pathogens 2022, 11, 477. https://doi.org/10.3390/pathogens11040477
AMA Style
Udonsom R, Mahittikorn A, Jirapattharasate C. Molecular Detection and Genetic Diversity of Tick-Borne Pathogens in Goats from the Southern Part of Thailand. Pathogens. 2022; 11(4):477. https://doi.org/10.3390/pathogens11040477
Chicago/Turabian StyleUdonsom, Ruenruetai, Aongart Mahittikorn, and Charoonluk Jirapattharasate. 2022. "Molecular Detection and Genetic Diversity of Tick-Borne Pathogens in Goats from the Southern Part of Thailand" Pathogens 11, no. 4: 477. https://doi.org/10.3390/pathogens11040477
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.