
Survival and Detection of Bivalve Transmissible Neoplasia from the Soft-Shell Clam Mya arenaria (MarBTN) in Seawater

 , ,
, ,  and
and
Abstract
:
{kind=link}
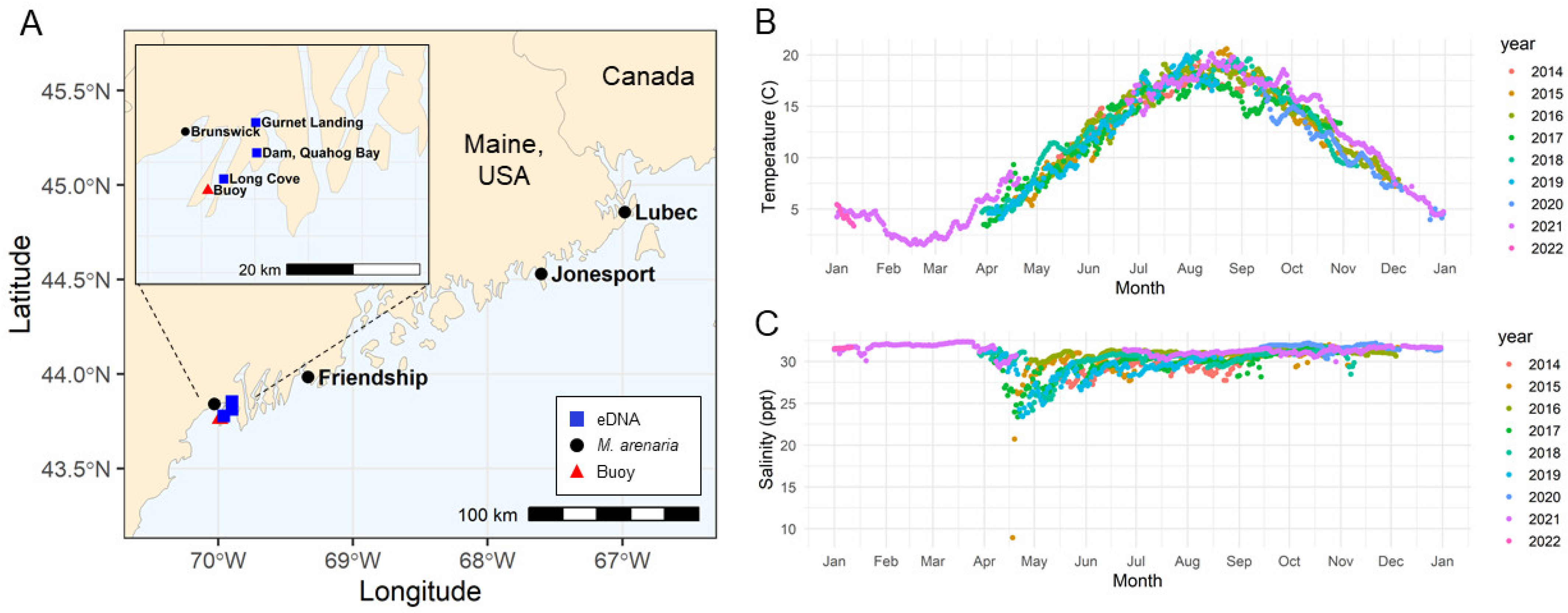{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Survival of MarBTN Cells in Seawater and the Effect of Environmental Variables
2.2. Detection of MarBTN eDNA Released from Diseased Clams in Laboratory Aquaria
2.3. Detection of MarBTN in Natural Sites of Known Endemic BTN
3. Discussion
4. Materials and Methods
4.1. Temperature and Salinity Data Access
4.2. Collection of Clams and MarBTN Cells
4.3. Counting Live Cells in Artificial Seawater
4.4. Validation of Erythrosine B Live/Dead Staining
4.5. Short-Term Cell Survival Assays
4.6. Long-Term Cell Survival Assays
4.7. eDNA Extraction from Aquaria
4.8. Field Seawater Collection and Extraction of MarBTN eDNA
4.9. qPCR of Hemocyte DNA and eDNA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murgia, C.; Pritchard, J.K.; Kim, S.Y.; Fassati, A.; Weiss, R.A. Clonal origin and evolution of a transmissible cancer. Cell 2006, 126, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Rebbeck, C.A.; Thomas, R.; Breen, M.; Leroi, A.M.; Burt, A. Origins and evolution of a transmissible cancer. Evol. Int. J. Org. Evol. 2009, 63, 2340–2349. [Google Scholar] [CrossRef]
- Pearse, A.M.; Swift, K. Allograft theory: Transmission of devil facial-tumour disease. Nature 2006, 439, 549. [Google Scholar] [CrossRef] [PubMed]
- Murchison, E.P.; Schulz-Trieglaff, O.B.; Ning, Z.; Alexandrov, L.B.; Bauer, M.J.; Fu, B.; Hims, M.; Ding, Z.; Ivakhno, S.; Stewart, C.; et al. Genome sequencing and analysis of the Tasmanian devil and its transmissible cancer. Cell 2012, 148, 780–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzger, M.J.; Reinisch, C.; Sherry, J.; Goff, S.P. Horizontal transmission of clonal cancer cells causes leukemia in soft-shell clams. Cell 2015, 161, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, B.J. Neoplastic diseases of commercially important marine bivalves. Aquat. Living Resour. 2004, 17, 449–466. [Google Scholar] [CrossRef]
- Carballal, M.J.; Barber, B.J.; Iglesias, D.; Villalba, A. Neoplastic diseases of marine bivalves. J. Invertebr. Pathol. 2015, 131, 83–106. [Google Scholar] [CrossRef]
- AboElkhair, M.; Synard, S.; Siah, A.; Pariseau, J.; Davidson, J.; Johnson, G.; Greenwood, S.; Casey, J.; Berthe, F.; Cepica, A. Reverse transcriptase activity in tissues of the soft shell clam Mya arenaria affected with haemic neoplasia. J. Invertebr. Pathol. 2009, 102, 133–140. [Google Scholar] [CrossRef]
- Oprandy, J.J.; Chang, P.W.; Pronovost, A.D.; Cooper, K.R.; Brown, R.S.; Yates, V.J. Isolation of a viral agent causing hematopoietic neoplasia in the soft-shell clam, Mya arenaria. J. Invertebr. Pathol. 1981, 38, 45–51. [Google Scholar] [CrossRef]
- Sunila, I.; Farley, C. Environmental limits for survival of sarcoma cells from the soft-shell clam Mya arenaria. Dis. Aquat. Org. 1989, 7, 111–115. [Google Scholar] [CrossRef]
- Moore, J.D. Pathogenesis of disseminated neoplasia in eastern Pacific Mytilus trossulus. Ph.D. Dissertation, University of Washington, Washington, DC, USA, 1993. [Google Scholar]
- Metzger, M.J.; Villalba, A.; Carballal, M.J.; Iglesias, D.; Sherry, J.; Reinisch, C.; Muttray, A.F.; Baldwin, S.A.; Goff, S.P. Widespread trans-mission of independent cancer lineages within multiple bivalve species. Nature 2016, 534, 705–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzger, M.J.; Goff, S.P. A Sixth Modality of Infectious Disease: Contagious Cancer from Devils to Clams and Beyond. PLoS Pathog. 2016, 12, e1005904. [Google Scholar] [CrossRef] [Green Version]
- A Yonemitsu, M.; Giersch, R.M.; Polo-Prieto, M.; Hammel, M.; Simon, A.; Cremonte, F.; Avilés, F.T.; Merino-Véliz, N.; Burioli, E.A.; Muttray, A.F.; et al. A single clonal lineage of transmissible cancer identified in two marine mussel species in South America and Europe. eLife 2019, 8, e47788. [Google Scholar] [CrossRef]
- Hammel, M.; Simon, A.; Arbiol, C.; Villalba, A.; Burioli, E.A.V.; Pepin, J.F.; Lamy, J.B.; Benabdelmouna, A.; Bernard, I.; Houssin, M.; et al. Prevalence and polymorphism of a mussel transmissible cancer in Europe. Mol. Ecol. 2021, 31, 736–751. [Google Scholar] [CrossRef]
- Skazina, M.; Odintsova, N.; Maiorova, M.; Ivanova, A.; Vainola, R.; Strelkov, P. First description of a widespread Mytilus trossu-lus-derived bivalve transmissible cancer lineage in M. trossulus itself. Sci. Rep. 2021, 11, 5809. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Souto, D.; Bruzos, A.L.; Diaz, S.; Rocha, S.; Pequeño-Valtierra, A.; Roman-Lewis, C.F.; Alonso, J.; Rodriguez, R.; Costas, D.; Rodriguez-Castro, J.; et al. Mitochondrial genome sequencing of marine leukaemias reveals cancer contagion between clam species in the Seas of Southern Europe. eLife 2022, 11, e66946. [Google Scholar] [CrossRef]
- Dujon, A.M.; Bramwell, G.; Roche, B.; Thomas, F.; Ujvari, B. Transmissible cancers in mammals and bivalves: How many examples are there? Predictions indicate widespread occurrence. Bioessays 2021, 43, e2000222. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.S.; Wolke, R.E.; Saila, S.B.; Brown, C.W. Prevalence of neoplasia in 10 New England populations of the soft-shell clam (Mya arenaria). Ann. N. Y. Acad. Sci. 1977, 298, 522–534. [Google Scholar] [CrossRef]
- Yevich, P.P.; Barszcz, C.A. Neoplasia in soft-shell clams (Mya arenaria) collected from oil-impacted sites. Ann. N. Y. Acad. Sci. 1978, 298, 409–426. [Google Scholar] [CrossRef]
- Farley, C.A.; Plutschak, D.L.; Scott, R.F. Epizootiology and distribution of transmissible sarcoma in Maryland softshell clams, Mya arenaria, 1984–1988. Environ. Health Perspect. 1991, 90, 35–41. [Google Scholar] [PubMed] [Green Version]
- Sunila, I. Serum-cell interactions in transmission of sarcoma in the soft shell clam, Mya arenaria L. Comp. Biochem. Physiol. Part A: Physiol. 1992, 102, 727–730. [Google Scholar] [CrossRef]
- Sticker, A. Transplantables Rundzellensarkom des Hundes. J. Cancer Res. Clin. Oncol. 1906, 4, 227–314. [Google Scholar] [CrossRef]
- Wade, H. An experimental investigation of infective sarcoma of the dog, with a consideration of its relationship to cancer. J. Pathol. Bacteriol. 1908, 12, 384–425. [Google Scholar] [CrossRef]
- Hamede, R.K.; McCallum, H.; Jones, M. Biting injuries and transmission of Tasmanian devil facial tumour disease. J. Anim. Ecol. 2013, 82, 182–190. [Google Scholar] [CrossRef]
- Bayer, S.R.; Countway, P.D.; Wahle, R.A. Developing an eDNA toolkit to quantify broadcast spawning events of the sea scallop Placopecten magellanicus: Moving beyond fertilization assays. Mar. Ecol. Prog. Ser. 2019, 621, 127–141. [Google Scholar] [CrossRef]
- Djurhuus, A.; Closek, C.J.; Kelly, R.P.; Pitz, K.J.; Michisaki, R.P.; Starks, H.A.; Walz, K.R.; Andruszkiewicz, E.A.; Olesin, E.; Hubbard, K.; et al. Environmental DNA reveals seasonal shifts and potential interactions in a marine community. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Parsons, K.M.; Everett, M.; Dahlheim, M.; Park, L. Water, water everywhere: Environmental DNA can unlock population structure in elusive marine species. R. Soc. Open Sci. 2018, 5, 180537. [Google Scholar] [CrossRef] [Green Version]
- Farrell, J.A.; Yetsko, K.; Whitmore, L.; Whilde, J.; Eastman, C.B.; Ramia, D.R.; Thomas, R.; Linser, P.; Creer, S.; Burkhalter, B.; et al. Environmental DNA monitoring of oncogenic viral shedding and genomic profiling of sea turtle fibropapillo-matosis reveals unusual viral dynamics. Commun Biol. 2021, 4, 565. [Google Scholar] [CrossRef]
- Shea, D.; Bateman, A.; Li, S.; Tabata, A.; Schulze, A.; Mordecai, G.; Ogston, L.; Volpe, J.P.; Frazer, L.N.; Connors, B.; et al. Environmental DNA from multiple pathogens is elevated near active Atlantic salmon farms. Proc. R. Soc. B Boil. Sci. 2020, 287, 20202010. [Google Scholar] [CrossRef] [PubMed]
- Maine State Legislature; Maine Office of Policy and Legal Analysis; Bentley, C.; Schneider, D. Report of the Commission to Study the Effects of Coastal and Ocean Acidification and its Existing and Potential Effects on Species that are Commercially Harvested and Grown Along the Maine Coast. Office of Policy and Legal Analysis. 2015. Available online: https://digitalmaine.com/opla_docs/145 (accessed on 2 February 2022).
- Weinberg, J.R.; Leavitt, D.F.; Lancaster, B.A.; Capuzzo, J.M. Experimental field studies with Mya arenaria (Bivalvia) on the induction and effect of hematopoietic neoplasia. J. Invertebr. Pathol. 1997, 69, 183–194. [Google Scholar] [CrossRef]
- Leavitt, D.F.; Capuzzo, J.M.; Smolowitz, R.M.; Miosky, D.L.; Lancaster, B.A.; Reinisch, C.L. Hematopoietic neoplasia in Mya arenaria: Prevalence and indices of physiological condition. Mar. Biol. 1990, 105, 313–321. [Google Scholar] [CrossRef]
- Burioli, E.A.V.; Hammel, M.; Bierne, N.; Thomas, F.; Houssin, M.; Destoumieux-Garzon, D.; Charriere, G.M. Traits of a mussel transmissible cancer are reminiscent of a parasitic life style. Sci Rep. 2021, 11, 24110. [Google Scholar] [CrossRef] [PubMed]
- Caza, F.; Bernet, E.; Veyrier, F.J.; Betoulle, S.; St-Pierre, Y. Hemocytes released in seawater act as Trojan horses for spreading of bacterial infections in mussels. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Carberry, L.; Roesler, C.; Drapeau, S. Correcting in situ chlorophyll fluorescence time-series observations for nonphotochemical quenching and tidal variability reveals nonconservative phytoplankton variability in coastal waters. Limnol. Oceanogr. Methods 2019, 17, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Renshaw, M.A.; Olds, B.P.; Jerde, C.; McVeigh, M.M.; Lodge, D.M. The room temperature preservation of filtered Environmental DNA samples and assimilation into a phenol-chloroform-isoamyl alcohol DNA extraction. Mol. Ecol. Resour. 2015, 15, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arriagada, G.; Metzger, M.J.; Muttray, A.F.; Sherry, J.; Reinisch, C.; Street, C.; Lipkin, W.I.; Goff, S.P. Activation of transcription and retrotransposition of a novel retroelement, Steamer, in neoplastic hemocytes of the mollusk Mya arenaria. Proc. Natl. Acad. Sci. USA 2014, 111, 14175–14180. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giersch, R.M.; Hart, S.F.M.; Reddy, S.G.; Yonemitsu, M.A.; Orellana Rosales, M.J.; Korn, M.; Geleta, B.M.; Countway, P.D.; Fernández Robledo, J.A.; Metzger, M.J. Survival and Detection of Bivalve Transmissible Neoplasia from the Soft-Shell Clam Mya arenaria (MarBTN) in Seawater. Pathogens 2022, 11, 283. https://doi.org/10.3390/pathogens11030283
Giersch RM, Hart SFM, Reddy SG, Yonemitsu MA, Orellana Rosales MJ, Korn M, Geleta BM, Countway PD, Fernández Robledo JA, Metzger MJ. Survival and Detection of Bivalve Transmissible Neoplasia from the Soft-Shell Clam Mya arenaria (MarBTN) in Seawater. Pathogens. 2022; 11(3):283. https://doi.org/10.3390/pathogens11030283
Chicago/Turabian StyleGiersch, Rachael M., Samuel F. M. Hart, Satyatejas G. Reddy, Marisa A. Yonemitsu, María J. Orellana Rosales, Madelyn Korn, Brook M. Geleta, Peter D. Countway, José A. Fernández Robledo, and Michael J. Metzger. 2022. "Survival and Detection of Bivalve Transmissible Neoplasia from the Soft-Shell Clam Mya arenaria (MarBTN) in Seawater" Pathogens 11, no. 3: 283. https://doi.org/10.3390/pathogens11030283
APA StyleGiersch, R. M., Hart, S. F. M., Reddy, S. G., Yonemitsu, M. A., Orellana Rosales, M. J., Korn, M., Geleta, B. M., Countway, P. D., Fernández Robledo, J. A., & Metzger, M. J. (2022). Survival and Detection of Bivalve Transmissible Neoplasia from the Soft-Shell Clam Mya arenaria (MarBTN) in Seawater. Pathogens, 11(3), 283. https://doi.org/10.3390/pathogens11030283