

Artificial Insemination as an Alternative Transmission Route for African Swine Fever Virus

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
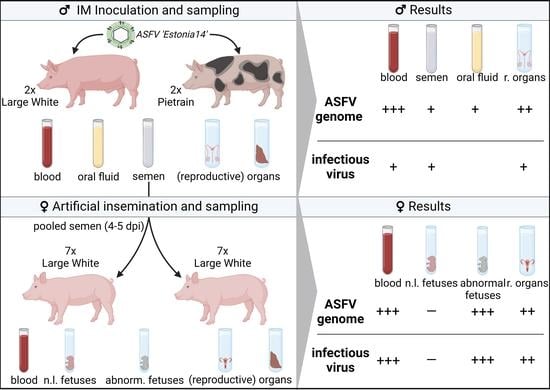
2.1. Experimental Design
2.2. Viruses and Cells
2.3. Sample Collection and Processing
2.4. DNA Extraction and qPCR
2.5. Virus Isolation
2.6. Serology
3. Results
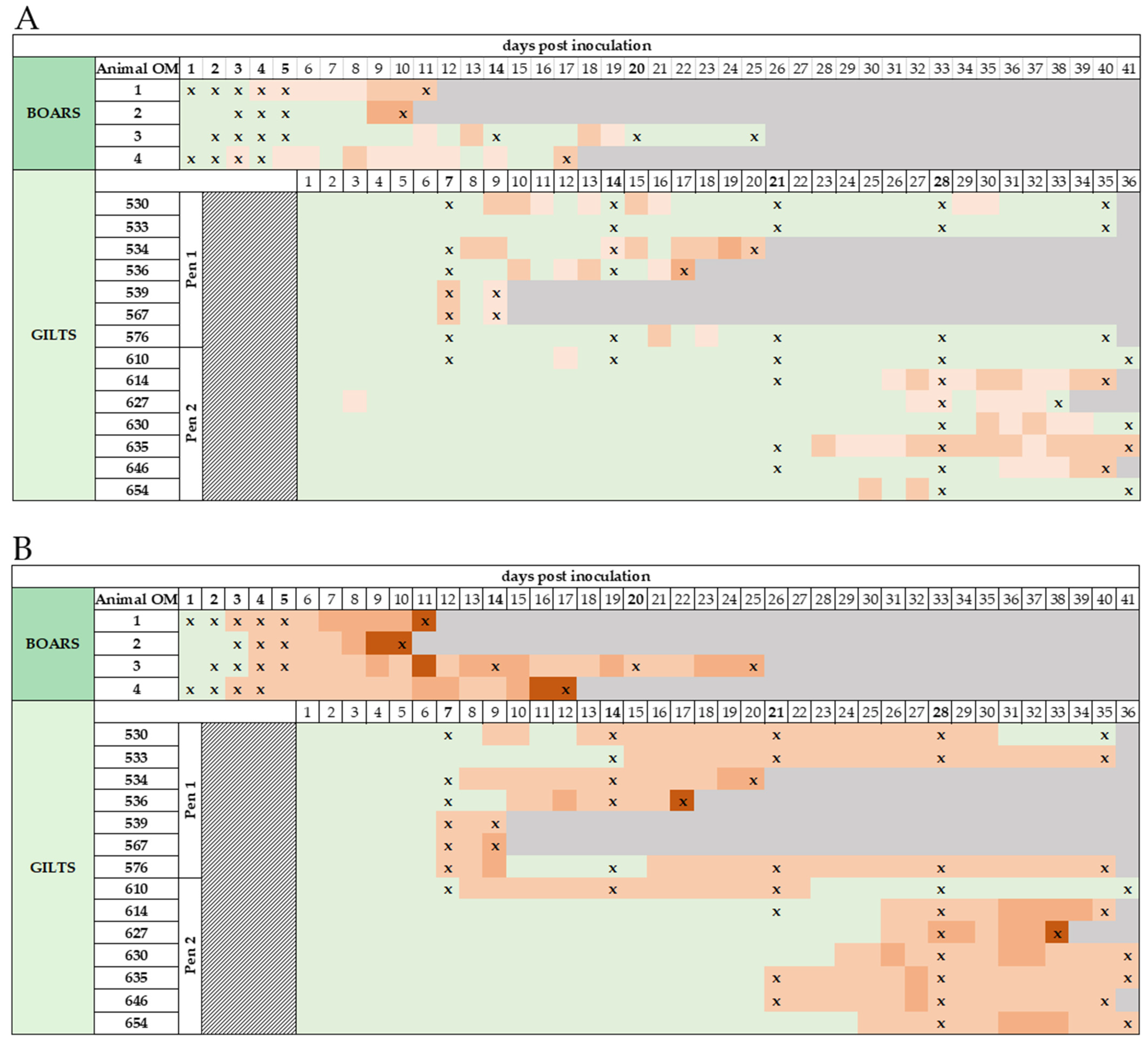
3.1. Clinical Signs, Clinical Scores (CS), and Macroscopic Lesions
3.2. Early Detection of ASFV Genome in Various Matrices Obtained from Boars
3.2.1. Boar Semen and EDTA Blood
3.2.2. Oral Fluids, Dummy Swabs, Serum, and Rectal Samples
3.3. ASFV Genome Load in Organs of Boars and Gilts
3.3.1. Organ Samples Routinely Tested in ASFV Pathogenesis Studies
3.3.2. Reproductive Organs
3.3.3. Embryos and Fetuses
3.4. Assessment of Infectious Virus in Reproductive Organs, Semen and Spleen
3.5. Comparative Serology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallardo, M.C.; Reoyo, A.T.; Fernandez-Pinero, J.; Iglesias, I.; Munoz, M.J.; Arias, M.L. African swine fever: A global view of the current challenge. Porcine Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef] [PubMed]
- Penrith, M.L.; Vosloo, W.; Jori, F.; Bastos, A.D. African swine fever virus eradication in Africa. Virus Res. 2013, 173, 228–246. [Google Scholar] [CrossRef] [PubMed]
- Cappai, S.; Rolesu, S.; Coccollone, A.; Laddomada, A.; Loi, F. Evaluation of biological and socio-economic factors related to persistence of African swine fever in Sardinia. Prev. Vet. Med. 2018, 152, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rolesu, S.; Mandas, D.; Loi, F.; Oggiano, A.; Dei Giudici, S.; Franzoni, G.; Guberti, V.; Cappai, S. African Swine Fever in Smallholder Sardinian Farms: Last 10 Years of Network Transmission Reconstruction and Analysis. Front. Vet. Sci. 2021, 8, 692448. [Google Scholar] [CrossRef] [PubMed]
- Broekhuijse, M.L.; Feitsma, H.; Gadella, B.M. Artificial insemination in pigs: Predicting male fertility. Vet. Q. 2012, 32, 151–157. [Google Scholar] [CrossRef][Green Version]
- Broekhuijse, M.L.; Gaustad, A.H.; Bolarin Guillen, A.; Knol, E.F. Efficient Boar Semen Production and Genetic Contribution: The Impact of Low-Dose Artificial Insemination on Fertility. Reprod. Domest. Anim. 2015, 50 (Suppl. S2), 103–109. [Google Scholar] [CrossRef]
- Waberski, D.; Riesenbeck, A.; Schulze, M.; Weitze, K.F.; Johnson, L. Application of preserved boar semen for artificial insemination: Past, present and future challenges. Theriogenology 2019, 137, 2–7. [Google Scholar] [CrossRef]
- Guerin, C.; Allietta, M.; Guerin, B.; Thibier, M. The use of gene amplification for the detection of the Aujeszky’s disease virus in the sperm of boars. Vet. Res. 1995, 26, 140–144. [Google Scholar]
- Thacker, B.J.; Larsen, R.E.; Joo, H.S.; Leman, A.D. Swine diseases transmissible with artificial insemination. J. Am. Vet. Med. Assoc. 1984, 185, 511–516. [Google Scholar]
- Yaeger, M.J.; Prieve, T.; Collins, J.; Hennings, J.; Nelson, E.; Benfield, D. Evidence for the transmission of porcine reproductive and respiratory syndrome (PRRS) virus in boar semen. Swine Health Prod. 1993, 1, 7–9. [Google Scholar]
- Gradil, C.; Molitor, T.; Harding, M.; Crabo, B. Excretion of porcine parvovirus through the genital tract of boars. Am. J. Vet. Res. 1990, 51, 359–362. [Google Scholar] [PubMed]
- Van Bussel-van Lierop, A.; van Wagenberg, C.; Somers, D. Serbian Pig Sector: An Overview; Fact Finding Mission: Opportunities for Collaboration between Serbia and The Netherlands; Wageningen UR (University & Research Centre) Livestock Research: Wageningen, The Netherlands, 2015; Livestock Research Report 2014. [Google Scholar]
- Roszyk, H.; Franzke, K.; Breithaupt, A.; Deutschmann, P.; Pikalo, J.; Carrau, T.; Blome, S.; Sehl-Ewert, J. The Role of Male Reproductive Organs in the Transmission of African Swine Fever-Implications for Transmission. Viruses 2021, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Pikalo, J.; Schoder, M.E.; Sehl, J.; Breithaupt, A.; Tignon, M.; Cay, A.B.; Gager, A.M.; Fischer, M.; Beer, M.; Blome, S. The African swine fever virus isolate Belgium 2018/1 shows high virulence in European wild boar. Transbound Emerg. Dis. 2020, 67, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Höper, D.; et al. Deletion at the 5’-end of Estonian ASFV strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef] [PubMed]
- Mittelholzer, C.; Moser, C.; Tratschin, J.-D.; Hofmann, M.A. Analysis of classical swine fever virus replication kinetics allows differentiation of highly virulent from avirulent strains. Vet. Microbiol. 2000, 74, 293–308. [Google Scholar] [CrossRef]
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and European wild boar with a Caucasian African swine fever virus isolate. Arch. Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef]
- Galindo-Cardiel, I.; Ballester, M.; Solanes, D.; Nofrarias, M.; Lopez-Soria, S.; Argilaguet, J.M.; Lacasta, A.; Accensi, F.; Rodriguez, F.; Segales, J. Standardization of pathological investigations in the framework of experimental ASFV infections. Virus Res. 2013, 173, 180–190. [Google Scholar] [CrossRef]
- Sehl, J.; Pikalo, J.; Schäfer, A.; Franzke, K.; Pannhorst, K.; Elnagar, A.; Blohm, U.; Blome, S.; Breithaupt, A. Comparative Pathology of Domestic Pigs and Wild Boar Infected with the Moderately Virulent African Swine Fever Virus Strain “Estonia 2014”. Pathogens 2020, 9, 662. [Google Scholar] [CrossRef]
- Carrascosa, A.L.; Bustos, M.J.; de Leon, P. Methods for growing and titrating African swine fever virus: Field and laboratory samples. Curr. Protoc. Cell Biol. 2011, 26, 2614. [Google Scholar] [CrossRef]
- Reicks, D.; Kuster, C.; Rossow, K. PED and boars: Research update. Am. Assoc. Swine Vet. 2015, 21–24. [Google Scholar]
- Schäfer, A.; Zani, L.; Pikalo, J.; Hühr, J.; Sehl, J.; Mettenleiter, T.C.; Breithaupt, A.; Blome, S.; Blohm, U. T-cell responses in domestic pigs and wild boar upon infection with the moderately virulent African swine fever virus strain ’Estonia2014’. Transbound Emerg Dis 2021, 68, 2733–2749. [Google Scholar] [CrossRef] [PubMed]
- Pikalo, J.; Deutschmann, P.; Fischer, M.; Roszyk, H.; Beer, M.; Blome, S. African Swine Fever Laboratory Diagnosis-Lessons Learned from Recent Animal Trials. Pathogens 2021, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.P.; Tian, K.; Nowotny, N. African Swine Fever, the forgotten pandemic. Transbound Emerg Dis 2021, 68, 2637–2639. [Google Scholar] [CrossRef] [PubMed]
- WOAH. African Swine Fever (ASF)—Situation Report; WOAH: Paris, France, 2022; Volume 22. [Google Scholar]
- Knox, R.V. Artificial insemination in pigs today. Theriogenology 2016, 85, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Hennecken, M.; Stegeman, J.A.; Elbers, A.R.; van Nes, A.; Smak, J.A.; Verheijden, J.H. Transmission of classical swine fever virus by artificial insemination during the 1997-1998 epidemic in The Netherlands: A descriptive epidemiological study. Vet. Q. 2000, 22, 228–233. [Google Scholar] [CrossRef]
- Nathues, C.; Perler, L.; Bruhn, S.; Suter, D.; Eichhorn, L.; Hofmann, M.; Nathues, H.; Baechlein, C.; Ritzmann, M.; Palzer, A.; et al. An Outbreak of Porcine Reproductive and Respiratory Syndrome Virus in Switzerland Following Import of Boar Semen. Transbound Emerg. Dis. 2016, 63, e251–e261. [Google Scholar] [CrossRef]
- Reicks, D.; Mun, C.; Rossow, K. Sampling of adult boars during early infection with porcine reproductive and respiratory syndrome virus for testing by polymerase chain reaction using a new blood collection technique (blood-swab method). J. Swine Health Prod. 2006, 14, 258–264. [Google Scholar]
- Christopher-Hennings, J.; Nelson, E.A.; Nelson, J.K.; Hines, R.J.; Swenson, S.L.; Hill, H.T.; Zimmerman, J.J.; Katz, J.B.; Yaeger, M.J.; Chase, C.C.; et al. Detection of porcine reproductive and respiratory syndrome virus in boar semen by PCR. J. Clin. Microbiol. 1995, 33, 1730–1734. [Google Scholar] [CrossRef]
- Schulze, M.; Jung, M.; Hensel, B. Science-based quality control in boar semen production. In Molecular Reproduction and Development; Wiley Online Library: Hoboken, NJ, USA, 2022. [Google Scholar] [CrossRef]
- Nybo, K. qPCR: Technical Replicate Variation. Biotechniques 2011, 50, 23. [Google Scholar] [CrossRef]
- Christopher-Hennings, J.; Nelson, E.A.; Nelson, J.K.; Rossow, K.D.; Shivers, J.L.; Yaeger, M.J.; Chase, C.C.; Garduno, R.A.; Collins, J.E.; Benfield, D.A. Identification of porcine reproductive and respiratory syndrome virus in semen and tissues from vasectomized and nonvasectomized boars. Vet. Pathol. 1998, 35, 260–267. [Google Scholar] [CrossRef]
- de Carvalho Ferreira, H.C.; Weesendorp, E.; Elbers, A.R.; Bouma, A.; Quak, S.; Stegeman, J.A.; Loeffen, W.L. African swine fever virus excretion patterns in persistently infected animals: A quantitative approach. Vet. Microbiol. 2012, 160, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Goonewardene, K.B.; Chung, C.J.; Goolia, M.; Blakemore, L.; Fabian, A.; Mohamed, F.; Nfon, C.; Clavijo, A.; Dodd, K.A.; Ambagala, A. Evaluation of oral fluid as an aggregate sample for early detection of African swine fever virus using four independent pen-based experimental studies. Transbound Emerg. Dis. 2021, 68, 2867–2877. [Google Scholar] [CrossRef]
- SwineReproNet. Swine Reproduction—The Female; University of Illinois: Champaign, IL, USA, 2003. [Google Scholar]
- Pellavio, G.; Todaro, F.; Alberizzi, P.; Scotti, C.; Gastaldi, G.; Lolicato, M.; Omes, C.; Caliogna, L.; Nappi, R.E.; Laforenza, U. HPV Infection Affects Human Sperm Functionality by Inhibition of Aquaporin-8. Cells 2020, 9, 1241. [Google Scholar] [CrossRef] [PubMed]
- Karniychuk, U.U.; Saha, D.; Vanhee, M.; Geldhof, M.; Cornillie, P.; Caij, A.B.; De Regge, N.; Nauwynck, H.J. Impact of a novel inactivated PRRS virus vaccine on virus replication and virus-induced pathology in fetal implantation sites and fetuses upon challenge. Theriogenology 2012, 78, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- McVicar, J.W. Quantitative aspects of the transmission of African swine fever. Am. J. Vet. Res. 1984, 45, 1535–1541. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Matrix | Boars | Gilts | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 530 | 533 | 534 | 536 | 539 | 567 | 576 | 610 | 614 | 627 | 630 | 635 | 646 | 654 | |
| Spleen | •• | ••• | — | •• | •• | •• | •• | •• | ••• | •• | •• | • | •• | •• | •• | •• | • | •• |
| Testis | •• | ••• | — | •• | ||||||||||||||
| Epididymis | •• | •• | — | •• | ||||||||||||||
| Bulbourethral gland | • | •• | — | • | ||||||||||||||
| Vesicular gland | •• | •• | •• | •• | ||||||||||||||
| Prostate | ••• | ••• | •• | •• | ||||||||||||||
| Semen 0 dpi | — | — | — | — | ||||||||||||||
| Semen 2 dpi | •• | — | — | •• | ||||||||||||||
| Semen 3 dpi | •• | — | •• | • | ||||||||||||||
| Semen 4 dpi | ••• | •• | •• | ••• | ||||||||||||||
| Semen 5 dpi | ••• | ••• | •• | |||||||||||||||
| Semen 14 dpi | •• | |||||||||||||||||
| Semen 20 dpi | •• | |||||||||||||||||
| Vagina | • | ••• | •• | •• | ••• | •• | •• | •• | ••• | •• | •• | •• | • | •• | ||||
| Cervix | •• | ••• | ••• | •• | •• | ••• | •• | • | •• | •• | • | •• | •• | •• | ||||
| Uterus | • | •• | ••• | ••• | ••• | •• | ••• | •• | ••• | ••• | •• | •• | •• | •• | ||||
| Salpinx | ••• | ••• | ••• | ••• | ••• | ••• | •• | ••• | •• | •• | •• | •• | •• | •• | ||||
| Ovaries | ••• | ••• | ••• | — | — | ••• | •• | ••• | •• | — | • | •• | •• | ••• | ||||
| ID Screen® ASF Indirect [S/P%] | INgezim PPA COMPAC [%] | IPT | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample [dpi]/OM | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 |
| −3 | — | — | — | — | — | — | — | — |  | |||
| −2 | — | — | — | — | — | — | — | — | ||||
| −1 | — | — | — | — | — | — | — | — | ||||
| 0 | — | — | — | — | — | — | — | — | ||||
| 2 | — | — | — | — | — | — | — | — | ||||
| 3 | — | — | — | — | — | — | — | — | ||||
| 4 | — | — | — | — | — | — | — | — | ||||
| 5 | — | — | — | — | — | — | — | — | ||||
| 10 | 40.4 | 37.6 | — | |||||||||
| 11 | 49.9 | 68.8 | + | |||||||||
| 14 | 60.8 | 57.4 | + | |||||||||
| 17 | 72.7 | 80.0 | + | |||||||||
| 20 | 48.6 | 79.9 | + | |||||||||
| 25 | 58.7 | 86.9 | + | |||||||||
| Pen 1 | Pen 2 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample [dpi]/OM | 530 | 533 | 534 | 536 | 539 | 567 | 576 | 610 | 614 | 627 | 630 | 635 | 646 | 654 | |
| ID Screen® ASF Indirect [S/P%] | 7 | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 9 | 11.1 | 24.7 | |||||||||||||
| 14 | — | — | — | — | — | — | — | — | — | — | — | — | |||
| 17 | 37.3 | ||||||||||||||
| 20 | 54.5 | ||||||||||||||
| 21 | — | — | — | — | — | — | — | — | — | — | |||||
| 28 | 14.9 | 17.5 | 19.3 | 13.4 | — | — | — | — | — | — | |||||
| 33 | 33.4 | ||||||||||||||
| 35 | 71.0 | 66.5 | 71.5 | 55,9 | 60.7 | ||||||||||
| 36 | 69.8 | 49.0 | 63.5 | 52.0 | |||||||||||
| INgezim PPA COMPAC [%] | 7 | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 9 | 18.9 | 14.4 | |||||||||||||
| 14 | — | — | — | — | — | — | — | — | — | — | — | — | |||
| 17 | 56.5 | ||||||||||||||
| 20 | 71.9 | ||||||||||||||
| 21 | — | — | — | — | — | — | — | — | — | — | |||||
| 28 | 54.5 | 57.1 | 61.7 | 50.2 | — | — | — | — | — | — | |||||
| 33 | 85.9 | ||||||||||||||
| 35 | 90.4 | 91.1 | 96.1 | 69.2 | 64.8 | ||||||||||
| 36 | 98.0 | 79.4 | 71.1 | 67.1 | |||||||||||
| IPT | 7 |  | | | | | | | | | | | | | |
| 9 | — | — | |||||||||||||
| 14 | | | | | | | | | | | | | | | |
| 17 | + | ||||||||||||||
| 20 | + | ||||||||||||||
| 21 | | | | | | | | | | | | | | | |
| 28 | + | + | + | + | | | | | | | |||||
| 33 | + | ||||||||||||||
| 35 | + | + | + | + | + | ||||||||||
| 36 | + | + | + | + | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedrichs, V.; Reicks, D.; Hasenfuß, T.; Gerstenkorn, E.; Zimmerman, J.J.; Nelson, E.A.; Carrau, T.; Deutschmann, P.; Sehl-Ewert, J.; Roszyk, H.; et al. Artificial Insemination as an Alternative Transmission Route for African Swine Fever Virus. Pathogens 2022, 11, 1539. https://doi.org/10.3390/pathogens11121539
Friedrichs V, Reicks D, Hasenfuß T, Gerstenkorn E, Zimmerman JJ, Nelson EA, Carrau T, Deutschmann P, Sehl-Ewert J, Roszyk H, et al. Artificial Insemination as an Alternative Transmission Route for African Swine Fever Virus. Pathogens. 2022; 11(12):1539. https://doi.org/10.3390/pathogens11121539
Chicago/Turabian StyleFriedrichs, Virginia, Darwin Reicks, Tobias Hasenfuß, Elisabeth Gerstenkorn, Jeffrey J. Zimmerman, Eric A. Nelson, Tessa Carrau, Paul Deutschmann, Julia Sehl-Ewert, Hanna Roszyk, and et al. 2022. "Artificial Insemination as an Alternative Transmission Route for African Swine Fever Virus" Pathogens 11, no. 12: 1539. https://doi.org/10.3390/pathogens11121539
APA StyleFriedrichs, V., Reicks, D., Hasenfuß, T., Gerstenkorn, E., Zimmerman, J. J., Nelson, E. A., Carrau, T., Deutschmann, P., Sehl-Ewert, J., Roszyk, H., Beer, M., Christopher-Hennings, J., & Blome, S. (2022). Artificial Insemination as an Alternative Transmission Route for African Swine Fever Virus. Pathogens, 11(12), 1539. https://doi.org/10.3390/pathogens11121539