
Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine

 ,
, 
 , ,
, ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
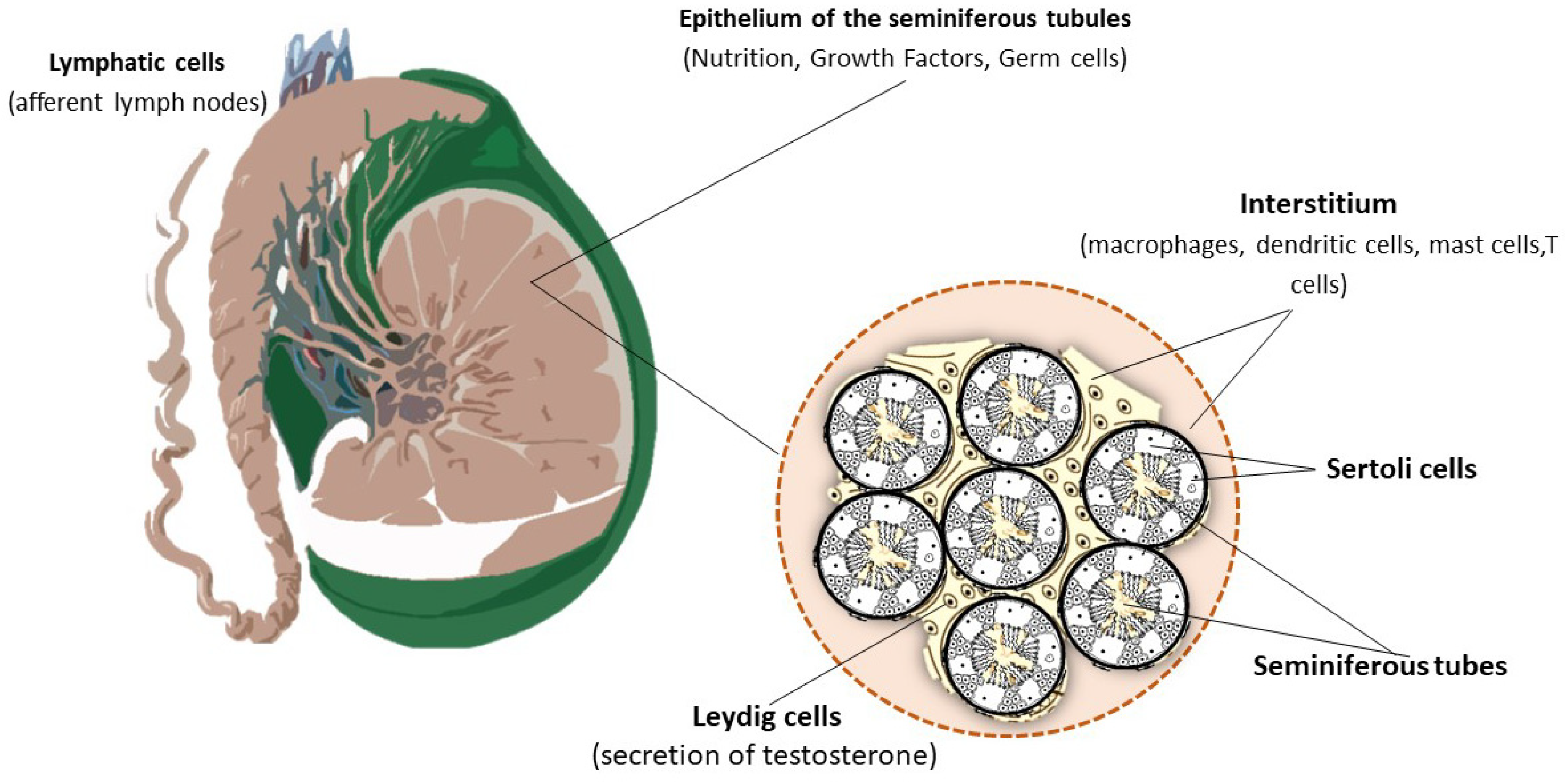
1.1. The Immune Environment of the Testis
1.2. Blood–Testis Barrier Function
1.3. The Immune Arsenal of the Testis
- (a)
- Macrophages
- (b)
- Dendritic Cells
- (c)
- T Cells
1.4. Composition and Function of the Testicular Microbiota
1.5. Seminal Dysbiosis with Particular Reference to Male Infertility
1.6. Microbial-Mediated Male Infertily
1.7. Therapeutic Correction of Testicular Dysbiosis with Natural Products
- (a)
- Probiotics
- (b)
- Prebiotics
- (c)
- Antioxidants
- (d)
- Polyphenols
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Barbăroșie, C.; Ambar, R.; Finelli, R. The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction. Int. J. Mol. Sci. 2020, 21, 3882. [Google Scholar] [CrossRef] [PubMed]
- Hermann, B.P.; Cheng, K.; Singh, A.; Roa-De La Cruz, L.; Mutoji, K.N.; Chen, I.C.; Gildersleeve, H.; Lehle, J.D.; Mayo, M.; Westernströer, B.; et al. The Mammalian Spermatogenesis Single-Cell Transcriptome, from Spermatogonial Stem Cells to Spermatids. Cell Rep. 2018, 25, 1650–1667.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Zeng, Q.; Ling, M.; Ma, R.; Chen, H.; Lin, F.; Li, Z.; Pan, L. Inhibition of IP3R/Ca2+ Dysregulation Protects Mice from Ventilator-Induced Lung Injury via Endoplasmic Reticulum and Mitochondrial Pathways. Front. Immunol. 2021, 12, 729094. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, A.; Wang, M.; Schulz, C.; Bhushan, S. Microenvironmental signals govern the cellular identity of testicular macrophages. J. Leukoc. Biol. 2018, 104, 757–766. [Google Scholar] [CrossRef]
- Forrester, J.V.; Xu, H.; Lambe, T.; Cornall, R. Immune privilege or privileged immunity? Mucosal Immunol. 2008, 1, 372–381. [Google Scholar] [CrossRef]
- O’Donnell, L.; Smith, L.B.; Rebourcet, D. Sperm-specific proteins: New implications for diagnostic development and cancer immunotherapy. Curr. Opin. Cell Biol. 2022, 77, 102104. [Google Scholar] [CrossRef]
- Stanton, P.G. Regulation of the blood-testis barrier. Semin. Cell Dev. Biol. 2016, 59, 166–173. [Google Scholar] [CrossRef]
- Schuppe, H.C.; Meinhardt, A. Immune privilege and inflammation of the testis. Immunol. Gametes Embryo Implant. 2005, 88, 1–14. [Google Scholar] [CrossRef]
- Chen, Q.; Deng, T.; Han, D. Testicular immunoregulation and spermatogenesis. Semin. Cell Dev. Biol. 2016, 59, 157–165. [Google Scholar] [CrossRef]
- Loveland, K.L.; Klein, B.; Pueschl, D.; Indumathy, S.; Bergmann, M.; Loveland, B.E.; Hedger, M.P.; Schuppe, H.C. Cytokines in Male Fertility and Reproductive Pathologies: Immunoregulation and Beyond. Front. Endocrinol. 2017, 8, 307. [Google Scholar] [CrossRef]
- Proctor, L. Priorities for the next 10 years of human microbiome research. Nature 2019, 569, 623–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmet, L.; Thijs, T.; Segers, A.; Verbeke, K.; Depoortere, I. Chronodisruption by chronic jetlag impacts metabolic and gastrointestinal homeostasis in male mice. Acta Physiol. 2021, 233, e13703. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Lehmann, C.J.; Cole, C.G.; Pamer, E.G. Translating Microbiome Research from and to the Clinic. Annu. Rev. Microbiol. 2022. [Google Scholar] [CrossRef]
- Baptissart, M.; Vega, A.; Martinot, E.; Pommier, A.J.; Houten, S.M.; Marceau, G.; de Haze, A.; Baron, S.; Schoonjans, K.; Lobaccaro, J.M.; et al. Bile acids alter male fertility through G-protein-coupled bile acid receptor 1 signaling pathways in mice. Hepatology 2014, 60, 1054–1065. [Google Scholar] [CrossRef]
- Sonowal, R.; Swimm, A.; Sahoo, A.; Luo, L.; Matsunaga, Y.; Wu, Z.; Bhingarde, J.A.; Ejzak, E.A.; Ranawade, A.; Qadota, H.; et al. Indoles from commensal bacteria extend healthspan. Proc. Natl. Acad. Sci. USA 2017, 114, E7506–E7515. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, V.; Zakroczymski, M.A.; Naaz, A.; Mukai, M.; Ju, Y.H.; Doerge, D.R.; Katzenellenbogen, J.A.; Helferich, W.G.; Cooke, P.S. Estrogenicity of the isoflavone metabolite equol on reproductive and non-reproductive organs in mice. Biol. Reprod. 2004, 71, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Nobs, S.P.; Zmora, N.; Elinav, E. Nutrition Regulates Innate Immunity in Health and Disease. Annu. Rev. Nutr. 2020, 40, 189–219. [Google Scholar] [CrossRef]
- Lundy, S.D.; Vij, S.C.; Eng, C. Reply to Eugenio Ventimiglia, EdoardoPozzi, Massimo Alfano, Francesco Montorsi, and Andrea Salonia’s Letter to the Editor re: Scott D. Lundy, Naseer Sangwan, Neel V. Parekh; et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur Urol 2021;79:826–36. Eur. Urol. 2021, 80, e55–e56. [Google Scholar] [CrossRef]
- Kaur, G.; Thompson, L.A.; Dufour, J.M. Sertoli cells—Immunological sentinels of spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Wang, T.; Han, D. Structural, cellular and molecular aspects of immune privilege in the testis. Front. Immunol. 2012, 3, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruthig, V.A.; Lamb, D.J. Updates in Sertoli Cell-Mediated Signaling During Spermatogenesis and Advances in Restoring Sertoli Cell Function. Front. Endocrinol. 2022, 13, 897196. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, A.; Hedger, M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol. Cell. Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef]
- Gu, X.; Li, S.Y.; Matsuyama, S.; DeFalco, T. Immune Cells as Critical Regulators of Steroidogenesis in the Testis and Beyond. Front. Endocrinol. 2022, 13, 894437. [Google Scholar] [CrossRef]
- Fijak, M.; Bhushan, S.; Meinhardt, A. Immunoprivileged sites: The testis. Methods Mol. Biol. 2011, 677, 459–470. [Google Scholar] [CrossRef]
- Hirai, S.; Naito, M.; Terayama, H.; Qu, N.; Kuerban, M.; Musha, M.; Ikeda, A.; Miura, M.; Itoh, M. The origin of lymphatic capillaries in murine testes. J. Androl. 2012, 33, 745–751. [Google Scholar] [CrossRef]
- Su, L.; Wang, Z.; Xie, S.; Hu, D.; Cheng, Y.C.; Mruk, D.D.; Guan, Y. Testin regulates the blood-testis barrier via disturbing occludin/ZO-1 association and actin organization. J. Cell. Physiol. 2020, 235, 6127–6138. [Google Scholar] [CrossRef]
- Mital, P.; Hinton, B.T.; Dufour, J.M. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol. Reprod. 2011, 84, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef]
- Ye, L.; Huang, W.; Liu, S.; Cai, S.; Hong, L.; Xiao, W.; Thiele, K.; Zeng, Y.; Song, M.; Diao, L. Impacts of Immunometabolism on Male Reproduction. Front. Immunol. 2021, 12, 658432. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Pinzon, W.; Korbutt, G.S.; Power, R.; Hooton, J.; Rajotte, R.V.; Rabinovitch, A. Testicular sertoli cells protect islet beta-cells from autoimmune destruction in NOD mice by a transforming growth factor-beta1-dependent mechanism. Diabetes 2000, 49, 1810–1818. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, Y.; Shiratsuchi, A. Phagocytic removal of apoptotic spermatogenic cells by Sertoli cells: Mechanisms and consequences. Biol. Pharm. Bull. 2004, 27, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative Stress, Testicular Inflammatory Pathways, and Male Reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhu, W.; Xue, S.; Han, D. Testicular defense systems: Immune privilege and innate immunity. Cell. Mol. Immunol. 2014, 11, 428–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; Korrovits, P.; Laan, M. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 2017, 32, 18–31. [Google Scholar] [CrossRef]
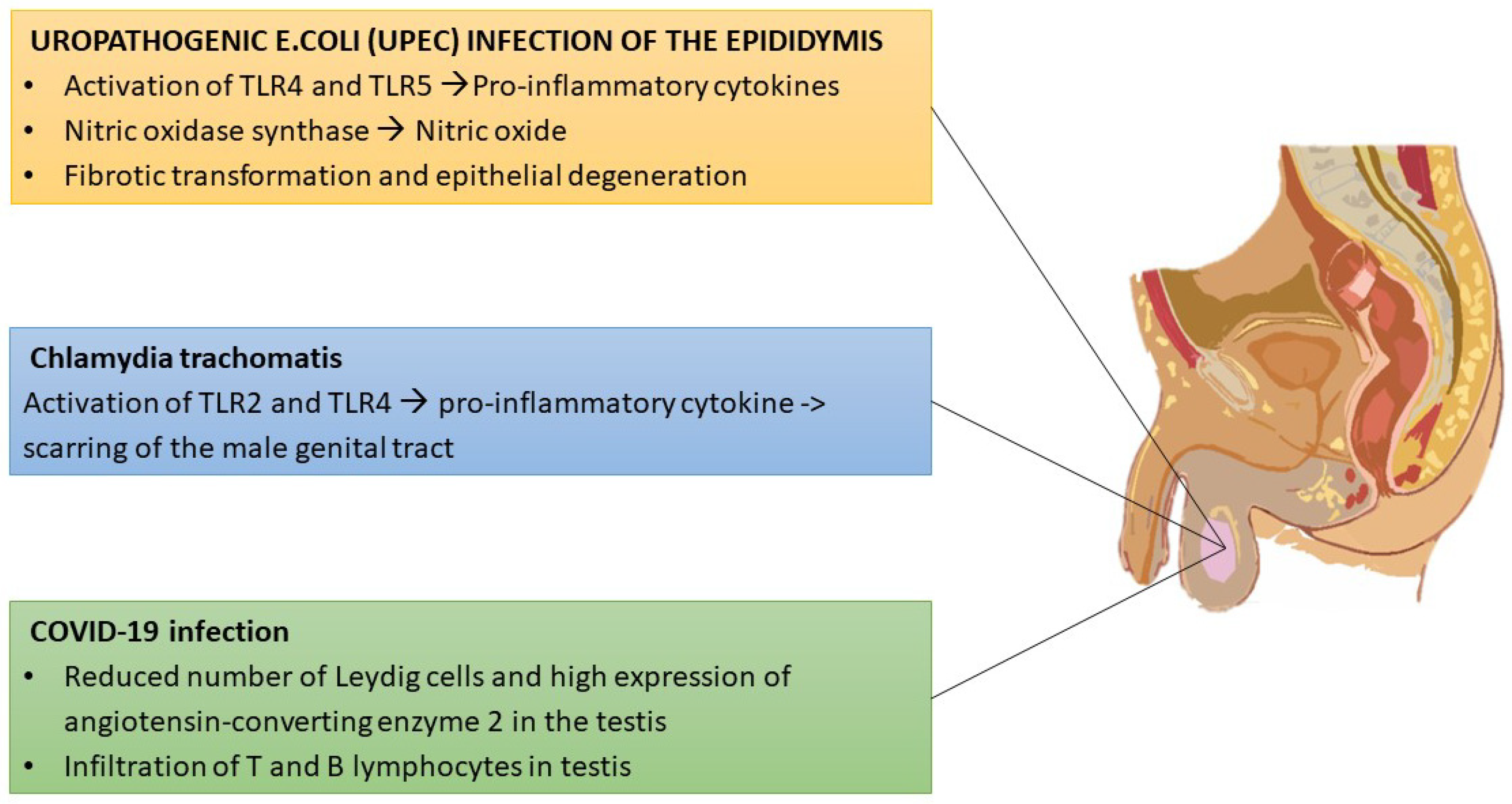
- Michel, V.; Duan, Y.; Stoschek, E.; Bhushan, S.; Middendorff, R.; Young, J.M.; Loveland, K.L.; Kretser, D.M.; Hedger, M.P.; Meinhardt, A. Uropathogenic Escherichia coli causes fibrotic remodelling of the epididymis. J. Pathol. 2016, 240, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Schuppe, H.C.; Pilatz, A.; Hossain, H.; Diemer, T.; Wagenlehner, F.; Weidner, W. Urogenital Infection as a Risk Factor for Male Infertility. Dtsch. Ärzteblatt Int. 2017, 114, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Hedger, M.P. Immunophysiology and pathology of inflammation in the testis and epididymis. J. Androl. 2011, 32, 625–640. [Google Scholar] [CrossRef]
- Gray, J.I.; Farber, D.L. Tissue-Resident Immune Cells in Humans. Annu. Rev. Immunol. 2022, 40, 195–220. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Tuganbaev, T.; Skelly, A.N.; Honda, K. T Cell Responses to the Microbiota. Annu. Rev. Immunol. 2022, 40, 559–587. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Meinhardt, A.; Dejucq-Rainsford, N.; Bhushan, S. Testicular macrophages: Development and function in health and disease. Trends Immunol. 2022, 43, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fijak, M.; Hossain, H.; Markmann, M.; Nüsing, R.M.; Lochnit, G.; Hartmann, M.F.; Wudy, S.A.; Zhang, L.; Gu, H.; et al. Characterization of the Micro-Environment of the Testis that Shapes the Phenotype and Function of Testicular Macrophages. J. Immunol. 2017, 198, 4327–4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleuger, C.; Silva, E.J.R.; Pilatz, A.; Bhushan, S.; Meinhardt, A. Differential Immune Response to Infection and Acute Inflammation Along the Epididymis. Front. Immunol. 2020, 11, 599594. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.R.; Atiyah, S.A.; Yser, H.T.; Khelewe, A.M.; Hameed, H.N. The influence of SARS-CoV-2 on semen parameters of infected infertile male in comparison with those that noninfected. J. Clin. Lab. Anal. 2022, e24568. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Acklin, J.A.; Liu, G.; Kenney, H.; Teterina, N.L.; Pletnev, A.G.; Lim, J.K. Zika virus tropism during early infection of the testicular interstitium and its role in viral pathogenesis in the testes. PLoS Pathog. 2020, 16, e1008601. [Google Scholar] [CrossRef]
- Bhushan, S.; Meinhardt, A. The macrophages in testis function. J. Reprod. Immunol. 2017, 119, 107–112. [Google Scholar] [CrossRef]
- Jarazo-Dietrich, S.; Jacobo, P.; Pérez, C.V.; Guazzone, V.A.; Lustig, L.; Theas, M.S. Up regulation of nitric oxide synthase-nitric oxide system in the testis of rats undergoing autoimmune orchitis. Immunobiology 2012, 217, 778–787. [Google Scholar] [CrossRef]
- De Rose, R.; Fernandez, C.S.; Hedger, M.P.; Kent, S.J.; Winnall, W.R. Characterisation of macaque testicular leucocyte populations and T-lymphocyte immunity. J. Reprod. Immunol. 2013, 100, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Duan, Y.G. The role of dendritic cells in male reproductive tract. Am. J. Reprod. Immunol. 2016, 76, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Gualdoni, G.S.; Jacobo, P.V.; Sobarzo, C.M.; Pérez, C.V.; Matzkin, M.E.; Höcht, C.; Frungieri, M.B.; Hill, M.; Anegon, I.; Lustig, L.; et al. Role of indoleamine 2,3-dioxygenase in testicular immune-privilege. Sci. Rep. 2019, 9, 15919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [Green Version]
- Pallotta, M.T.; Orabona, C.; Volpi, C.; Vacca, C.; Belladonna, M.L.; Bianchi, R.; Servillo, G.; Brunacci, C.; Calvitti, M.; Bicciato, S.; et al. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat. Immunol. 2011, 12, 870–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, D.H.; Mellor, A.L. Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J. Clin. Investig. 2007, 117, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Fijak, M.; Meinhardt, A. The testis in immune privilege. Immunol. Rev. 2006, 213, 66–81. [Google Scholar] [CrossRef]
- Jacobo, P.; Guazzone, V.A.; Theas, M.S.; Lustig, L. Testicular autoimmunity. Autoimmun. Rev. 2011, 10, 201–204. [Google Scholar] [CrossRef]
- Garza, K.M.; Agersborg, S.S.; Baker, E.; Tung, K.S. Persistence of physiological self antigen is required for the regulation of self tolerance. J. Immunol. 2000, 164, 3982–3989. [Google Scholar] [CrossRef] [Green Version]
- Kekäläinen, E.; Tuovinen, H.; Joensuu, J.; Gylling, M.; Franssila, R.; Pöntynen, N.; Talvensaari, K.; Perheentupa, J.; Miettinen, A.; Arstila, T.P. A defect of regulatory T cells in patients with autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy. J. Immunol. 2007, 178, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.G.; Yu, C.F.; Novak, N.; Bieber, T.; Zhu, C.H.; Schuppe, H.C.; Haidl, G.; Allam, J.P. Immunodeviation towards a Th17 immune response associated with testicular damage in azoospermic men. Int. J. Androl. 2011, 34 Pt 2, e536–e545. [Google Scholar] [CrossRef] [PubMed]
- Jacobo, P.; Guazzone, V.A.; Jarazo-Dietrich, S.; Theas, M.S.; Lustig, L. Differential changes in CD4+ and CD8+ effector and regulatory T lymphocyte subsets in the testis of rats undergoing autoimmune orchitis. J. Reprod. Immunol. 2009, 81, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zeng, Q.; Yu, D.; Duan, Y.G. T Lymphocytes and Testicular Immunity: A New Insight into Immune Regulation in Testes. Int. J. Mol. Sci. 2020, 22, 57. [Google Scholar] [CrossRef] [PubMed]
- Bhati, M.; Llamosas, E.; Jacques, D.A.; Jeffries, C.M.; Dastmalchi, S.; Ripin, N.; Nicholas, H.R.; Matthews, J.M. Interactions between LHX3- and ISL1-family LIM-homeodomain transcription factors are conserved in Caenorhabditis elegans. Sci. Rep. 2017, 7, 4579. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, S.; Theas, M.S.; Guazzone, V.A.; Jacobo, P.; Wang, M.; Fijak, M.; Meinhardt, A.; Lustig, L. Immune Cell Subtypes and Their Function in the Testis. Front. Immunol. 2020, 11, 583304. [Google Scholar] [CrossRef]
- Hedger, M.P.; Meinhardt, A. Local regulation of T cell numbers and lymphocyte-inhibiting activity in the interstitial tissue of the adult rat testis. J. Reprod. Immunol. 2000, 48, 69–80. [Google Scholar] [CrossRef]
- Dai, Z.; Nasr, I.W.; Reel, M.; Deng, S.; Diggs, L.; Larsen, C.P.; Rothstein, D.M.; Lakkis, F.G. Impaired recall of CD8 memory T cells in immunologically privileged tissue. J. Immunol. 2005, 174, 1165–1170. [Google Scholar] [CrossRef] [Green Version]
- Nasr, I.W.; Wang, Y.; Gao, G.; Deng, S.; Diggs, L.; Rothstein, D.M.; Tellides, G.; Lakkis, F.G.; Dai, Z. Testicular immune privilege promotes transplantation tolerance by altering the balance between memory and regulatory T cells. J. Immunol. 2005, 174, 6161–6168. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Dai, H.; Wan, N.; Moore, Y.; Vankayalapati, R.; Dai, Z. Interaction of programmed death-1 and programmed death-1 ligand-1 contributes to testicular immune privilege. Transplantation 2009, 87, 1778–1786. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, J.; Liu, B.; Jiang, Y.; Chen, W.; Li, J.; He, Q.; He, Z. The roles and mechanisms of Leydig cells and myoid cells in regulating spermatogenesis. Cell. Mol. Life Sci. 2019, 76, 2681–2695. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Saari, T.; Salmi, T.T.; Pöllänen, P.; Pelliniemi, L.J. Reactions of Leydig cells and blood vessels to lymphoblastic leukemia in the rat testis. Leukemia 1995, 9, 908–914. [Google Scholar] [PubMed]
- Sainio-Pöllänen, S.; Sundström, J.; Erkkilä, S.; Hänninen, A.; Vainiopää, M.; Martikainen, M.; Salminen, E.; Veräjänkorva, E.; Antola, H.; Nikula, H.; et al. CD106 (VCAM-1) in testicular immunoregulation. J. Reprod. Immunol. 1997, 33, 221–238. [Google Scholar] [CrossRef]
- Dutta, D.; Park, I.; Guililat, H.; Sang, S.; Talapatra, A.; Hanson, L.; Mills, N.C. Ethylene dimethane sulfonate (EDS) ablation of Leydig cells in adult rat depletes testosterone resulting in epididymal sperm granuloma: Testosterone replacement prevents granuloma formation. Reprod. Biol. 2019, 19, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Schneider, E.; Klug, J.; Bhushan, S.; Hackstein, H.; Schuler, G.; Wygrecka, M.; Gromoll, J.; Meinhardt, A. Testosterone replacement effectively inhibits the development of experimental autoimmune orchitis in rats: Evidence for a direct role of testosterone on regulatory T cell expansion. J. Immunol. 2011, 186, 5162–5172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monsivais, D.; Matzuk, M.M.; Pangas, S.A. The TGF-β Family in the Reproductive Tract. Cold Spring Harb. Perspect. Biol. 2017, 9, a022251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Secco, V.; Riccioli, A.; Padula, F.; Ziparo, E.; Filippini, A. Mouse Sertoli cells display phenotypical and functional traits of antigen-presenting cells in response to interferon gamma. Biol. Reprod. 2008, 78, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Fallarino, F.; Luca, G.; Calvitti, M.; Mancuso, F.; Nastruzzi, C.; Fioretti, M.C.; Grohmann, U.; Becchetti, E.; Burgevin, A.; Kratzer, R.; et al. Therapy of experimental type 1 diabetes by isolated Sertoli cell xenografts alone. J. Exp. Med. 2009, 206, 2511–2526. [Google Scholar] [CrossRef] [Green Version]
- Alfano, M.; Ferrarese, R.; Locatelli, I.; Ventimiglia, E.; Ippolito, S.; Gallina, P.; Cesana, D.; Canducci, F.; Pagliardini, L.; Viganò, P.; et al. Testicular microbiome in azoospermic men-first evidence of the impact of an altered microenvironment. Hum. Reprod. 2018, 33, 1212–1217. [Google Scholar] [CrossRef]
- Molina, N.M.; Plaza-Díaz, J.; Vilchez-Vargas, R.; Sola-Leyva, A.; Vargas, E.; Mendoza-Tesarik, R.; Galán-Lázaro, M.; Mendoza-Ladrón de Guevara, N.; Tesarik, J.; Altmäe, S. Assessing the testicular sperm microbiome: A low-biomass site with abundant contamination. Reprod. Biomed. Online 2021, 43, 523–531. [Google Scholar] [CrossRef]
- Yang, H.; Büttner, A.; Albiol, L.; Julien, C.; Thiele, T.; Figge, C.; Kramer, I.; Kneissel, M.; Duda, G.N.; Checa, S.; et al. Cortical bone adaptation to a moderate level of mechanical loading in male Sost deficient mice. Sci. Rep. 2020, 10, 22299. [Google Scholar] [CrossRef]
- Lundy, S.D.; Sangwan, N.; Parekh, N.V.; Selvam, M.K.P.; Gupta, S.; McCaffrey, P.; Bessoff, K.; Vala, A.; Agarwal, A.; Sabanegh, E.S.; et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur. Urol. 2021, 79, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Wilharm, A.; Brigas, H.C.; Sandrock, I.; Ribeiro, M.; Amado, T.; Reinhardt, A.; Demera, A.; Hoenicke, L.; Strowig, T.; Carvalho, T.; et al. Microbiota-dependent expansion of testicular IL-17-producing Vγ6+γδ T cells upon puberty promotes local tissue immune surveillance. Mucosal Immunol. 2021, 14, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Gachet, C.; Prat, M.; Burucoa, C.; Grivard, P.; Pichon, M. Spermatic Microbiome Characteristics in Infertile Patients: Impact on Sperm Count, Mobility, and Morphology. J. Clin. Med. 2022, 11, 1505. [Google Scholar] [CrossRef] [PubMed]
- Charitos, I.A.; Topi, S.; Gagliano-Candela, R.; De Nitto, E.; Polimeno, L.; Montagnani, M.; Santacroce, L. The toxic effects of endocrine disrupting chemicals (EDCs) on gut microbiota: Bisphenol A (BPA). A review. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 716–727. [Google Scholar] [CrossRef]
- Santacroce, L.; Bottalico, L.; Topi, S.; Castellaneta, F.; Charitos, I.A. The “Scourge of the Renaissance”. A Short Review about Treponema pallidum infection. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 335–343. [Google Scholar] [CrossRef]
- Alfano, M.; Pederzoli, F.; Locatelli, I.; Ippolito, S.; Longhi, E.; Zerbi, P.; Ferrari, M.; Brendolan, A.; Montorsi, F.; Drago, D.; et al. Impaired testicular signaling of vitamin A and vitamin K contributes to the aberrant composition of the extracellular matrix in idiopathic germ cell aplasia. Fertil. Steril. 2019, 111, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.Y.; Li, R.J.W.; Lim, Y.M.; Batchuluun, B.; Liu, H.; Waise, T.M.Z.; Lam, T.K.T. FXR in the dorsal vagal complex is sufficient and necessary for upper small intestinal microbiome-mediated changes of TCDCA to alter insulin action in rats. Gut 2021, 70, 1675–1683. [Google Scholar] [CrossRef]
- Santacroce, L.; Man, A.; Charitos, I.A.; Haxhirexha, K.; Topi, S. Current knowledge about the connection between health status and gut microbiota from birth to elderly. A narrative review. Front. Biosci. 2021, 26, 135–148. [Google Scholar] [CrossRef]
- Ding, N.; Zhang, X.; Zhang, X.D.; Jing, J.; Liu, S.S.; Mu, Y.P.; Peng, L.L.; Yan, Y.J.; Xiao, G.M.; Bi, X.Y.; et al. Impairment of spermatogenesis and sperm motility by the high-fat diet-induced dysbiosis of gut microbes. Gut 2020, 69, 1608–1619. [Google Scholar] [CrossRef] [Green Version]
- Tremellen, K. Gut Endotoxin Leading to a Decline IN Gonadal function (GELDING)—A novel theory for the development of late onset hypogonadism in obese men. Basic Clin. Androl. 2016, 26, 7. [Google Scholar] [CrossRef] [Green Version]
- Sonnex, C. Toll-like receptors and genital tract infection. Int. J. STD AIDS 2010, 21, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Wegierska, A.E.; Charitos, I.A.; Topi, S.; Potenza, M.A.; Montagnani, M.; Santacroce, L. The Connection between Physical Exercise and Gut Microbiota, Implications for Competitive Sports Athletes. Sports Med. 2022; ahead of print. [Google Scholar] [CrossRef]
- Sarkar, O.; Bahrainwala, J.; Chandrasekaran, S.; Kothari, S.; Mathur, P.P.; Agarwal, A. Impact of inflammation on male fertility. Front. Biosci. 2011, 3, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.G.; Sharma, R.K.; Ollero, M.; Saleh, R.A.; Lopez, M.C.; Thomas, A.J., Jr.; Evenson, D.P.; Agarwal, A. Increased DNA damage in sperm from leukocytospermic semen samples as determined by the sperm chromatin structure assay. Fertil. Steril. 2002, 78, 319–329. [Google Scholar] [CrossRef]
- Zheng, W.; Wu, H.; Liu, C.; Yan, Q.; Wang, T.; Wu, P.; Liu, X.; Jiang, Y.; Zhan, S. Identification of COVID-19 and Dengue Host Factor Interaction Networks Based on Integrative Bioinformatics Analyses. Front. Immunol. 2021, 12, 707287. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, A.S.; Larsen, N.; Pedersen-Skovsgaard, T.; Berg, R.M.; Møller, K.; Svendsen, K.D.; Jakobsen, M.; Pedersen, B.K. Effects of Lactobacillus acidophilus NCFM on insulin sensitivity and the systemic inflammatory response in human subjects. Br. J. Nutr. 2010, 104, 1831–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkesworth, S.; Moore, S.E.; Fulford, A.J.; Barclay, G.R.; Darboe, A.A.; Mark, H.; Nyan, O.A.; Prentice, A.M. Evidence for metabolic endotoxemia in obese and diabetic Gambian women. Nutr. Diabetes 2013, 3, e83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, A.; Ciurea, C.N.; Pasaroiu, D.; Savin, A.I.; Toma, F.; Sular, F.; Santacroce, L.; Mare, A. New perspectives on the nutritional factors influencing growth rate of Candida albicans in diabetics. An in vitro study. MemóriasDo Inst. Oswaldo Cruz 2017, 112, 587–592. [Google Scholar] [CrossRef]
- Zhu, Y.Z.; Sun, H.; Fu, Y.; Wang, J.; Song, M.; Li, M.; Li, Y.F.; Miao, L.G. Effects of sub-chronic aluminum chloride on spermatogenesis and testicular enzymatic activity in male rats. Life Sci. 2014, 102, 36–40. [Google Scholar] [CrossRef]
- Pitteloud, N.; Hardin, M.; Dwyer, A.A.; Valassi, E.; Yialamas, M.; Elahi, D.; Hayes, F.J. Increasing insulin resistance is associated with a decrease in Leydig cell testosterone secretion in men. J. Clin. Endocrinol. Metab. 2005, 90, 2636–2641. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, C.; Marques, P.I.; Cavadas, B.; Damião, I.; Almeida, V.; Barros, N.; Barros, A.; Carvalho, F.; Gomes, S.; Seixas, S. Characterization of microbiota in male infertility cases uncovers differences in seminal hyperviscosity and oligoasthenoteratozoospermia possibly correlated with increased prevalence of infectious bacteria. Am. J. Reprod. Immunol. 2018, 79, e12838. [Google Scholar] [CrossRef]
- Zhao, L.; Yao, C.; Xing, X.; Jing, T.; Li, P.; Zhu, Z.; Yang, C.; Zhai, J.; Tian, R.; Chen, H.; et al. Single-cell analysis of developing and azoospermia human testicles reveals central role of Sertoli cells. Nat. Commun. 2020, 11, 5683. [Google Scholar] [CrossRef] [PubMed]
- Mändar, R.; Türk, S.; Korrovits, P.; Ausmees, K.; Punab, M. Impact of sexual debut on culturable human seminal microbiota. Andrology 2018, 6, 510–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, S.L.; Chiu, C.M.; Lin, F.M.; Huang, W.C.; Liang, C.; Yang, T.; Yang, T.L.; Liu, C.Y.; Wu, W.Y.; Chang, Y.A.; et al. Bacterial communities in semen from men of infertile couples: Metagenomic sequencing reveals relationships of seminal microbiota to semen quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [Green Version]
- Mändar, R.; Punab, M.; Korrovits, P.; Türk, S.; Ausmees, K.; Lapp, E.; Preem, J.K.; Oopkaup, K.; Salumets, A.; Truu, J. Seminal microbiome in men with and without prostatitis. Int. J. Urol. 2017, 24, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConaghy, J.R.; Panchal, B. Epididymitis: An Overview. Am. Fam. Physician 2016, 94, 723–726. [Google Scholar] [PubMed]
- Fijak, M.; Pilatz, A.; Hedger, M.P.; Nicolas, N.; Bhushan, S.; Michel, V.; Tung, K.S.K.; Schuppe, H.C.; Meinhardt, A. Infectious, inflammatory and ‘autoimmune’ male factor infertility: How do rodent models inform clinical practice? Hum. Reprod. Update 2018, 24, 416–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kretser, D.M.; Huidobro, C.; Southwick, G.J.; Temple-Smith, P.D. The role of the epididymis in human infertility. J. Reprod. Fertil. Suppl. 1998, 53, 271–275. [Google Scholar] [PubMed]
- Guiton, R.; Henry-Berger, J.; Drevet, J.R. The immunobiology of the mammalian epididymis: The black box is now open! Basic Clin. Androl. 2013, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Pilatz, A.; Lochnit, G.; Karnati, S.; Paradowska-Dogan, A.; Lang, T.; Schultheiss, D.; Schuppe, H.C.; Hossain, H.; Baumgart-Vogt, E.; Weidner, W.; et al. Acute epididymitis induces alterations in sperm protein composition. Fertil. Steril. 2014, 101, 1609–1617.e5. [Google Scholar] [CrossRef]
- Schirinzi, A.; Cazzolla, A.P.; Mascolo, E.; Palmieri, G.; Pesce, F.; Gesualdo, L.; Santacroce, L.; Ballini, A.; Lovero, R.; Di Serio, F. Determination of the Upper Reference Limit of Human Epididymis Secretory Protein 4 (HE4) in Healthy Male Individuals and Correlation with Renal and Fertility Markers. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 912–918. [Google Scholar] [CrossRef]
- Stammler, A.; Hau, T.; Bhushan, S.; Meinhardt, A.; Jonigk, D.; Lippmann, T.; Pilatz, A.; Schneider-Hüther, I.; Middendorff, R. Epididymitis: Ascending infection restricted by segmental boundaries. Hum. Reprod. 2015, 30, 1557–1565. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.J.R.; Ribeiro, C.M.; Mirim, A.F.M.; Silva, A.A.S.; Romano, R.M.; Hallak, J.; Avellar, M.C.W. Lipopolysaccharide and lipotheicoic acid differentially modulate epididymal cytokine and chemokine profiles and sperm parameters in experimental acute epididymitis. Sci. Rep. 2018, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Mutoji, K.; Singh, A.; Nguyen, T.; Gildersleeve, H.; Kaucher, A.V.; Oatley, M.J.; Oatley, J.M.; Velte, E.K.; Geyer, C.B.; Cheng, K.; et al. TSPAN8 Expression Distinguishes Spermatogonial Stem Cells in the Prepubertal Mouse Testis. Biol. Reprod. 2016, 95, 117. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Bhushan, S.; Rajesh, A.; Suraj, S.K.; Lu, Y.; Meinhardt, A.; Yenugu, S. Uropathogenic Escherichia coli (UPEC) induced antimicrobial gene expression in the male reproductive tract of rat: Evaluation of the potential of Defensin 21 to limit infection. Andrology 2015, 3, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Bryan, E.R.; Kollipara, A.; Trim, L.K.; Armitage, C.W.; Carey, A.J.; Mihalas, B.; Redgrove, K.A.; McLaughlin, E.A.; Beagley, K.W. Hematogenous dissemination of Chlamydia muridarum from the urethra in macrophages causes testicular infection and sperm DNA damage. Biol. Reprod. 2019, 101, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Mackern-Oberti, J.P.; Motrich, R.D.; Breser, M.L.; Sánchez, L.R.; Cuffini, C.; Rivero, V.E. Chlamydia trachomatis infection of the male genital tract: An update. J. Reprod. Immunol. 2013, 100, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Emerson, C.; Dinsmore, W.W.; Quah, S.P. Are we missing mumps epididymo-orchitis? Int. J. STD AIDS 2007, 18, 341–342. [Google Scholar] [CrossRef]
- Giagulli, V.A.; Guastamacchia, E.; Magrone, T.; Jirillo, E.; Lisco, G.; De Pergola, G.; Triggiani, V. Worse progression of COVID-19 in men: Is testosterone a key factor? Andrology 2021, 9, 53–64. [Google Scholar] [CrossRef]
- Santacroce, L.; Charitos, I.A.; Carretta, D.M.; De Nitto, E.; Lovero, R. The human coronaviruses (HCoVs) and the molecular mechanisms of SARS-CoV-2 infection. J. Mol. Med. 2021, 99, 93–106. [Google Scholar] [CrossRef]
- Schirinzi, A.; Cazzolla, A.P.; Lovero, R.; Lo Muzio, L.; Testa, N.F.; Ciavarella, D.; Palmieri, G.; Pozzessere, P.; Procacci, V.; Di Serio, F.; et al. New Insights in Laboratory Testing for COVID-19 Patients: Looking for the Role and Predictive Value of Human epididymis secretory protein 4 (HE4) and the Innate Immunity of the Oral Cavity and Respiratory Tract. Microorganisms 2020, 8, 1718. [Google Scholar] [CrossRef]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chen, S.; Huang, B.; Zhong, J.M.; Su, H.; Chen, Y.J.; Cao, Q.; Ma, L.; He, J.; Li, X.F.; et al. Pathological Findings in the Testes of COVID-19 Patients: Clinical Implications. Eur. Urol. Focus 2020, 6, 1124–1129. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Diet and Male Fertility: The Impact of Nutrients and Antioxidants on Sperm Energetic Metabolism. Int. J. Mol. Sci. 2022, 23, 2542. [Google Scholar] [CrossRef]
- Suliga, E.; Głuszek, S. The relationship between diet, energy balance and fertility in men. Int. J. Vitam. Nutr. Res. 2020, 90, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Willett, W.C. The Mediterranean diet and health: A comprehensive overview. J. Intern. Med. 2021, 290, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Bachir, B.G.; Jarvi, K. Infectious, inflammatory, and immunologic conditions resulting in male infertility. Urol. Clin. N. Am. 2014, 41, 67–81. [Google Scholar] [CrossRef]
- Morielli, T.; O’Flaherty, C. Oxidative stress impairs function and increases redox protein modifications in human spermatozoa. Reproduction 2015, 149, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Ferramosca, A.; Conte, A.; Moscatelli, N.; Zara, V. A high-fat diet negatively affects rat sperm mitochondrial respiration. Andrology 2016, 4, 520–525. [Google Scholar] [CrossRef] [Green Version]
- Casas, R.; Estruch, R.; Sacanella, E. The Protective Effects of Extra Virgin Olive Oil on Immune-mediated Inflammatory Responses. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 23–35. [Google Scholar] [CrossRef]
- Magrone, T.; Spagnoletta, A.; Salvatore, R.; Magrone, M.; Dentamaro, F.; Russo, M.A.; Difonzo, G.; Summo, C.; Caponio, F.; Jirillo, E. Olive Leaf Extracts Act as Modulators of the Human Immune Response. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 85–93. [Google Scholar] [CrossRef]
- Müller, A.K.; Albrecht, F.; Rohrer, C.; Koeberle, A.; Werz, O.; Schlörmann, W.; Glei, M.; Lorkowski, S.; Wallert, M. Olive Oil Extracts and Oleic Acid Attenuate the LPS-Induced Inflammatory Response in Murine RAW264.7 Macrophages but Induce the Release of Prostaglandin E2. Nutrients. 2021, 13, 4437. [Google Scholar] [CrossRef] [PubMed]
- Santacroce, L.; Inchingolo, F.; Topi, S.; Del Prete, R.; Di Cosola, M.; Charitos, I.A.; Montagnani, M. Potential beneficial role of probiotics on the outcome of COVID-19 patients: An evolving perspective. Diabetes Metab. Syndr. 2021, 15, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Versalovic, J. Probiotic Lactobacillus reuteri biofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Valcarce, D.G.; Genovés, S.; Riesco, M.F.; Martorell, P.; Herráez, M.P.; Ramón, D.; Robles, V. Probiotic administration improves sperm quality in asthenozoospermic human donors. Benef. Microbes 2017, 8, 193–206. [Google Scholar] [CrossRef]
- Pacifici, L.; Santacroce, L.; Dipalma, G.; Haxhirexha, K.; Topi, S.; Cantore, S.; Altini, V.; Pacifici, A.; De Vito, D.; Pettini, F.; et al. Gender medicine: The impact of probiotics on male patients. Clin. Ter. 2021, 171, e8–e15. [Google Scholar] [CrossRef]
- Abbasi, B.; Abbasi, H.; Niroumand, H. Synbiotic (FamiLact) administration in idiopathic male infertility enhances sperm quality, DNA integrity, and chromatin status: A triple-blinded randomized clinical trial. Int. J. Reprod. Biomed. 2021, 19, 235–244. [Google Scholar] [CrossRef]
- Dardmeh, F.; Alipour, H.; Gazerani, P.; van der Horst, G.; Brandsborg, E.; Nielsen, H.I. Lactobacillus rhamnosus PB01 (DSM 14870) supplementation affects markers of sperm kinematic parameters in a diet-induced obesity mice model. PLoS ONE 2017, 12, e0185964. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Gong, L.Z.; Xu, J.X. Antioxidative activity and protective effect of probiotics against high-fat diet-induced sperm damage in rats. Animal 2013, 7, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Di Domenico, M.; Ballini, A.; Boccellino, M.; Scacco, S.; Lovero, R.; Charitos, I.A.; Santacroce, L. The Intestinal Microbiota May Be a Potential Theranostic Tool for Personalized Medicine. J. Pers. Med. 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.E.; Kishibe, M.M.; Keller, R.; Caetano, H.R.D.S.; Rufino, M.N.; Sanches, O.C.; Giometti, I.C.; Giuffrida, R.; Bremer-Neto, H. Prebiotics mannan-oligosaccharides accelerate sexual maturity in rats: A randomized preclinical study. Vet. World 2021, 14, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Parekh, N.; Panner Selvam, M.K.; Henkel, R.; Shah, R.; Homa, S.T.; Ramasamy, R.; Ko, E.; Tremellen, K.; Esteves, S.; et al. Male Oxidative Stress Infertility (MOSI): Proposed Terminology and Clinical Practice Guidelines for Management of Idiopathic Male Infertility. World J. Men’s Health 2019, 37, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.A.; El-Beshbishy, H.A.; Banjar, Z.M. Mitochondrial dysfunction induced impairment of spermatogenesis in LPS-treated rats: Modulatory role of lycopene. Eur. J. Pharmacol. 2012, 677, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Schisterman, E.F.; Sjaarda, L.A.; Clemons, T.; Carrell, D.T.; Perkins, N.J.; Johnstone, E.; Lamb, D.; Chaney, K.; Van Voorhis, B.J.; Ryan, G.; et al. Effect of Folic Acid and Zinc Supplementation in Men on Semen Quality and Live Birth among Couples Undergoing Infertility Treatment: A Randomized Clinical Trial. JAMA 2020, 323, 35–48. [Google Scholar] [CrossRef]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. The effect of antioxidants on male factor infertility: The Males, Antioxidants, and Infertility (MOXI) randomized clinical trial. Fertil. Steril. 2020, 113, 552–560.e3. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Russo, M.A.; Jirillo, E. Recent Advances on the Anti-Inflammatory and Antioxidant Properties of Red Grape Polyphenols: In Vitro and In Vivo Studies. Antioxidants 2019, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.A.; Abdeljawaad, K.A.A.; Abdelrahman, A.H.M.; Jaragh-Alhadad, L.A.; Oraby, H.F.; Elkaeed, E.B.; Mekhemer, G.A.H.; Gabr, G.A.; Shawky, A.M.; Sidhom, P.A.; et al. Exploring Natural Product Activity and Species Source Candidates for Hunting ABCB1 Transporter Inhibitors: An In Silico Drug Discovery Study. Molecules 2022, 27, 3104. [Google Scholar] [CrossRef]
- Juan, M.E.; González-Pons, E.; Munuera, T.; Ballester, J.; Rodríguez-Gil, J.E.; Planas, J.M. trans-Resveratrol, a natural antioxidant from grapes, increases sperm output in healthy rats. J. Nutr. 2005, 135, 757–760. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Brevini, T.A.L.; Gandolfi, F.; Boiti, C.; Zerani, M.; Maranesi, M. The Role of Resveratrol in Mammalian Reproduction. Molecules 2020, 25, 4554. [Google Scholar] [CrossRef]
- Magrone, T.; Jirillo, E. The New Era of Nutraceuticals: Beneficial Effects of Polyphenols in Various Experimental and Clinical Settings. Curr. Pharm. Des. 2018, 24, 5229–5231. [Google Scholar] [CrossRef] [PubMed]
- Ranawat, P.; Pathak, C.M.; Khanduja, K.L. A new perspective on the quercetin paradox in male reproductive dysfunction. Phytother. Res. 2013, 27, 802–810. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Products against Testicular Dysbiosis | |||
|---|---|---|---|
| PROBIOTICS | PREBIOTICS | POLYPHENOLS | ANTI-OXIDANTS |
| Mann-oligosaccharides acceleration of sexual maturity in rats | Controversial results with quercetin and resveratrol since they are endowed with both anti-oxidants and pro-oxidants activities |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santacroce, L.; Imbimbo, C.; Ballini, A.; Crocetto, F.; Scacco, S.; Cantore, S.; Di Zazzo, E.; Colella, M.; Jirillo, E. Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine. J. Pers. Med. 2022, 12, 1335. https://doi.org/10.3390/jpm12081335
Santacroce L, Imbimbo C, Ballini A, Crocetto F, Scacco S, Cantore S, Di Zazzo E, Colella M, Jirillo E. Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine. Journal of Personalized Medicine. 2022; 12(8):1335. https://doi.org/10.3390/jpm12081335
Chicago/Turabian StyleSantacroce, Luigi, Ciro Imbimbo, Andrea Ballini, Felice Crocetto, Salvatore Scacco, Stefania Cantore, Erika Di Zazzo, Marica Colella, and Emilio Jirillo. 2022. "Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine" Journal of Personalized Medicine 12, no. 8: 1335. https://doi.org/10.3390/jpm12081335
APA StyleSantacroce, L., Imbimbo, C., Ballini, A., Crocetto, F., Scacco, S., Cantore, S., Di Zazzo, E., Colella, M., & Jirillo, E. (2022). Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine. Journal of Personalized Medicine, 12(8), 1335. https://doi.org/10.3390/jpm12081335