Manipulation of Pattern of Cell Differentiation in a hetR Mutant of Anabaena sp. PCC 7120 by Overexpressing hetZ Alone or with hetP


The State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
*
Author to whom correspondence should be addressed.
Life 2018, 8(4), 60; https://doi.org/10.3390/life8040060
Submission received: 2 October 2018
/
Revised: 23 November 2018
/
Accepted: 27 November 2018
/
Published: 30 November 2018
(This article belongs to the Special Issue Developmental Biology in Cyanobacteria)
Abstract
:In the filamentous cyanobacterium, Anabaena sp. PCC 7120, single heterocysts differentiate at semi-regular intervals in response to nitrogen stepdown. HetR is a principal regulator of heterocyst differentiation, and hetP and hetZ are two genes that are regulated directly by HetR. In a hetR mutant generated from the IHB (Institute of Hydrobiology) substrain of PCC 7120, heterocyst formation can be restored by moderate expression of hetZ and hetP. The resulting heterocysts are located at terminal positions. We used a tandem promoter, PrbcLPpetE, to express hetZ and hetP strongly in the hetR mutant. Co-expression of hetZ and hetP enabled the hetR mutant to form multiple contiguous heterocysts at both terminal and intercalary positions. Expression of hetZ, alone resulted in terminally located heterocysts, whereas expression of hetP, alone produced enlarged cells in strings. In the absence of HetR, formation of heterocysts was insensitive to the peptide inhibitor, RGSGR.
1. Introduction
Cyanobacteria are the most ancient, oxygenic, photosynthetic organisms on earth [1,2]. Some filamentous cyanobacteria produce specialized cells called heterocysts to fix N2 under aerobic conditions [3,4]. Heterocyst pattern formation represents one of the earliest multicellular patterning processes in the history of life [5].
Anabaena sp. PCC 7120 (Anabaena 7120) is a model strain often used for molecular studies of heterocyst differentiation and patterning. In response to nitrogen stepdown, this strain transforms some CO2-fixing vegetative cells into single N2-fixing heterocysts at semi-regular intervals [3]. Spacing of heterocysts depends largely on an activator-inhibitor patterning system [6,7]. HetR has been proposed to be the master regulator (activator) of heterocyst differentiation [8], and diffusible RGSGR-containing peptides (inhibitor) have been shown to inactivate HetR in vitro [9,10,11] and to induce the degradation of HetR in cells [12]. In addition, gene patA [13], which encodes a CheY-like response regulator with a PATAN domain at N-terminus [14], is required for a semi-regular pattern. In a patA mutant, heterocysts form almost exclusively at the ends of filaments. Both activation of RGSGR-containing peptide genes in differentiating cells [15,16] and expression of patA in vegetative cells [10] are required for normal pattern formation and depend on direct or indirect regulation by HetR [17].
In the wild type of Anabaena 7120, HetR accumulates in differentiating cells [18,19] and directly activates the expression of hetP [20] and hetZ [9]. The latter two genes are required for heterocyst differentiation and mediate the regulation under the control of HetR [21]. In the IHB substrain of Anabaena 7120 (Anabaena 7120 IHB) [22], co-expression of hetZ and hetP from PntcA restored heterocyst formation at the ends of filaments in the hetR::C.CE2 mutant, whereas expression of hetZ or hetP did not [21]; use of PpetE produced similar results (Zhang and Xu, unpublished). In a different version of Anabaena 7120, expression of hetZ from PpetE enabled a hetR deletion mutant to form multiple contiguous heterocysts (Mch) at intercalary and terminal positions [23]. The two versions of Anabaena 7120 appeared to differ partly from each other in the functions of hetZ and hetP, especially in the extent to which the expression of hetZ can bypass the hetR mutation.
Because hetZ and hetP overlap functionally [21], further enhanced expression of hetZ may also be able to restore heterocyst formation in the hetR mutant. Before the use of PntcA, we had used the PrbcLPpetE tandem promoter to express hetZ and (or) hetP in the hetR mutant of the Anabaena 7120 IHB. By expressing hetZ or by co-expressing hetZ and hetP from the tandem promoter, we were able to generate two different patterns in the hetR background: single terminal heterocysts and Mch. In addition to hetR, hetZ and hetP, a caspase-hemoglobinase-fold protease gene, called hetF, is also required for heterocyst differentiation in Anabaena 7120 [24]. In a hetF::Tn5-1087b mutant, however, PrbcLPpetE-hetZ allowed heterocysts to be formed at both terminal and intercalary positions.
2. Materials and Methods
2.1. General
Anabaena 7120 and derivatives, listed in Table 1, were cultured in BG11 medium [4] in the light, 30 μE m−2 s−1, on a rotary shaker. Erythromycin (5 μg mL−1), neomycin (20 μg mL−1) or spectinomycin (10 μg mL−1) was added to the medium as appropriate. Anabaena filaments were washed 3 times with BG110 (without nitrate) before induction of heterocyst formation in the same medium. Nitrogenase activities were measured as previously described [21].
Alcian blue staining of a heterocyst polysaccharide layer was performed as described by Hebbar and Curtis (2000) [26]. Heterocyst frequencies (>300 cells counted per sample) and frequency distribution of intervals between heterocysts (>1000 cells counted per sample) were analyzed using three cultures in parallel. Data are means ± SD.
2.2. Plasmid Construction and Conjugation
Plasmid construction is briefly described here and detailed in Table S1. A PCR fragment containing PpetE was cloned in pTA2 (T-vector; Toyobo, Osaka, Japan), and a PCR fragment containing PrbcL was cloned in pMD18-T (T-vector; Takara, Shiga, Japan). PpetE was then excised and inserted downstream of PrbcL to generate PrbcLPpetE. PrbcLPpetE-hetZ and PrbcLPpetE-hetP were generated by overlap PCR [27] and cloned into pMD-18T. PpetE-hetP was generated by inserting the fragment containing hetP downstream of PpetE, excised and cloned downstream of PrbcLPpetE-hetZ to generate PrbcLPpetE-hetZ-PpetE-hetP. PrbcLPpetE-hetZ, PrbcLPpetE-hetP and PrbcLPpetE-hetZ-PpetE-hetP were then cloned into a shuttle vector derived from pRL25C [28], producing pHB4382, pHB4409 and pHB4551, respectively. Fragments cloned by PCR were all confirmed by sequencing. Plasmids were introduced into Anabaena 7120 and its derivatives by conjugation with the aid of a helper plasmid that carried methylase genes [29].
3. Results
3.1. Formation of Mch in a hetR Mutant that Overexpresses hetZ and hetP
We employed the tandem promoter PrbcLPpetE to express hetZ and hetP in the hetR background. PrbcL is strongly expressed in vegetative cells but not expressed in heterocysts [30], and PpetE is moderately expressed in both cell types [31]. Use of PpetE in addition to PrbcL was to ensure that the expression of hetZ and hetP was not switched off in developing heterocysts. Because PrbcL may enhance the expression of a reporter gene by 30–40 fold [32], the tandem promoter PrbcLPpetE could increase the expression to a higher level.
The hetR::C.CE2 mutant carrying PrbcLPpetE-hetZ produced heterocysts (including developing heterocysts, all but ca. 3% at the ends of filaments), and showed anoxic nitrogenase activity at 24 h after nitrogen stepdown and aerobic nitrogenase activity at 48 h (Table 2). The differentiated cells were stained by Alcian blue; they occasionally showed polar nodules, indicative of heterocysts. The frequency of differentiated cells was little changed in the second 24 h after nitrogen stepdown. The strain bearing PrbcLPpetE-hetP, however, showed few differentiating cells in the first 24 h, but some enlarged cells in the second 24 h (Figure 1). Because it showed anaerobic nitrogenase activity (Table 2), some of those enlarged cells evidently had heterocyst-like functions. Most of such cells formed in strings in a small fraction of filaments, contained particulate inclusions, were larger, and had thicker envelopes than heterocysts.
We then tested co-expression of hetZ and hetP from the tandem promoter in the hetR mutant (Figure 1). In BG11 medium (with nitrate), hetR::C.CE2 with PrbcLPpetE-hetZ-PpetE-hetP produced heterocysts at a frequency of 3.0%, whereas the wild type strain produced almost no heterocysts under the same conditions. After nitrogen stepdown, the heterocyst frequency increased to 11.5% at 24 h and 37.8% at 96 h. Co-expression of hetZ and hetP resulted in higher anaerobic and aerobic nitrogenase activities than expression of hetZ or hetP alone (Table 2). However, unlike hetR::C.CE2 with PntcA-hetZ-hetP in the previous report [21], this strain showed no diazotrophic growth.
The plasmid carrying PrbcLPpetE-hetZ-PpetE-hetP was also introduced into the wild type and the hetZ::C.K2 ΔhetP double mutant [21]. The resulting strains also produced heterocysts in BG11 and showed increases in heterocyst frequency after nitrogen stepdown (Table 2). In the hetZ hetP double mutant, heterocyst formation was not restored by PhetR-hetR on the pDU1-based plasmid [21] but by PhetZ-hetZ or PhetP-hetP on the plasmid (Figure 2). PrbcLPpetE-hetZ-PpetE-hetP on the plasmid enabled the double mutant to form Mch.
3.2. Distribution of Heterocysts along Filaments
RGSGR has been established as an important factor in heterocyst pattern formation [16]. HetR is the only identified target for RGSGR or RGSGR-containing peptides. Now that heterocysts can be formed in a hetR mutant that overexpresses hetZ and hetP, it becomes feasible to test whether heterocyst formation without HetR is still susceptible to inhibition by RGSGR. Usually, heterocyst formation in the wild-type Anabaena 7120 can be significantly inhibited by 1 μM RGSGR and completely inhibited by 10 μM RGSGR [15]. However, RGSGR up to 50 μM showed no inhibitory effect on heterocyst formation in a hetR mutant that expresses hetZ or co-expresses hetZ and hetP (Table 2). PrbcLPpetE-hetZ-PpetE-hetP also enabled the wild type and the hetZ hetP double mutant to form heterocysts in the presence of 50 μM RGSGR (Table 2).
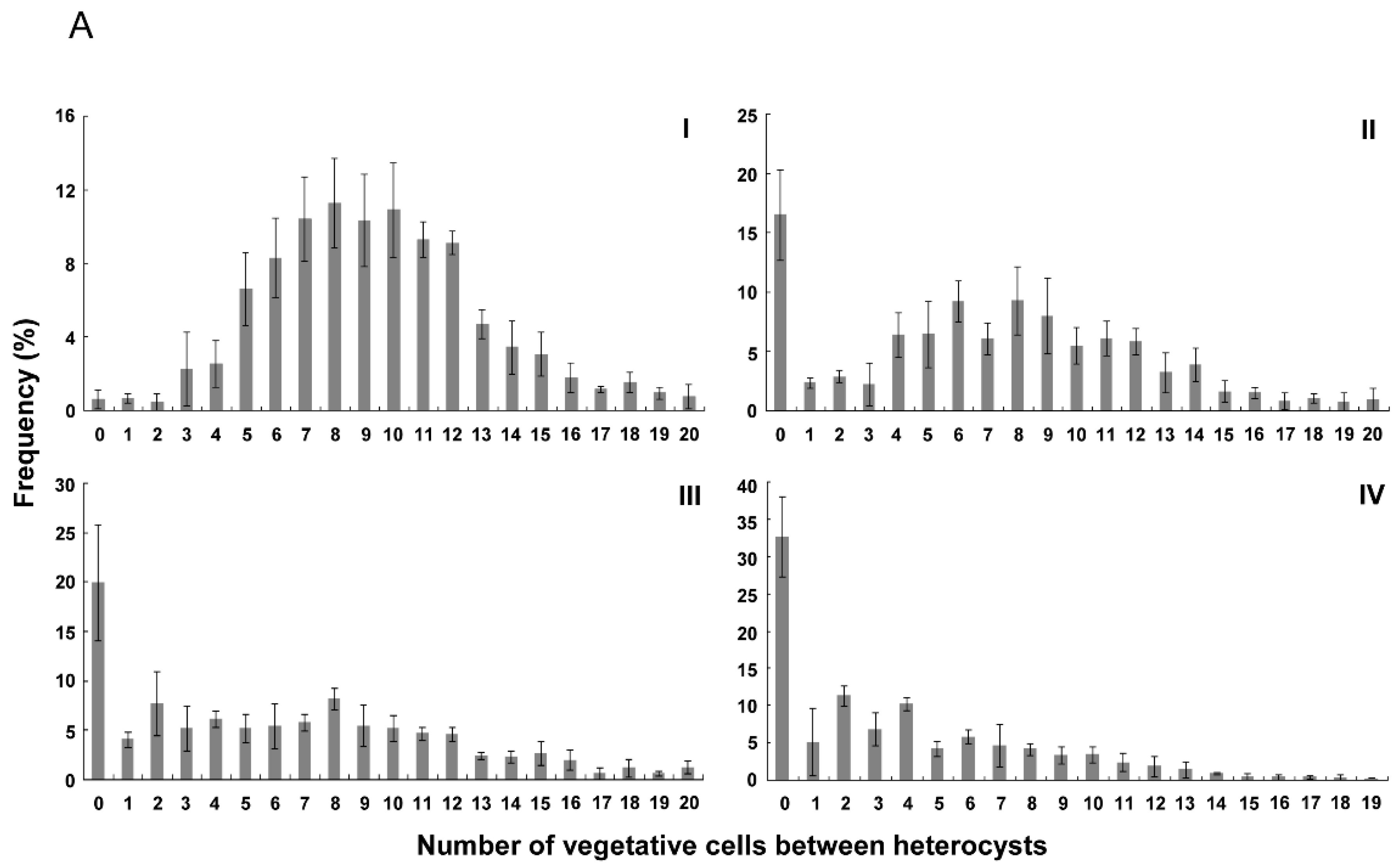
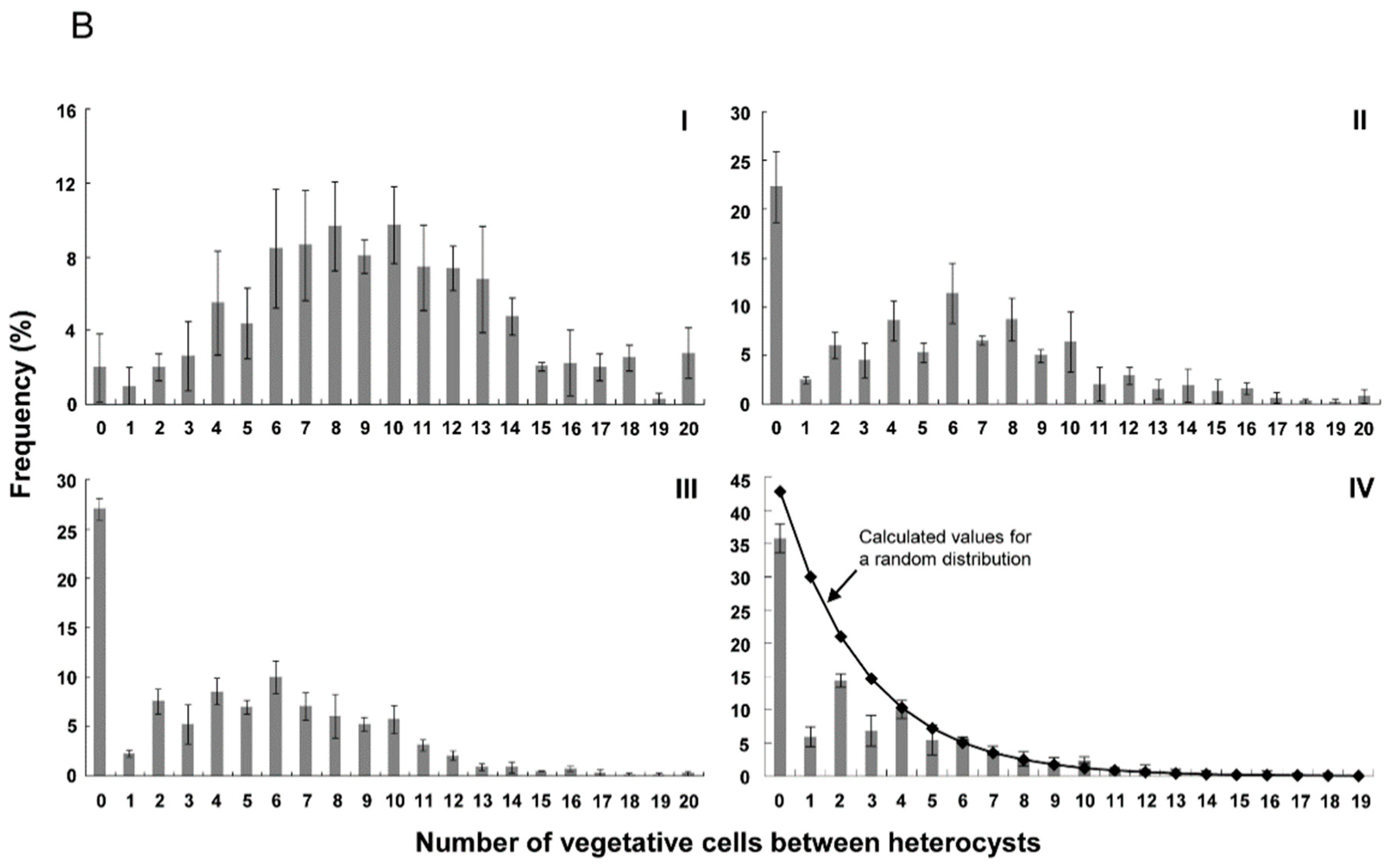
Figure 3 shows the distribution of number of vegetative cells between heterocysts in the wild type and strains with PrbcLPpetE-hetZ-PpetE-hetP. As reported by Khudyakov and Golden (2004) [33], there was a bias toward even numbers of intervals, probably due to cell division after differentiation of heterocysts. In hetR::C.CE2 with PrbcLPpetE-hetZ-PpetE-hetP, more than 30% of heterocysts were formed in strings after nitrogen stepdown. At 48 h, frequencies of 0 and even number intervals in this strain were close to calculated values of a random distribution (Figure 3B-IV).
In the hetZ hetP double mutant with PrbcLPpetE-hetZ-PpetE-hetP, more than 20% of heterocysts were in strings (≥2 cells). At 24 h, intervals in the range of 2 to 12 cells were at similar frequencies (Figure 3A); at 48 h, a peak at 6 cells emerged (Figure 3B). In the wild type with PrbcLPpetE-hetZ-PpetE-hetP, more than 15% of heterocysts were in strings, and the frequency distribution peaked at 6-8 cells at 24 h and at 6 cells at 48 h. As a control, in the wild type of Anabaena 7120, frequency distribution peaked at 8-10 cells, with <2% of heterocysts in strings.
It is noteworthy that in strains with PrbcLPpetE-hetZ or PrbcLPpetE-hetZ-PpetE-hetP, formation of heterocysts decreased gradually from serial subculture in BG11. Therefore, long-repeated subcultures should be avoided. In this study, heterocyst differentiation was observed with newly acquired exconjugants after two passages.
3.3. Heterocyst Formation in a hetF Background
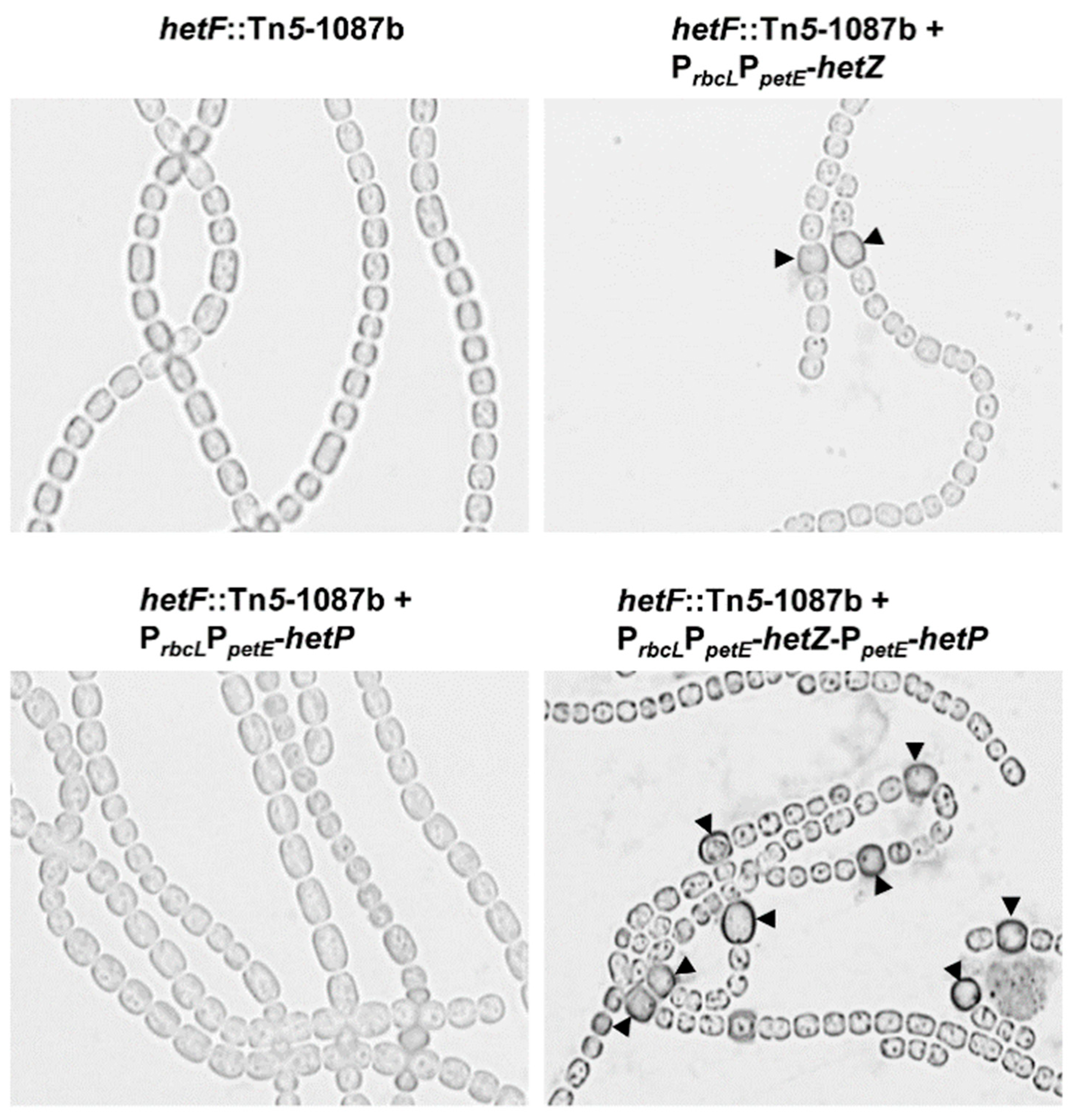
In a hetR-minus background, expression of hetZ from PrbcLPpetE or co-expression of hetZ and hetP from PntcA [21] or PpetE (Zhang and Xu, unpublished) restores heterocyst formation at the terminal positions of filaments. Such restoration might be due to the lack of expression of patA in vegetative cells [10] of a hetR mutant. To test this hypothesis, we introduced PrbcLPpetE-hetZ, PrbcLPpetE-hetP and PrbcLPpetE-hetZ-PpetE-hetP, respectively, into a hetF::Tn5-1087b mutant [9] to find out whether single heterocysts could be formed at intercalary positions in a hetF background, where hetR is active. Expression of hetZ or co-expression of hetZ and hetP restored heterocyst formation in a hetF mutant, whereas expression of hetP did not (Figure 4). In the strain expressing hetZ, about 47% of heterocysts were at intercalary positions (heterocyst frequency ca. 2.5% at 24 h). From 24 h to 48 h after nitrogen stepdown, the co-expressing strain showed an increase in heterocyst frequency from ca. 7.7% to ca. 21.3%, producing Mch, whereas the strains with hetZ or hetP—expressed alone—remained essentially unchanged.
4. Discussion
Restoration of heterocyst formation in a hetR-minus background has been shown in two laboratories by expression of hetZ and hetP [21] or by hetZ alone [23]. Even though both showed heterocyst formation without a functional hetR, the two reports differed in the phenotypes and genes involved. One group [23] proposed that the differences might be due to the promoters, PntcA and PpetE, used in the two studies. However, PpetE had been tested in the IHB version of Anabaena 7120 (Zhang and Xu, unpublished) and had found results similar to those produced with PntcA. Therefore, the differences are more likely due to divergence in genetic backgrounds of versions of Anabaena 7120 [22,23].
In this study, we used the PrbcLPpetE tandem promoter to express the genes in the hetR mutant of Anabaena 7120 IHB and found two different patterns of heterocyst distribution, namely, with single heterocysts only at terminal positions, or with Mch at both terminal and intercalary positions. Now that heterocyst formation without HetR has been shown with moderate expression of hetZ and hetP, the use of stronger expression may further intensify the phenotype. We admit that expression of hetZ and hetP from the tandem promoter had certain side effects on cells, because the phenotypes would gradually disappear if the strains continued to be cultured and subcultured in BG11. Therefore, the observations must be performed with cultures within a suitable range of generations.
In Anabaena 7120 IHB, moderate co-expression of hetZ and hetP [21] or strong expression of hetZ (from PrbcLPpetE) (Figure 1) in the hetR mutant led to heterocyst formation predominantly at terminal positions. Because the expression of patA in vegetative cells is dependent on HetR and required for heterocyst formation at the intercalary positions [10], we assumed that the patA-like phenotype in these strains were due to the lack of a functional HetR-patA system. When hetP was added to the expression from PrbcLPpetE, heterocyst formation was so much more enhanced that the lack of a HetR-patA system was bypassed, and Mch were produced at intercalary and terminal positions. Consistent with the assumption, in a hetF mutant with PrbcLPpetE-hetZ, where HetR-patA system probably remained active in vegetative cells, single heterocysts formed at both terminal and intercalary positions (Figure 4).
In parallel to the HetR-patA system that determines heterocyst formation at intercalary positions, the HetR-RGSGR system plays a key role in determination of heterocyst spacing. HetR is the only identified target for RGSGR-containing peptides in terms of heterocyst differentiation. Therefore, heterocyst formation without HetR may not be inhibited by RGSGR. Addition of RGSGR at a high concentration to different strains under N-deficient conditions confirmed this assumption (Table 2). Because of the lack of HetR as the sensor of RGSGR, the distribution of heterocysts in the hetR mutant with PrbcLPpetE-hetZ-PpetE-hetP (Figure 3B) resembled that of the PatS-resistant hetRR223W mutant described in a previous report [32]. However, in the wild type or the hetZ hetP double mutant that carried the same plasmid, the normal heterocyst pattern was partially maintained. To some extent, the distribution of heterocysts in the latter two strains were affected by the HetR-RGSGR system.
In summary, we have been able to generate different patterns of heterocyst distribution in hetR and hetF mutants by overexpressing hetZ, hetP or both. These results extended the previously reported findings on restoration of heterocyst formation in the hetR mutant [21] and provided some information that may help to understand heterocyst pattern formation, especially the phenotype of a patA mutant.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-1729/8/4/60/s1, Table S1: A list of plasmids and primers.
Author Contributions
H.Z. performed the experiments; X.X. designed the experiments and wrote the manuscript; X.X. and Z.H. analyzed the data.
Funding
This study was supported by the National Natural Science Foundation of China (Grant no. 31270132 and 31770044).
Acknowledgments
We thank Hong Gao and Xiantao Fang for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rasmussen, B.; Fletcher, I.R.; Brocks, J.J.; Kilburn, M.R. Reassessing the first appearance of eukaryotes and cyanobacteria. Nature 2008, 455, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W. The paleobiological record of photosynthesis. Photosynth. Res. 2011, 107, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Wolk, C.P.; Ernst, A.; Elhai, J. Heterocyst metabolism and development. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 769–823. [Google Scholar]
- Castenholz, R.W. Oxygenic photosynthetic bacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 2001; pp. 474–487. [Google Scholar]
- Tomitani, A.; Knoll, A.H.; Cavanaugh, C.M.; Ohno, T. The evolutionary diversification of cyanobacteria: Molecular-phylogenetic and paleontological perspectives. Proc. Natl. Acad. Sci. USA 2006, 103, 5442–5447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinhardt, H.; Gierer, A. Pattern formation by local self-activation and lateral inhibition. Bioessays 1991, 22, 753–760. [Google Scholar] [CrossRef]
- Kondo, S.; Miura, T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science 2006, 329, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Expression of the Anabaena hetR gene from a copper-regulated promoter leads to heterocyst differentiation under repressing conditions. Proc. Natl. Acad. Sci. USA 2001, 98, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Cai, Y.; Hou, S.; Xu, X. Identification of the HetR-recognition sequence upstream of hetZ in Anabaena sp. strain PCC 7120. J. Bacteriol. 2012, 194, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Zhou, F.; Peng, S.; Gao, H.; Xu, X. The HetR-binding site that activates expression of patA in vegetative cells is required for normal heterocyst patterning in Anabaena sp. PCC 7120. Sci. Bull. 2015, 60, 192–201. [Google Scholar] [CrossRef]
- Hu, H.X.; Jiang, Y.L.; Zhao, M.X.; Cai, K.; Liu, S.; Wen, B.; Lv, P.; Zhang, Y.; Peng, J.; Zhong, H.; et al. Structural insights 249 into HetR-PatS interaction involved in cyanobacterial pattern formation. Sci. Rep. 2015, 5, 16470. [Google Scholar] [CrossRef] [PubMed]
- Risser, D.D.; Callahan, S.M. Genetic and cytological evidence that heterocyst patterning is regulated by inhibitor gradients that promote activator decay. Proc. Natl. Acad. Sci. USA 2009, 106, 19884–19888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Scappino, L.; Haselkorn, R. The patA gene product, which contains a region similar to CheY of Escherichia coli, controls heterocyst pattern formation in the cyanobacterium Anabaena 7120. Proc. Natl. Acad. Sci. USA 1992, 89, 5655–5659. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Koonin, E.V.; Haselkorn, R.; Galperin, M.Y. Cyanobacterial response regulator PatA contains a conserved N-terminal domain (PATAN) with an alpha-helical insertion. Bioinformatics 2006, 22, 1297–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.S.; Golden, J.W. Heterocyst pattern formation controlled by a diffusible peptide. Science 1998, 282, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Khudyakov, I. Ancient association of cyanobacterial multicellularity with the regulator HetR and an RGSGR pentapeptide-containing protein (PatX). Mol. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Vioque, A.; Haas, F.; Hess, W.R.; Muro-Pastor, A.M. Dynamics of transcriptional start site selection during nitrogen stress-induced cell differentiation in Anabaena sp. PCC7120. Proc. Natl. Acad. Sci. USA 2011, 108, 20130–20135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Cao, Z.; Zhao, J. Characterization of HetR protein turnover in Anabaena sp. PCC 7120. Arch. Microbiol. 1998, 169, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Black, T.A.; Cai, Y.; Wolk, C.P. Spatial expression and autoregulation of hetR, a gene involved in the control of heterocyst development in Anabaena. Mol. Microbiol. 1993, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Higa, K.C.; Callahan, S.M. Ectopic expression of hetP can partially bypass the need for hetR in heterocyst differentiation by Anabaena sp. strain PCC 7120. Mol. Microbiol. 2010, 77, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, S.; Wang, Y.; Xu, X. Functional overlap of hetP and hetZ in regulation of heterocyst differentiation in Anabaena sp. strain PCC 7120. J. Bacteriol. 2018, 200, e00707-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, Y.; Li, C.; Gao, H.; Zhang, C.-C.; Xu, X. Three substrains of the cyanobacterium Anabaena sp. PCC 7120 display divergence in genomic sequences and hetC function. J. Bacteriol. 2018, 200, e00076-18. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Rivers, O.S.; Tom, S.K.; Oshiro, R.T.; Ushijima, B.; Swenson, V.A.; Philmus, B.; Gaylor, M.O.; Cozy, L.M. The hetZ gene indirectly regulates heterocyst development at the level of pattern formation in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2018, 109, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Risser, D.D.; Callahan, S.M. HetF and PatA control levels of HetR in Anabaena sp. strain PCC 7120. J. Bacteriol. 2008, 190, 7645–7654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, Y.; Khudyakov, I.; Fan, Q.; Gao, H.; Ning, D.; Wolk, C.P.; Xu, X. A gene cluster that regulates both heterocyst differentiation and pattern formation in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2007, 66, 1429–1443. [Google Scholar] [CrossRef] [PubMed]
- Hebbar, P.B.; Curtis, S.E. Characterization of devH, a gene encoding a putative DNA binding protein required for heterocyst function in Anabaena sp. strain PCC 7120. J. Bacteriol. 2000, 182, 3572–3581. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.; Hunt, H.D.; Ho, S.N.; Pullen, J.K.; Pease, L.R. Engineering hybrid genes without the use of restriction enzymes: Gene splicing by overlap extension. Gene 1989, 77, 61–68. [Google Scholar] [CrossRef]
- Wolk, C.P.; Cai, Y.; Cardemil, L.; Flores, E.; Hohn, B.; Murry, M.; Schmetterer, G.; Schrautemeier, B.; Wilson, R. Isolation and complementation of mutants of Anabaena sp. strain PCC 7120 unable to grow aerobically on dinitrogen. J. Bacteriol. 1988, 170, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 1998–2005. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Wolk, C.P. Developmental regulation and spatial pattern of expression of the structural genes for nitrogenase in the cyanobacterium Anabaena. EMBO J. 1990, 9, 3379–3388. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Oshiro, R.T.; Cozy, L.M.; Callahan, S.M. Transcriptional dynamics of developmental genes assessed with an FMN-dependent fluorophore in mature heterocysts of Anabaena sp. strain PCC 7120. Microbiology 2014, 160, 1874–1881. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J. Strong and regulated promoters in the cyanobacterium Anabaena PCC 7120. FEMS Microbiol. Lett. 1993, 114, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Khudyakov, I.Y.; Golden, J.W. Different functions of HetR, a master regulator of heterocyst differentiation in Anabaena sp. PCC 7120, can be separated by mutation. Proc. Natl. Acad. Sci. USA 2004, 101, 16040–16045. [Google Scholar] [CrossRef] [PubMed]
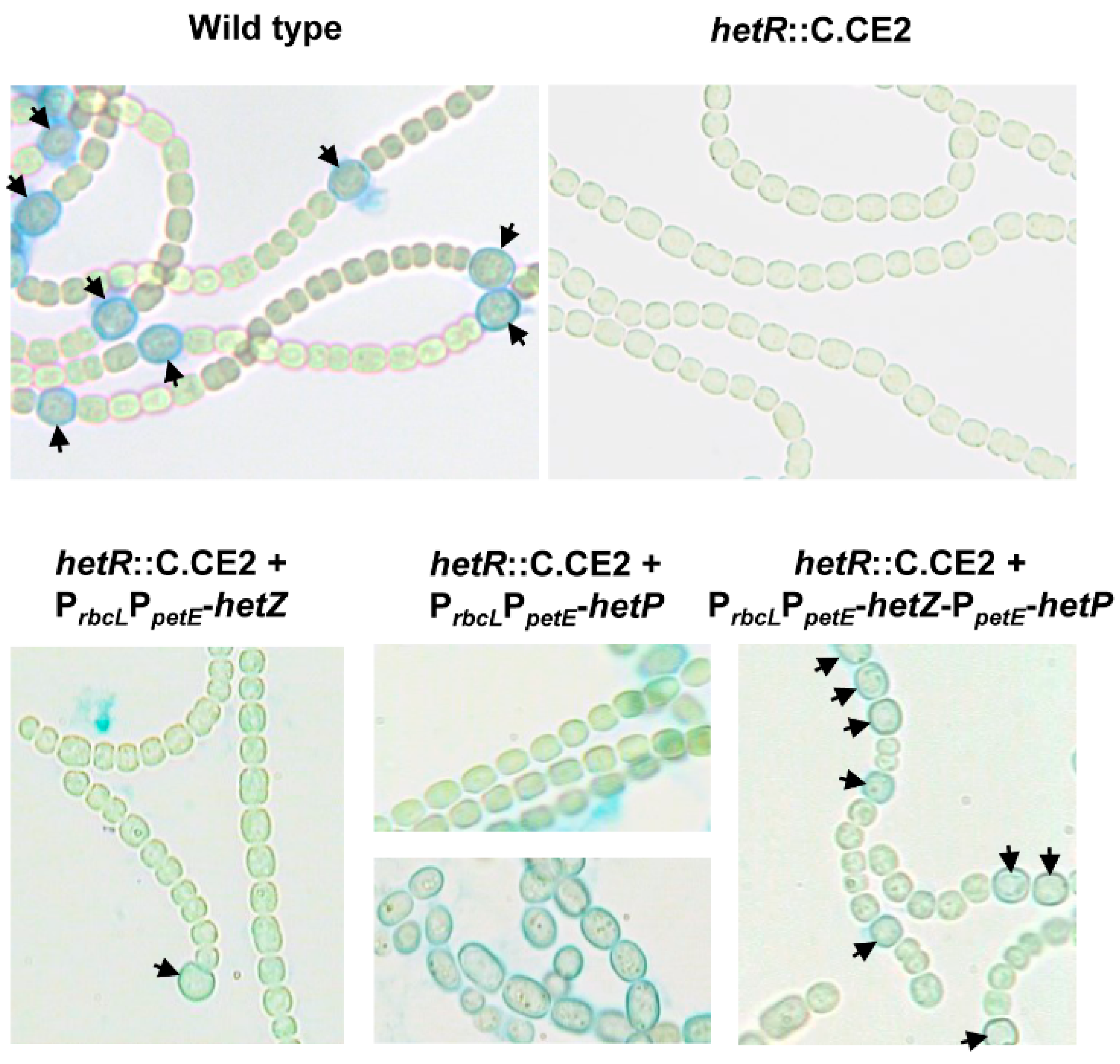
Figure 1.
Heterocyst formation in hetR::C.CE2 expressing hetZ, hetP or both from PrbcLPpetE. Filaments were stained by Alcian blue at 24 h after nitrogen stepdown. Most filaments of hetR::C.CE2 with PrbcLPpetE-hetP showed no cell differentiation (upper panel), but a small number of filaments produced enlarged cells in strings (lower panel). Arrowheads point to mature or developing heterocysts.
Figure 1.
Heterocyst formation in hetR::C.CE2 expressing hetZ, hetP or both from PrbcLPpetE. Filaments were stained by Alcian blue at 24 h after nitrogen stepdown. Most filaments of hetR::C.CE2 with PrbcLPpetE-hetP showed no cell differentiation (upper panel), but a small number of filaments produced enlarged cells in strings (lower panel). Arrowheads point to mature or developing heterocysts.

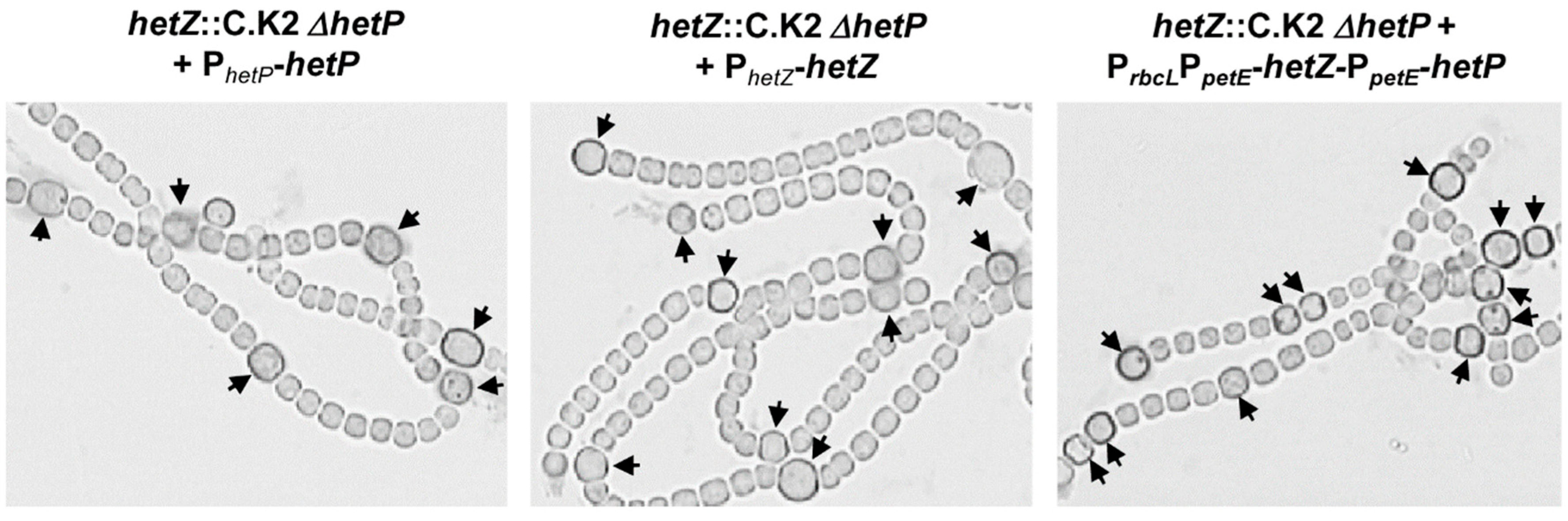
Figure 2.
Restoration of heterocyst formation in the hetZ::C.K2 ΔhetP double mutant by expressing hetZ, hetP or both. Filaments were stained by Alcian blue at 24 h after N-stepdown. Arrowheads point to mature or developing heterocysts.
Figure 2.
Restoration of heterocyst formation in the hetZ::C.K2 ΔhetP double mutant by expressing hetZ, hetP or both. Filaments were stained by Alcian blue at 24 h after N-stepdown. Arrowheads point to mature or developing heterocysts.

Figure 3.
Distribution of numbers of vegetative cells between heterocysts at 24 h (A) and 48 h (B) after nitrogen stepdown. I, the wild type of Anabaena 7120 (WT); II, WT + PrbcLPpetE-hetZ-PpetE-hetP; III, hetZ::C.K2ΔhetP + PrbcLPpetE-hetZ-PpetE-hetP; IV, hetR::C.CE2 + PrbcLPpetE-hetZ-PpetE-hetP. Intervals longer than 19 cells are shown as 20. The calculated values for a random distribution were generated by Microsoft Excel. At 24 h after nitrogen stepdown, hetR::C.CE2 carrying PrbcLPpetE-hetZ-PpetE-hetP produced heterocysts in 69.9 ± 2.4% of filaments, and of these filaments, the heterocyst frequency was 20.5 ± 2.9%. At 48 h, it produced heterocysts in 74.7 ± 4.0% of filaments, the heterocyst frequency of these filaments was 24.0 ± 0.8%. Please note that in hetR::C.CE2 carrying PrbcLPpetE-hetZ-PpetE-hetP, filaments with heterocysts were usually shorter than those without differentiation and that heterocyst frequencies in Table 2 were calculated based on filaments with or without heterocysts.
Figure 3.
Distribution of numbers of vegetative cells between heterocysts at 24 h (A) and 48 h (B) after nitrogen stepdown. I, the wild type of Anabaena 7120 (WT); II, WT + PrbcLPpetE-hetZ-PpetE-hetP; III, hetZ::C.K2ΔhetP + PrbcLPpetE-hetZ-PpetE-hetP; IV, hetR::C.CE2 + PrbcLPpetE-hetZ-PpetE-hetP. Intervals longer than 19 cells are shown as 20. The calculated values for a random distribution were generated by Microsoft Excel. At 24 h after nitrogen stepdown, hetR::C.CE2 carrying PrbcLPpetE-hetZ-PpetE-hetP produced heterocysts in 69.9 ± 2.4% of filaments, and of these filaments, the heterocyst frequency was 20.5 ± 2.9%. At 48 h, it produced heterocysts in 74.7 ± 4.0% of filaments, the heterocyst frequency of these filaments was 24.0 ± 0.8%. Please note that in hetR::C.CE2 carrying PrbcLPpetE-hetZ-PpetE-hetP, filaments with heterocysts were usually shorter than those without differentiation and that heterocyst frequencies in Table 2 were calculated based on filaments with or without heterocysts.


Figure 4.
Heterocyst formation in the hetF::Tn5-1087b mutant expressing hetZ, hetP or both from PrbcLPpetE. Filaments were stained by Alcian blue at 24 h after N-stepdown. Arrowheads point to mature or developing heterocysts.
Figure 4.
Heterocyst formation in the hetF::Tn5-1087b mutant expressing hetZ, hetP or both from PrbcLPpetE. Filaments were stained by Alcian blue at 24 h after N-stepdown. Arrowheads point to mature or developing heterocysts.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Anabaena strains used in this work.
| Strains | Derivation/Relevant Characteristics a | Reference or Source |
|---|---|---|
| Anabaena 7120 | Wild Type (WT) | FACHB b |
| hetF::Tn5-1087b | Emr, Tn5-1087b inserted within hetF 310 bp from its 3′ terminus | [9] |
| hetR::C.CE2 | CmrEmr, C.CE2 (a chloramphenicol-resistance and erythromycin-resistance cassette) inserted into the ClaI site of hetR | [9] |
| hetR::C.CE2 (pHB4382) | CmrEmrNmr, pHB4382 bearing PrbcLPpetE-hetZ introduced into the hetR::C.CE2 mutant | This study |
| hetR::C.CE2 (pHB4409) | CmrEmrNmrSmrSpr, pHB4409 bearing PrbcLPpetE-hetP introduced into the hetR::C.CE2 mutant | This study |
| hetR::C.CE2 (pHB4551) | CmrEmrNmrSmrSpr, pHB4551 bearing PrbcLPpetE-hetZ-PpetE-hetP introduced into the hetR::C.CE2 mutant | This study |
| hetZ::C.K2 ΔhetP | CmrEmrNmr, hetZ hetP double mutant | [21] |
| hetZ::C.K2 ΔhetP (pHB1462) | CmrEmrNmrSmrSpr, pHB1462 bearing PhetZ-hetZ [25] introduced into the hetZ hetP double mutant | This study |
| hetZ::C.K2 ΔhetP (pHB4550) | CmrEmrNmrSmrSpr, pHB4550 bearing PhetP-hetP introduced into the hetZ hetP double mutant | This study |
| hetZ::C.K2 ΔhetP (pHB4551) | CmrEmrNmrSmrSpr, pHB4551 bearing PrbcLPpetE-hetZ-PpetE-hetP introduced into the hetZ hetP double mutant | This study |
| WT (pHB4551) | NmrSmrSpr, pHB4551 bearing PrbcLPpetE-hetZ-PpetE-hetP introduced into Anabaena 7120 | This study |
a Cm, chloramphenicol; Em, erythromycin; Nm, neomycin; Sm, streptomycin; Sp, spectinomycin; stated otherwise, the PrbcL, PhetZ, and PhetP templates for PCR reactions were Anabaena 7120 genomic DNA; b FACHB, Freshwater Algal Culture Collection of the Institute of Hydrobiology, Chinese Academy of Sciences.
Table 2.
Nitrogenase activities and heterocyst frequencies of Anabaena strains after nitrogen stepdown.
Table 2.
Nitrogenase activities and heterocyst frequencies of Anabaena strains after nitrogen stepdown.
| Strains | Hours | Nitrogenase Activity (μmol C2H4 mg Chla−1 h−1) a | Heterocyst Frequency (%) b | Diazotrophic Growth | ||
|---|---|---|---|---|---|---|
| Anoxic | Aerobic | −RGSGR | +RGSGR c | |||
| WT d | 24 | 3.65 ± 0.62 | 2.83 ± 0.11 | 9.4 ± 0.4 | 0 | Good |
| 48 | 6.74 ± 0.63 | 3.50 ± 0.57 | 10.6 ± 0.7 | Not tested | ||
| WT + PrbcLPpetE-hetZ-PpetE-hetP e1 | 24 | Not measured | Not measured | 11.9 ± 1.2 | 11.4 ± 1.3 | Moderate |
| 48 | Not measured | Not measured | 15.8 ± 2.8 | Not tested | ||
| hetR::C.CE2 | 24 | 0 | 0 | 0 | Not tested | No |
| 48 | 0 | 0 | 0 | Not tested | ||
| hetR::C.CE2 + PrbcLPpetE-hetZ | 24 | 1.04 ± 0.14 | 0 | 2.9 ± 0.9 | 3.6 ± 0.4 | No |
| 48 | 1.20 ± 0.46 | 0.81 ± 0.13 | 3.5 ± 0.9 | Not tested | ||
| hetR::C.CE2 + PrbcLPpetE-hetP | 24 | 0.17 ± 0.10 | 0 | Not counted f | Not tested | No |
| 48 | 1.57 ± 0.26 | 0 | Not counted | Not tested | ||
| hetR::C.CE2 + PrbcLPpetE-hetZ-PpetE-hetP e2 | 24 | 4.00 ± 1.86 | 0.90 ± 0.07 | 11.5 ± 2.0 | 12.9 ± 0.9 | No |
| 48 | 4.12 ± 0.27 | 1.52 ± 0.78 | 17.5 ± 2.0 | Not tested | ||
| hetZ::C.K2 ΔhetP | 24 | 0 | 0 | 0 | Not tested | No |
| 48 | 0 | 0 | 0 | Not tested | ||
| hetZ::C.K2 ΔhetP + PrbcLPpetE-hetZ-PpetE-hetP e3 | 24 | Not measured | Not measured | 14.3 ± 1.2 | 15.7 ± 0.7 | Moderate |
| 48 | Not measured | Not measured | 17.6 ± 0.4 | Not tested | ||
a Nitrogenase activity was evaluated based on acetylene reduction over 6 h. b Mature and developing heterocysts were both included. c RGSGR was added to BG110 at a final concentration of 50 μM. d Nitrogenase activity and heterocyst frequency of the wild type varied between different batches of experiments under the described conditions. For example, at 24 h, the nitrogenase activity under aerobic conditions may change in a range from 2.8 to 9.1 μmole C2H4 mg Chla−1 h−1, and heterocyst frequency may change from 8.7 to 10.8%. Data presented are from one batch of experiments. e1~e3 These strains produced heterocysts in BG11 (with nitrate) at 7.4 ± 0.9%, 3.0 ± 0.5% and 6.1 ± 1.0% respectively, while all other strains produced heterocysts at 0~0.4% under the same conditions. f It was difficult to distinguish N2-fixing cells from those enlarged cells in this strain.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, H.; Xu, X. Manipulation of Pattern of Cell Differentiation in a hetR Mutant of Anabaena sp. PCC 7120 by Overexpressing hetZ Alone or with hetP. Life 2018, 8, 60. https://doi.org/10.3390/life8040060
AMA Style
Zhang H, Xu X. Manipulation of Pattern of Cell Differentiation in a hetR Mutant of Anabaena sp. PCC 7120 by Overexpressing hetZ Alone or with hetP. Life. 2018; 8(4):60. https://doi.org/10.3390/life8040060
Chicago/Turabian StyleZhang, He, and Xudong Xu. 2018. "Manipulation of Pattern of Cell Differentiation in a hetR Mutant of Anabaena sp. PCC 7120 by Overexpressing hetZ Alone or with hetP" Life 8, no. 4: 60. https://doi.org/10.3390/life8040060
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.