
Evolutionary Steps in the Emergence of Life Deduced from the Bottom-Up Approach and GADV Hypothesis (Top-Down Approach)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction


2. General Approach for the Study of the Origin of Life (Bottom-Up Approach)
2.1. Chemical Analysis of Extracts from Old Rocks and Meteorites
- Twelve amino acids were found from about 3.1 billion-year-old pre-Cambrian Fig Tree Chart from the mining area of eastern Transvaal, South Africa [16]. Extracts from 3.8 billion-year-old Isua rock formed on the primitive Earth were also chemically analyzed, and both simple amino acids and hydrocarbons were detected from the extracts [17]. However, Nagy et al. have concluded, based on the extent of amino acid racemization, that the amino acids may be modern to a few tens of thousands of years old [17]. Van Zuilen et al. also asserted that previously presented evidence for ancient traces of life in the highly metamorphosed Early Archaean rock, or the metasomatic rock records, should be reassessed, which were earlier thought to provide the basis for inferences about early life [18].
- Several amino acids were detected in extracts of meteorites from space, such as Marchison meteorite, which may be similar to rocks on the primitive Earth [19]. Therefore, nowadays, amino acids identified in the Murchison chondroritic meteorite are thought to have been delivered to the early Earth by meteorites, asteroids, comets, and interplanetary dust particles, which may trigger the appearance of life by assisting in the synthesis of proteins via prebiotic polymerization reactions [20,21,22].
2.2. Physical Evolution Experiments
- About 60 years ago, Miller published results showing that amino acids and nucleobases were synthesized by repeated electrical discharging into a reducing gas mixture containing CH4, NH3, H2, and H2O, which imitates lightning in the primitive atmosphere [4,23,24,25]. Many geologists today consider that the early atmosphere was rather weakly reducing or even neutral, composed of mainly CO2 and N2, based on later studies on the primitive atmosphere [26]. However, it has been confirmed that significant amounts of amino acids can still be synthesized even with weakly reducing or neutral primitive atmospheric gas [27,28].
- It is supposed that deep-sea hydrothermal environments were important sites for the synthesis of bioorganic molecules, leading to the emergence of life [29]. Simple amino acids were synthesized with experimental equipment mimicking hydrothermal vents in a deep sea on the primitive Earth [30]. It has been also demonstrated that oligopeptides were synthesized from glycine in a flow reactor simulating a submarine hydrothermal system [31,32,33].
- Oró et al. suggested that cometary collisions with the primitive Earth provided the planet with both free energy and volatiles as important sources for creation of transient, gaseous environments, in which prebiotic synthesis may have taken place [21]. Experiments reproducing meteorite impacts or heavy bombardments to the primitive Earth were also carried out. After the impact, numerous organic molecules, including fatty acids, amines, nucleobases, and amino acids were detected. So, it is considered that organic molecules on the early Earth may have arisen from such impact syntheses [34,35].
2.3. Planetary Exploration and Astronomical Observation
2.4. Chemical Evolution Experiments
2.5. Biochemical Evolution Experiments
2.5.1. Catalytic Activities of [GADV]-Peptides Produced by Repeated Heat-Drying Processes
2.5.2. Catalytic Activities of [GADV]-Random Octapeptides
2.6. Limitations of Bottom-Up Approaches
3. A Top-down Approach for Understanding the Origin of Life ([GADV]-Protein World Hypothesis)
3.1. Origin of Entirely New Genes
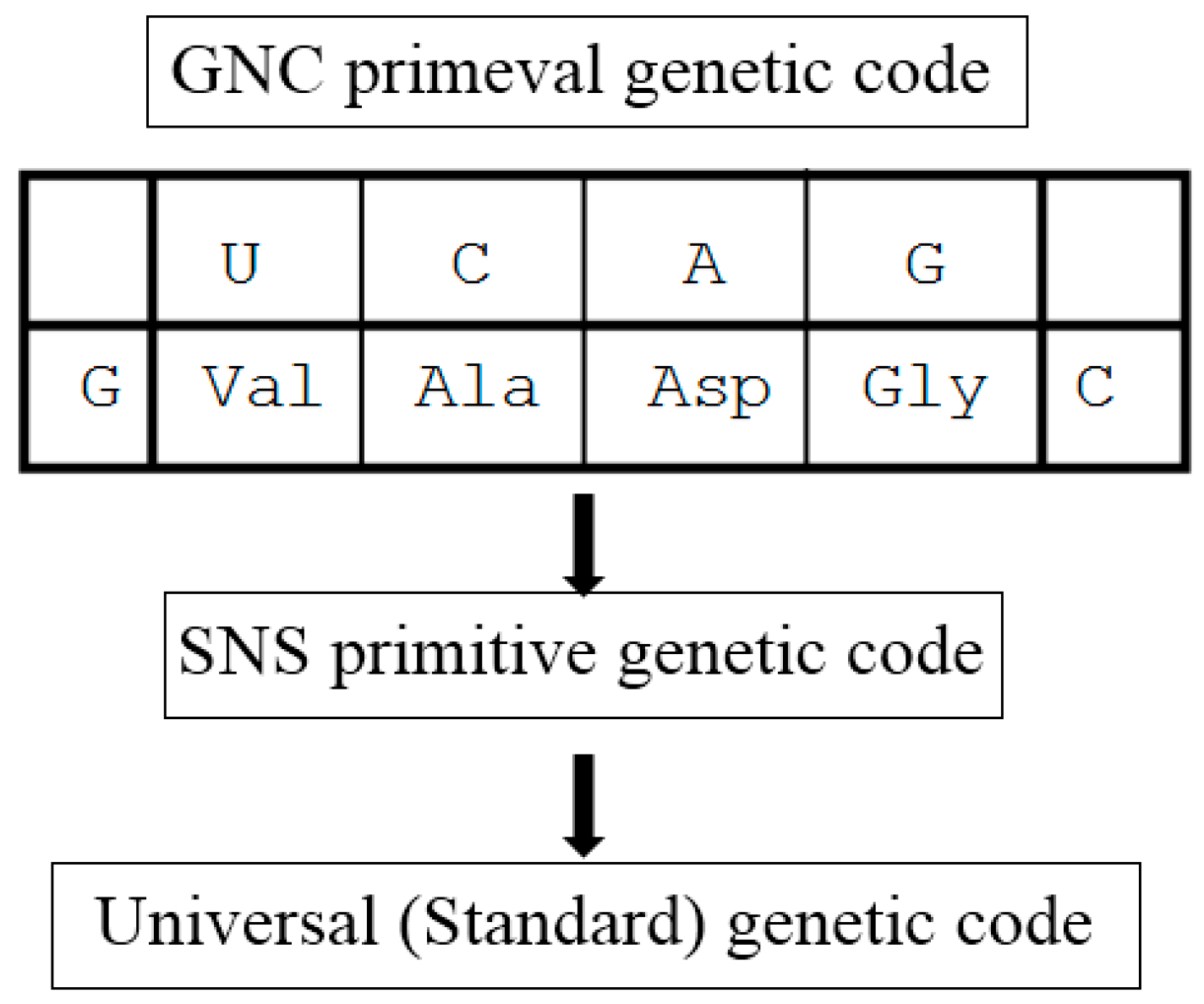
3.2. Origin of the Genetic Code
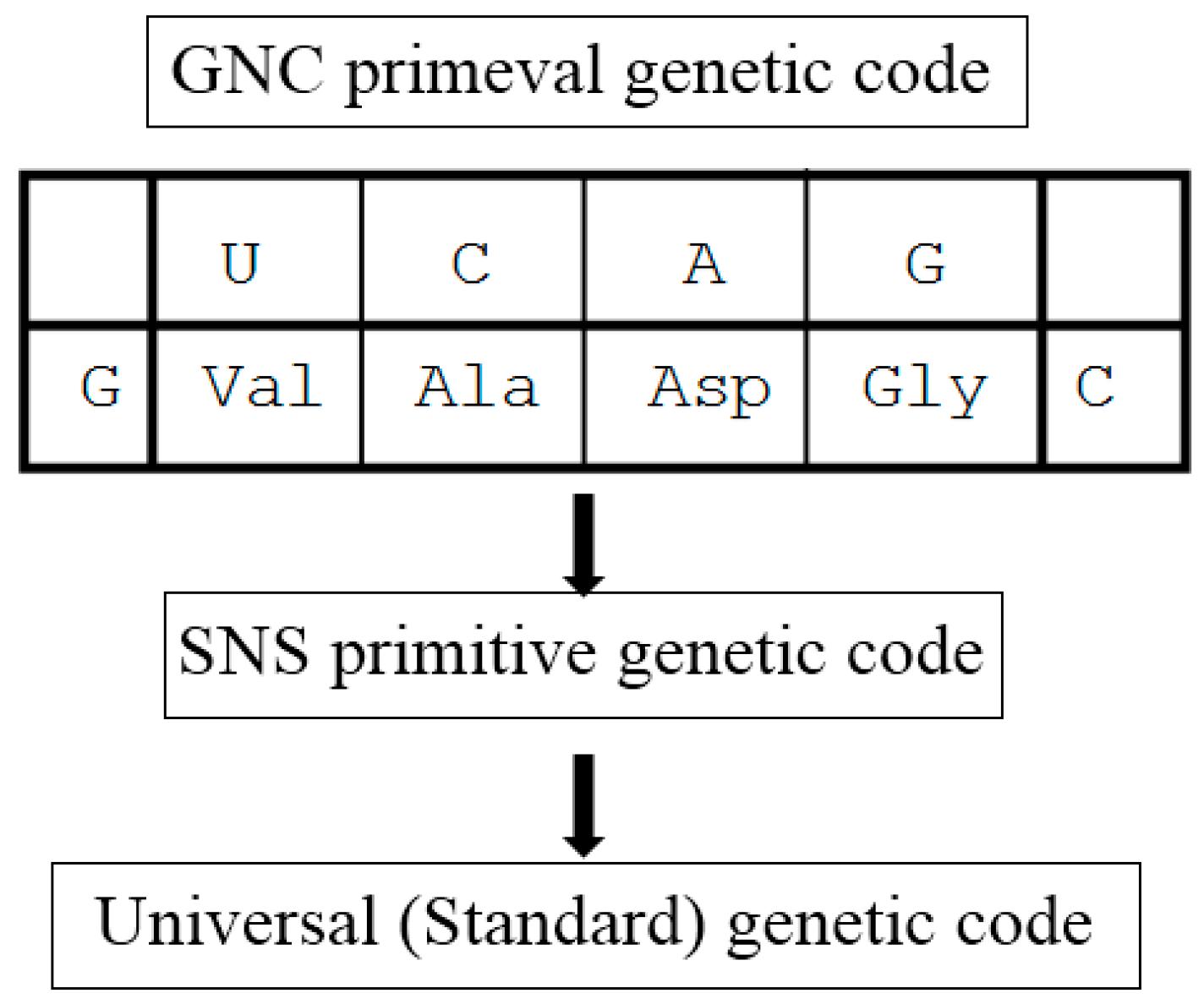
3.3. [GADV]-Protein World Hypothesis: GADV Hypothesis


3.4. Strengths of the GADV Hypothesis
3.5. Limitations of the GADV Hypothesis
4. The Necessity of Combining the Bottom-Up and Top-Down Approaches
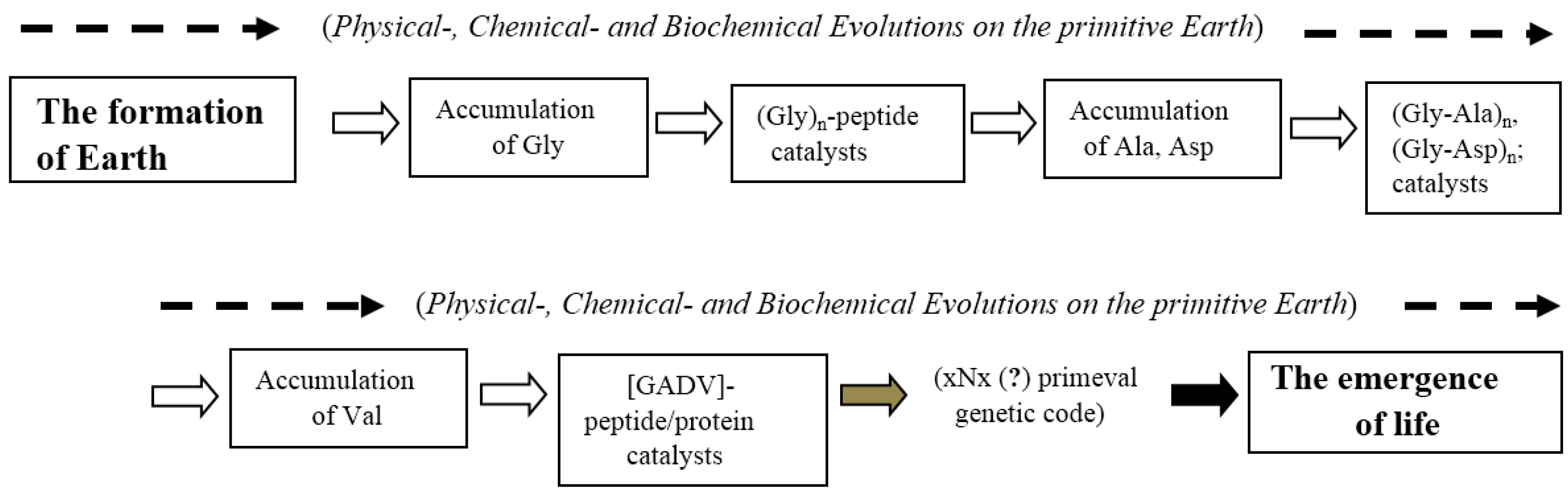
4.1. The Most Significant Evolutionary Steps according to Bottom-Up Approaches
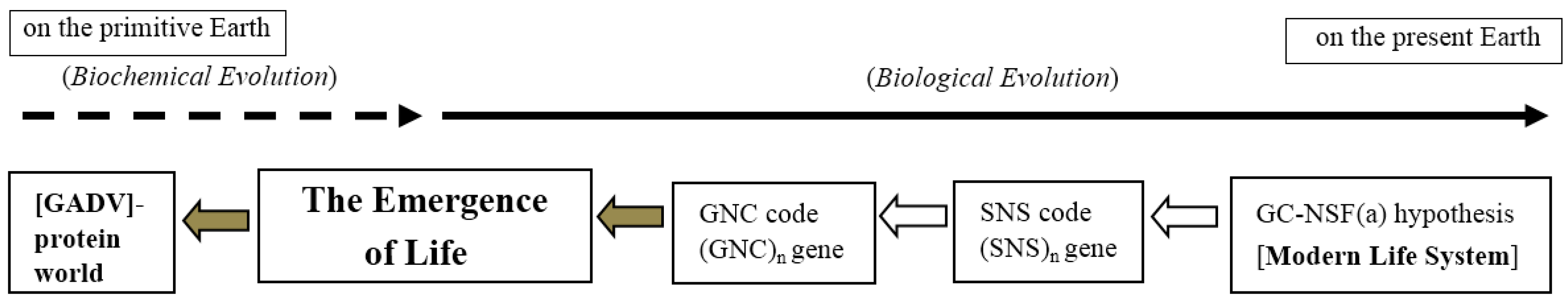
4.2. The Most Ancient Event Deduced Using the GADV Hypothesis (Top-Down Approach)
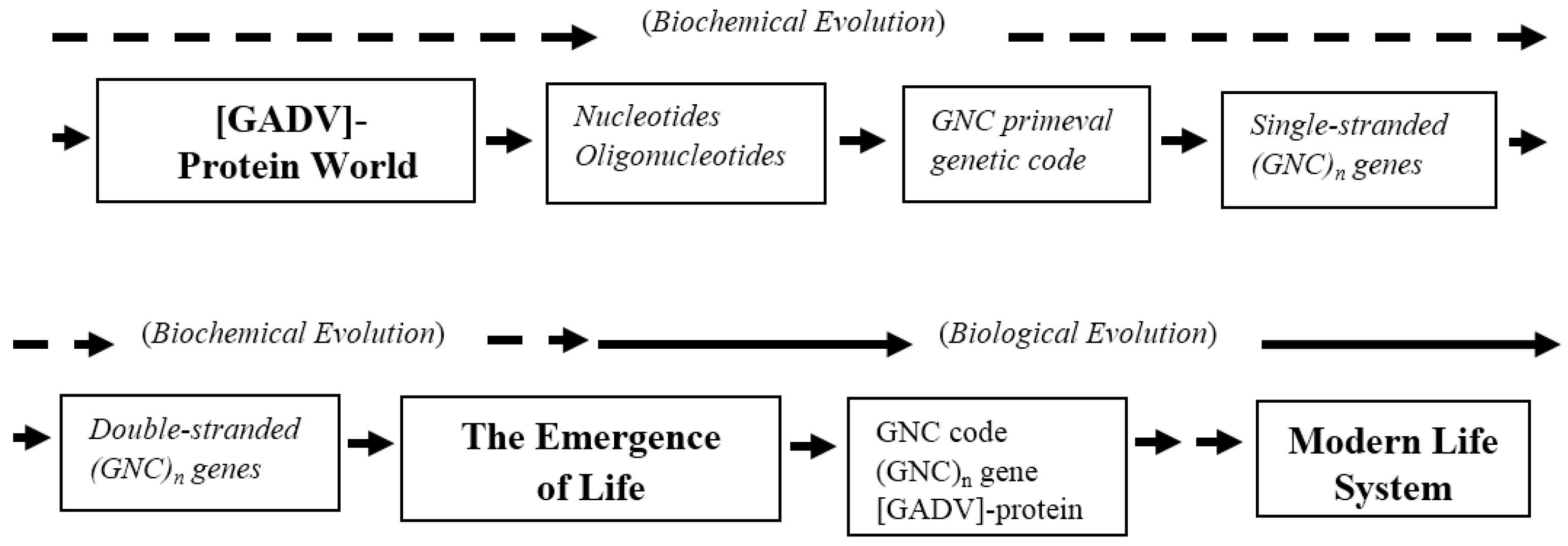
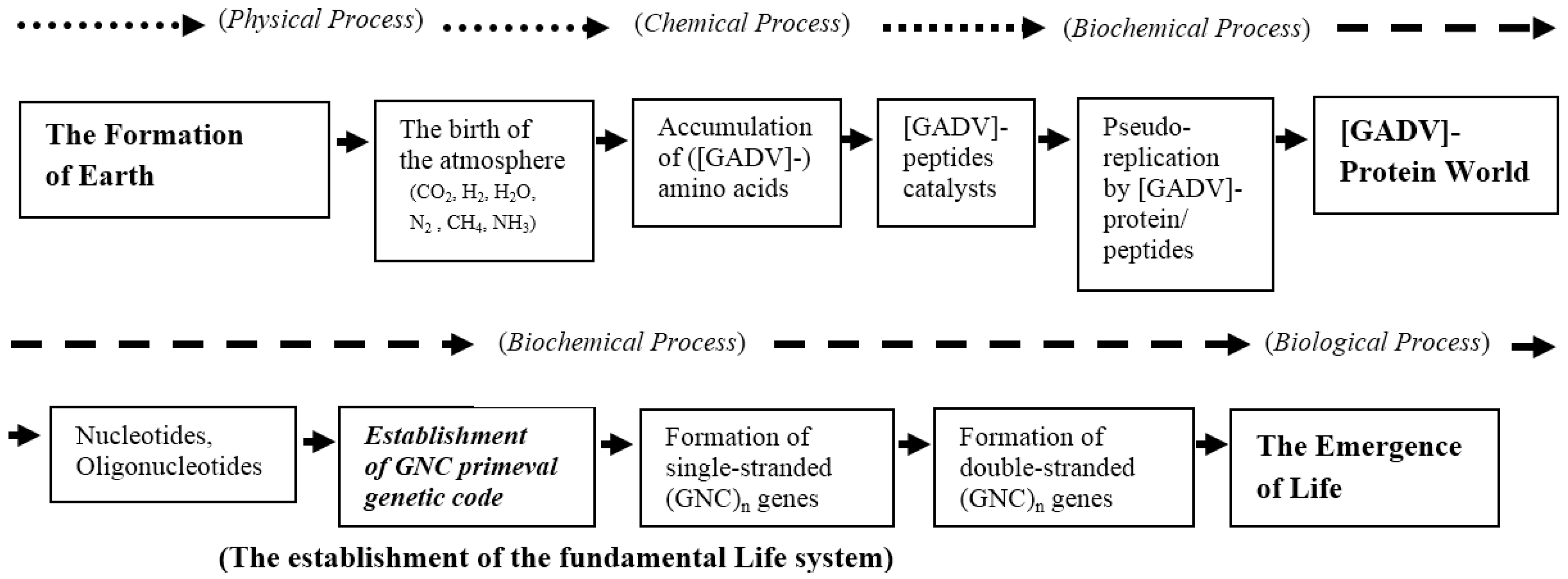
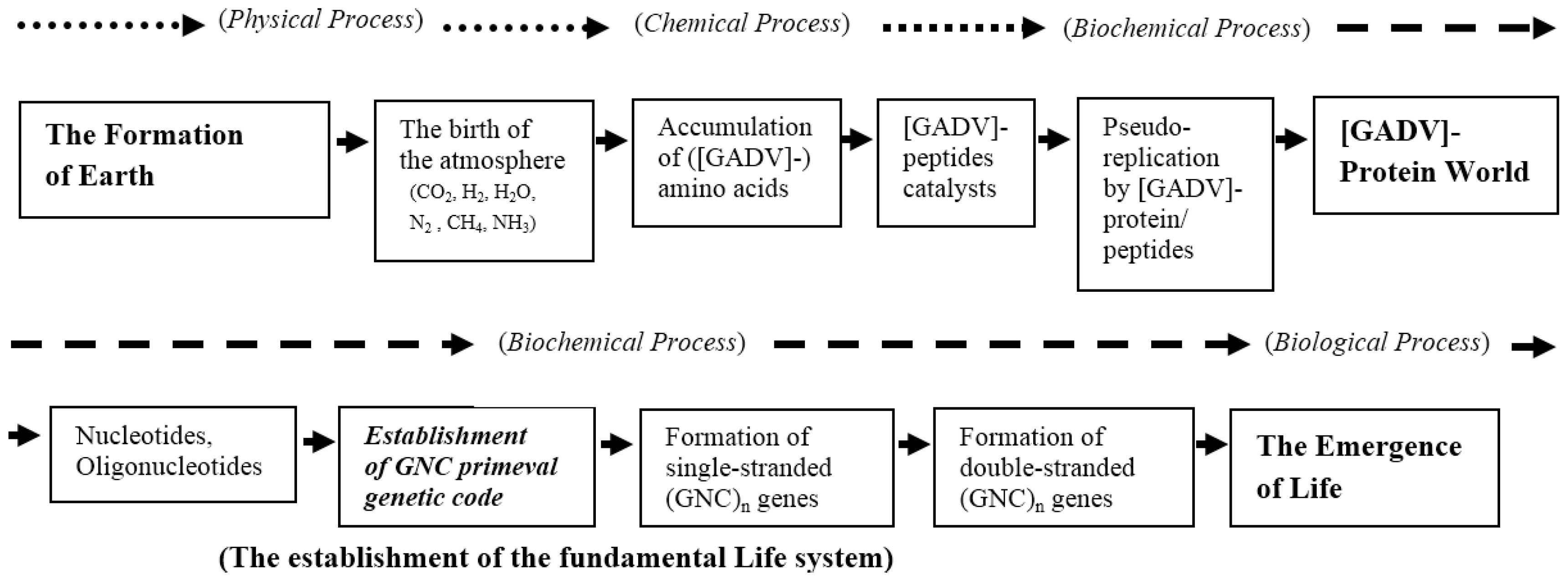
4.3. Deduced Evolutionary Steps from the Formation of Earth to the Emergence of Life
5. Discussion
5.1. Why Can [GADV]-Amino Acids Accumulate by Prebiotic Means?
5.2. Why Can Water-Soluble Globular Proteins Be Formed in the Absence of Any Genetic System?
5.3. Life Could Emerge with Some Good Luck and Trial and Error
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Gilbert, W. The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of ribosomal RNA intervening sequence of Tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Miller, S.L.; Orgel, L.E. The Origins of Life on the Earth; Prentice-Hall: Englewood Cliffs, NJ, USA, 1974. [Google Scholar]
- Francis, B.R. The Hypothesis that the genetic code originated in coupled synthesis of proteins and the evolutionary predecessors of nucleic acids in primitive cells. Life 2015, 5, 467–505. [Google Scholar] [CrossRef] [PubMed]
- Van der Gulik, P.T.; Speijer, D. How amino acids and peptides shaped the RNA world. Life 2015, 5, 230–246. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L. Prebiotic metabolic networks? Mol. Syst. Biol. 2014, 10, 729–730. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Caetano-Anollés, G. The evolutionary history of protein fold families and proteomes confirms that the archaeal ancestor is more ancient than the ancestors of other superkingdoms. BMC Evol. Biol. 2012, 12, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G. The RNA dreamtime: Modern cells feature proteins that might have supported a prebiotic polypeptide world but nothing indicates that RNA world ever was. Bioessays 2010, 32, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Maury, C.P. Self-propagating β-sheet polypeptide structures as prebiotic informational molecular entities: The amyloid world. Orig. Life Evol. Biosph. 2009, 39, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Maury, C.P. Origin of life. Primordial genetics: Information transfer in a pre-RNA world based on self-replicating beta-sheet amyloid conformers. J. Theor. Biol. 2015, 382, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.W. What RNA World? Why a Peptide/RNA Partnership Merits Renewed Experimental Attention. Life 2015, 5, 294–320. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, G. The place of RNA in the origin and early evolution of the genetic machinery. Life 2014, 4, 1050–1091. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L. A new start from ground zero? Orig. Life Evol. Biosph. 2014, 44, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A. Paradoxes in the origin of life. Orig. Life Evol. Biosph. 2014, 44, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Kvenvolden, K.A.; Peterson, E.; Pollock, G.E. Optical Configulation of Amino-Acids in Pre-Cambrian Fig Tree Chert. Nature 1969, 221, 141–143. [Google Scholar] [CrossRef]
- Nagy, B.; Engel, M.H.; Zumberge, J.E.; Ogiono, H.; Chang, S.Y. Amino acids and hydrocarbons ~3,800-Myr old in the Isua Rocks, southwestern Greenland. Nature 1981, 289, 53–56. [Google Scholar] [CrossRef]
- Van Zuilen, M.A.; Lepland, A.; Arrhenius, G. Reassessing the evidence for the earliest traces of life. Nature 2002, 418, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Botta, O.; Glavin, D.P.; Kminek, G.; Bada, J.L. Relative amino acid concentrations as a signature for parent body processes of carbonaceous chondrites. Orig. Life Evol. Biosph. 2002, 32, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Meierhenrich, U.J.; Muñoz Caro, G.M.; Bredehöft, J.H.; Jessberger, E.K.; Thiemann, W.H.-P. Identification of diamino acids in the Murchison meteorite. Proc. Natl. Acad. Sci. USA 2004, 101, 9182–9186. [Google Scholar] [CrossRef] [PubMed]
- Oró, J.; Mills, T.; Lazcano, A. Comets and the formation of biochemical compounds on the primitive Earth—A review. Orig. Life Evol. Biosph. 1992, 21, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Chyba, C.F.; Sagan, C. Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life. Nature 1992, 355, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L. Production of some organic compounds under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Plankensteiner, K.; Reiner, H.; Rode, B.M. Amino acids on the rampant primordial Earth: Electric discharges and the hot salty ocean. Mol. Divers. 2006, 10, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Kasting, J.F. Earth’s early atmosphere. Science 1993, 259, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Cleaves, H.J.; Chalmers, J.H.; Lazcano, A.; Miller, S.L.; Bada, J.L. A reassessment of prebiotic organic synthesis in neutral planetary atmosphere. Orig. Life Evol. Biosph. 2008, 38, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.T.; Cleaves, H.J.; Dworkin, J.P.; Glavin, D.P.; Callahan, M.; Aubrey, A.; Lazcano, A.; Bada, J.L. Primodial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proc. Natl. Acad. Sci. USA 2011, 108, 5526–5531. [Google Scholar] [CrossRef] [PubMed]
- Corliss, J.B.; Dymond, J.; Gordon, L.I.; Edmond, J.M.; Herzen, R.P.; Ballard, R.D.; Green, K.; Williams, D.; Bainbridge, A.; Crane, K.; et al. Submarine thermal springs on the galapagos rift. Science 1979, 203, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Holm, N.G.; Andersson, E. Hydrothermal simulation experiments as a tool for studies of the origin of life on Earth and other terrestrial planets: A review. Astrobiology 2005, 5, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Chandru, K.; Obayashi, Y.; Kaneko, T.; Kobayashi, K. Formation of amino acid condensates partly having peptide bonds in a simulated submarine hydrothermal environment. Viva Origin. 2013, 41, 24–28. [Google Scholar]
- Imai, E.; Honda, H.; Hatori, K.; Brack, A.; Matsuno, K. Elongation of oligopeptides in a simulated submarine hydrothermal system. Science 1999, 283, 831–833. [Google Scholar] [CrossRef] [PubMed]
- Ogata, Y.; Imai, E.; Honda, H.; Hatori, K.; Matsuno, K. Hydrothermal circulation of seawater through hot vents and contribution of interface chemistry to prebiotic synthesis. Orig. Life Evol. Biosph. 2000, 30, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Otake, T.; Taniguchi, T.; Furukawa, Y.; Kawamura, F.; Nakazawa, H.; Kakegawa, T. Stability of amino acids and their oligomerization under high-pressure conditions: implications for prebiotic chemistry. Astrobiology 2011, 11, 799–813. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, Y.; Sekine, T.; Oba, M.; Kakegawa, T.; Nakazawa, H. Biomolecule formation by oceanic impacts on early Earth. Nat. Geosci. 2009, 2, 62–66. [Google Scholar] [CrossRef]
- Ebihara, M.; Sekimoto, S.; Shirai, N.; Hamajima, Y.; Yamamoto, M.; Kumagai, K.; Oura, Y.; Ireland, T.R.; Kitajima, F.; Nagao, K.; et al. Neutron activation analysis of a particle returned from asteroid Itokawa. Science 2011, 333, 1119–1121. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Hilchenbach, M.; Langevin, Y.; Kissel, J.; Silen, J.; Briois, C.; Engrand, C.; Hornung, K.; Baklouti, D.; Bardyn, A.; et al. Comet 67P/Churyumov-Gerasimenko sheds dust coat accumulated over the past four years. Nature 2015, 518, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Nummelin, A.; Dickens, J.E.; Bergman, P.; Hjalmarson, A.; Irvine, W.M.; Ikeda, M.; Ohishi, M. Abundances of ethylene oxide and acetaldehyde in hot molecular cloud cores. Astron. Astrophys. 1998, 337, 275–286. [Google Scholar] [PubMed]
- Zaia, D.A.; Zaia, C.T.; de Santana, H. Which amino acids should be used in prebiotic chemistry studies? Orig. Life Evol. Biosph. 2008, 38, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, G. Evolution of the first metabolic cycles. Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Rode, B.M. Peptides and the origin of life. Peptides 1999, 20, 773–786. [Google Scholar] [CrossRef]
- Shuwannachot, Y.; Rode, B.M. Catalysis of dialanine formation by glycine in the salt-induced peptide formation reaction. Orig. Life Evol. Biosph. 1998, 28, 79–90. [Google Scholar] [CrossRef]
- Plankensteiner, K.; Reiner, H.; Rode, B.M. Catalytically increased prebiotic peptide formation: Ditryptophan, dilysine, and diserine. Orig. Life Evol. Biosph. 2005, 35, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Gorlero, M.; Wieczorek, R.; Adamala, K.; Giorgi, A.; Schininà, M.E.; Stano, P.; Luisi, P.L. Ser-His catalyses the formation of peptides and PNAs. FEBS Lett. 2009, 583, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, R.; Dörr, M.; Chotera, A.; Luisi, P.L.; Monnard, P.A. Formation of RNA Phosphodiester Bond by Histidine-Containing Dipeptides. ChemBioChem 2013, 14, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oba, T.; Fukushima, J.; Maruyama, M.; Iwamoto, R.; Ikehara, K. Catalytic activities of [GADV]-peptiders. Formation and establishment of [GADV]-protein world for the emergence of life. Orig. Life Evol. Biosph. 2005, 35, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Van der Gulik, P.; Massar, S.; Gilis, D.; Buhrman, H.; Rooman, M. The First Peptides: The evolutionary transition between prebiotic amino acids and early proteins. J. Theor. Biol. 2009, 261, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Higgs, P.G. A four-column theory for the origin of the genetic code: Tracing the evolutionary pathways that gave rise to an optimized code. Biol. Direct. 2009, 24, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K.; Omori, Y.; Arai, R.; Hirose, A. A novel theory on the origin of the genetic code: A GNC-SNS hypothesis. J. Mol. Evol. 2002, 54, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Stryer, L. Biochemistry, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1988. [Google Scholar]
- Ikehara, K. Origins of gene, genetic code, protein and life: Comprehensive view of life system from a GNC-SNS primitive genetic code hypothesis. J. Biosci. 2002, 27, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K.; Amada, F.; Yoshida, S.; Mikata, Y.; Tanaka, A. A possible origin of newly-born bacterial genes: Significance of GC-rich nonstop frame on antisense strand. Nucleic Acids Res. 1996, 24, 4249–4255. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K.; Okazawa, E. Unusually biased nucleotide sequences on sense strands of Flavobacterium sp. genes produce nonstop frames on the corresponding antisense strands. Nucleic Acids Res. 1993, 21, 2193–2199. [Google Scholar] [PubMed]
- Ikehara, K. Simulation of gene evolution: Evidence for GC-NSF(a) hypothesis on the origin of genes. Viva Origin. 2003, 31, 201–215. [Google Scholar]
- Ikehara, K. Mechanisms for creation of “original ancestor genes”. J. Biol. Macromol. 2005, 5, 21–30. [Google Scholar]
- Ikehara, K.; Yoshida, S. SNS hypothesis on the origin of the genetic code. Viva Origin. 1996, 26, 301–310. [Google Scholar]
- Ikehara, K. Pseudo-replication of [GADV]-proteins and origin of life. Int. J. Mol. Sci. 2009, 10, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K. Possible steps to the emergence of life: The [GADV]-protein world hypothesis. Chem. Rec. 2005, 5, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K. [GADV]-Protein World Hypothesis on the Origin of Life. In Genesis—in the Beginning; Seckbach, J., Ed.; Springer: Berlin, Germany, 2012; pp. 107–121. [Google Scholar]
- Ikehara, K. [GADV]-protein world hypothesis on the origin of life. Orig. Life Evol. Biosph. 2014, 44, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, K. Protein ordered sequences are formed by random joining of amino acids in protein 0th-order structure, followed by evolutionary process. Orig. Life Evol. Biosph. 2014, 44, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M. An extension of the coevolution theory of the origin of the genetic code. Biol. Direct 2008, 3, 37. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikehara, K. Evolutionary Steps in the Emergence of Life Deduced from the Bottom-Up Approach and GADV Hypothesis (Top-Down Approach). Life 2016, 6, 6. https://doi.org/10.3390/life6010006
Ikehara K. Evolutionary Steps in the Emergence of Life Deduced from the Bottom-Up Approach and GADV Hypothesis (Top-Down Approach). Life. 2016; 6(1):6. https://doi.org/10.3390/life6010006
Chicago/Turabian StyleIkehara, Kenji. 2016. "Evolutionary Steps in the Emergence of Life Deduced from the Bottom-Up Approach and GADV Hypothesis (Top-Down Approach)" Life 6, no. 1: 6. https://doi.org/10.3390/life6010006