Anaerobic Thermophiles

Abstract
:1. Introduction



2. Growth Conditions

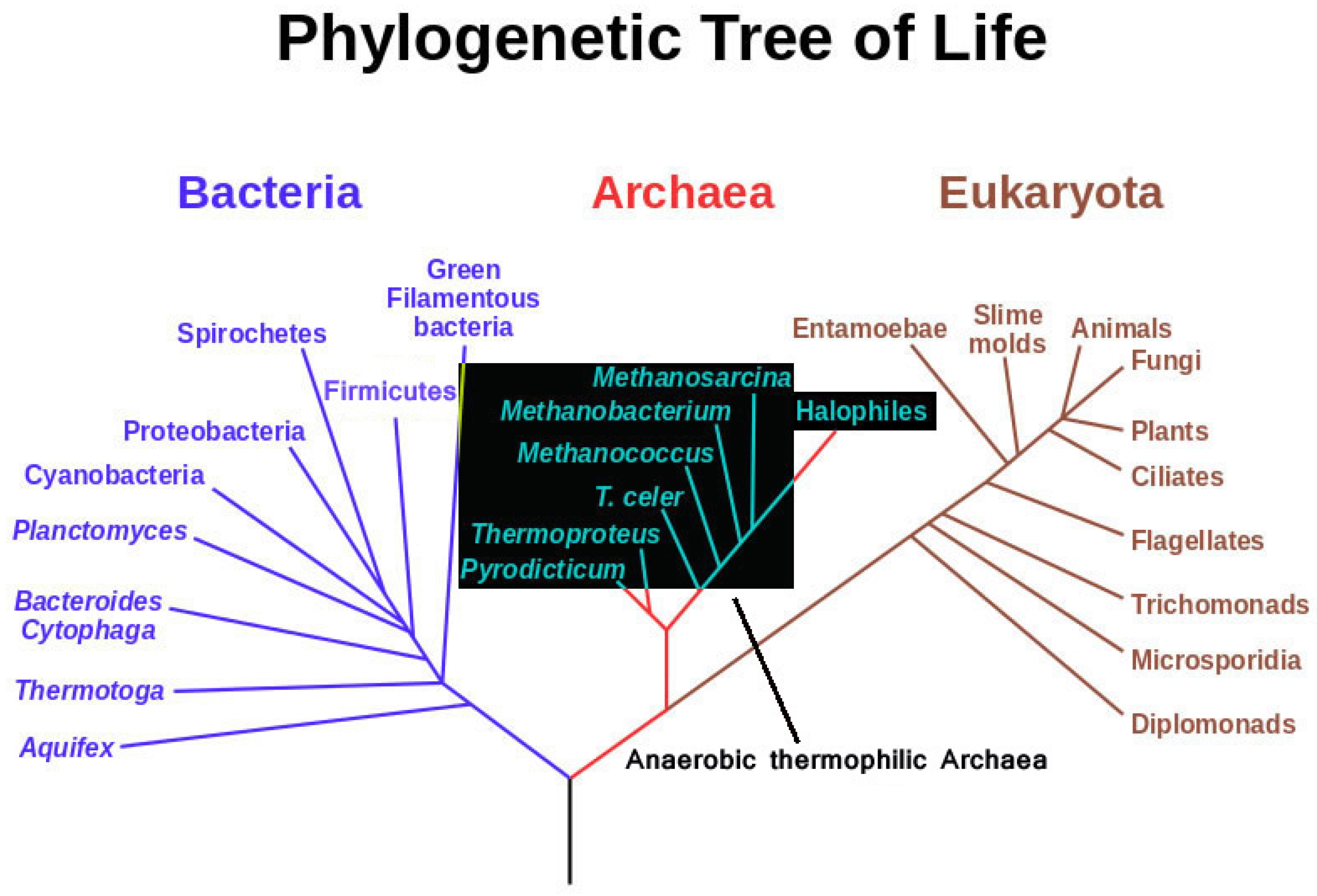{kind=link}
{kind=link}
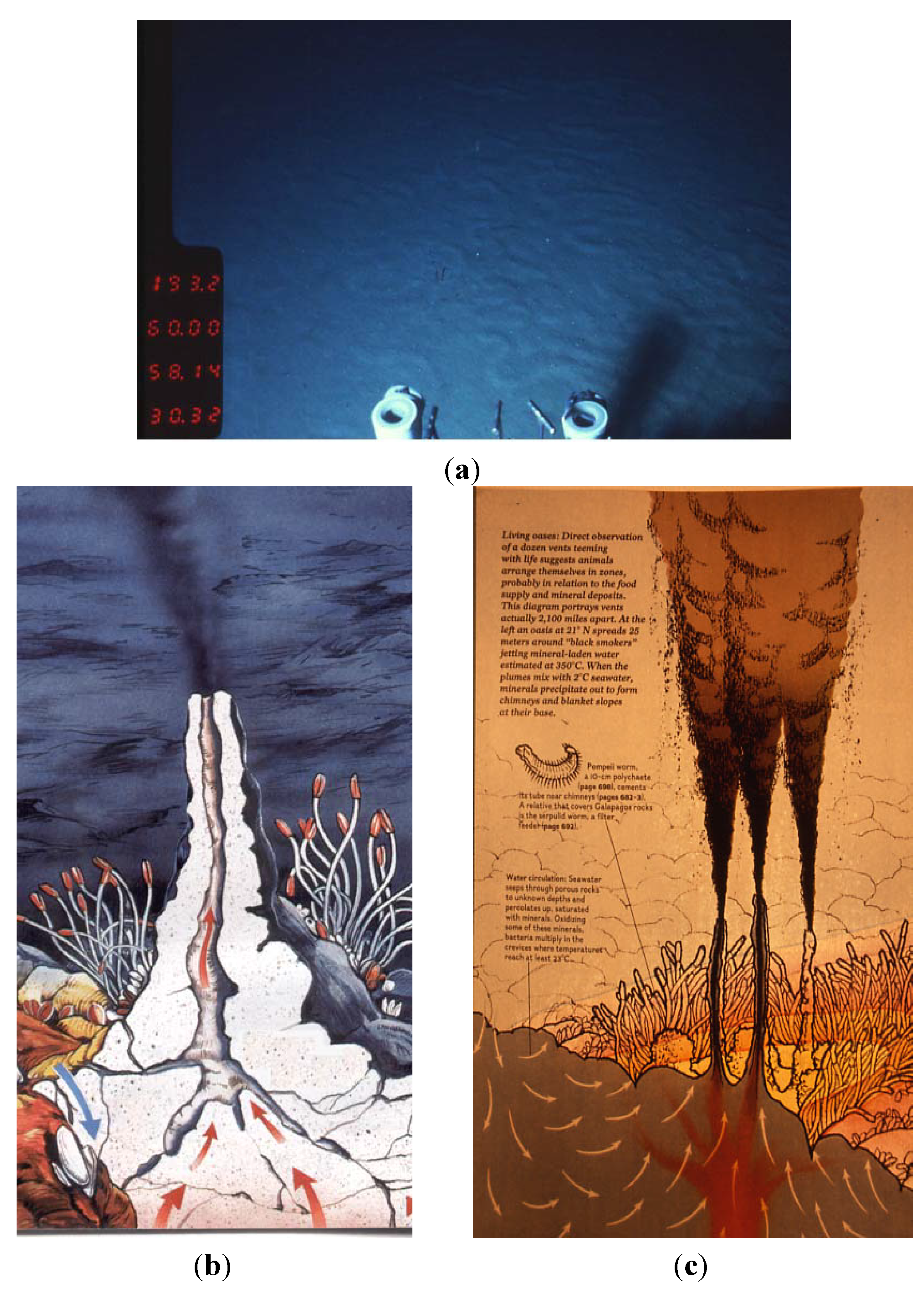{kind=link}
| Species | O2-relationship and metabolism | Temperature range (optimum) | pH range (optimum) | Originally isolated from |
|---|---|---|---|---|
| Bacteria; Proteobacteria; Gammaproteobacteria; Chromatiales; Chromatiaceae; | ||||
| Genus: Thermochromatium | ||||
| Thermochromatium tepidum | AN | 34–57 (48–50) | (7) | Mammoth Hot Spring, Yellowstone National Park, USA |
| PA/PH | ||||
| Bacteria; Bacteroidetes/Chlorobi group; Bacteroidetes; Bacteroidetes; Bacteroidales; Bacteroidaceae; | ||||
| Genera: Acetomicrobium, Anaerophaga | ||||
| Anaerophaga sp. strain HPS1 | NR | NR | NR | Offshore hot spring sediment, China |
| Bacteria; Spirochaetes; Spirochaetes; Spirochaetales; Spirochaetaceae; | ||||
| Genus: Spirochaeta, Exilispira | ||||
| Spirochaeta thermophila | AN | 40–73 (66–68) | 5.9–7.7 (7.5) | Marine hot spring on the beach of an island of Kamchatka, also from a hot spring on Raoul Island, New Zealand |
| COH | ||||
| Bacteria; Firmicutes; Clostridia; Halanaerobiales; Halanaerobiaceae; | ||||
| Genus: Halothermothrix | ||||
| Halothermothrix orenii | AN | 45–68 (60) | 5.5–8.2 (6.5–7) | Chott El Guettar hypersaline lake, Tunisia |
| COH | ||||
| Bacteria; Firmicutes; Clostridia; Natranaerobiales; Natranaerobiaceae; | ||||
| Genus: Natranaerobius | ||||
| Natranaerobius thermophilus | AN | 35–56 (53) | 8.5–10.6 (9.5) | Sediment of alkaline, hypersaline lakes of the Wadi An Natrun |
| COH | ||||
| Bacteria; Firmicutes; Clostridia; Thermoanaerobacteriales; Thermoanaerobacteriaceae; Syntrophomonadaceae; | ||||
| Genera: Coprothermobacter, Gelria, Moorella, Thermacetogenium, Mahella, Thermoanaerobacterium, Thermoanaerobacter, Thermosediminibacter, Caldanaerobacter, Thermovenabulum, Tepidanaerobacter, Ammonifex, Thermanaeromonas, Thermhydrogenium, Caldanaerovirga, Fervidicola, Caldanaerobius | ||||
| Coprothermobacter proteolyticus | AN | 35–70 (63) | 5–8.5 (7.5) | Thermophilic digester fermenting tannery wastes and cattle manure |
| COH | ||||
| Moorella thermoacetica | AN | 45–65 (55–60) | NR | Horse manure |
| COH | ||||
| Thermoanaerobacter ethanolicus | AN | 37–78 (69]) | 4.4–9.9 (5.8–8.5) | Hot springs, Yellowstone National Park, USA |
| COH | ||||
| Thermoanaerobacter pseudoethanolicus | AN | (65) | NR | Hot Spring, Yellowstone National Park, USA |
| COH | ||||
| Caldanaerobacter subterraneus subsp. tengcongensis | AN | 50–80 (75) | 5.5–9 9 (7–7.5) | Hot spring, Tengcong, China |
| COH | ||||
| Ammonifex degensii | AN | 57–77 (70) | 5–8 (7.5) | Kawah Candradimuka crater, Dieng Plateau, Java, Indonesia |
| F-CLA | ||||
| Bacteria; Firmicutes; Clostridia; Clostridiales; Acidaminococcaceae; | ||||
| Genus: Thermosinus | ||||
| Thermosinus carboxydivorans | AN | 40–68 (60) | 6.5–7.6 (6.8–7) | Norris Basin hot spring, Yellowstone National Park, USA |
| CLA | ||||
| Bacteria; Firmicutes; Clostridia; Clostridiales; Peptococcaceae; | ||||
| Genera: Desulfotomaculum, Pelotomaculum, Carboxydothermus, Thermincola | ||||
| Pelotomaculum thermopropionicum | AN | 45–65 (55) | 6.7–7.5 (7) | Thermophilic upflow anaerobic sludge blanket reactor |
| COH | ||||
| Carboxydothermus hydrogenoformans | AN | 40–78 (70-72) | 6.4–7.7 (6.8–7) | Freshwater hydrothermal springs, Kunashir Island, Kamchatka, Russia |
| CLA | ||||
| Bacteria; Firmicutes; Clostridia; Clostridiales; Syntrophomonadaceae; | ||||
| Genera: Anaerobaculum, Syntrophothermus, Thermanaerovibrio, Carboxydocella, Anaerobranca, Thermosyntropha, Caldicellulosiruptor | ||||
| Caldicellulosiruptor lactoaceticus | AN | 50–78 (68) | 5.8–8.2 (7) | Hveragerði alkaline hot spring, Iceland |
| COH | ||||
| Caldicellulosiruptor owensensis | AN | 50–80 (75) | 5.5–9 (7.5) | Freshwater pond within the dry Owens Lake bed, California, USA |
| COH | ||||
| Caldicellulosiruptor kristjanssonii | AN | 45–82 (78) | 5.8–8 (7) | Hot spring, Iceland |
| COH | ||||
| Caldicellulosiruptor saccharolyticus | AN | 45–80 (70) | 5.5–8.0 (7.0) | Geothermal spring, Taupo, New Zealand |
| COH | ||||
| Caldicellulosiruptor bescii | AN | |||
| COH | ||||
| Caldicellulosiruptor kronotskyensis | AN | (70) | (7) | Hot spring, Kamchatka, Russia |
| COH | ||||
| Caldicellulosiruptor hydrothermalis | AN | (65) | (7) | Hot spring, Kamchatka, Russia |
| COH | ||||
| Bacteria; Firmicutes; Clostridia; Clostridiales; Heliobacteriaceae; | ||||
| Genus: Heliobacterium | ||||
| Heliobacterium modesticaldum | AN | 25–56 (52) | (6–7) | Iceland, Yellowstone National Park, USA |
| PH & COH | ||||
| Bacteria; Firmicutes; Clostridia; Clostridiales; Clostridiaceae; Caldicoprobacteraceae; Veillonellaceae; | ||||
| Genera: Alkaliphilus, Clostridium, Tepidibacter, Caloramator, Garciella, Caminicella, Caloranaerobacter, Thermobrachium, Thermohalobacter, Tepidimicrobium, Fervidicella, Caldicoprobacter, Sporolituus, Thermotalea, Lutispora | ||||
| Clostridium thermocellum | AN | 28–69 (60) | (6.1–7.5) | Louisiana cotton bale and Compost heap |
| COH | ||||
| Clostridium stercorarium subsp. stercorarium | AN | (65) | (7.3) | Compost heap |
| COH | ||||
| Bacteria; Firmicutes; Bacilli; Bacillales; Bacillaceae; | ||||
| Genera: Anoxybacillus, Bacillus, Geobacillus, Vulcanibacillus | ||||
| Anoxybacillusflavithermus | FAE | 30–72 (60–65) | 5.5–9 (7) | A hot spring, New Zealand |
| COH | ||||
| Geobacillus thermodenitrificans | FAE | 45–70 | 6–8 | Sugar beet juice from extraction installations; Austria |
| COH | ||||
| Geobacillus thermoleovorans | FAE | 35–78 (55–65) | (6.2–6.8) | Soil near hot water effluent, Bethlehem, PA, USA |
| COH | ||||
| Geobacillus thermoglucosidiasus | FAN | 40–70 (60) | 6–9 (7) | Japan soil |
| COH | ||||
| Bacteria; Proteobacteria; delta/epsilon subdivisions; Deltaproteobacteria; Desulfurellales; Desulfurellaceae; | ||||
| Genera: Desulfurella, Hippea | ||||
| Hippea maritima | AN | 40–65 (52–54) | 5.4–6.5 (5.8–6.2) | Shallow water hot vents, Bay of Plenty, New Zealand and Matupi Harbour, Papua New Guinea |
| COH | ||||
| Bacteria; Proteobacteria; delta/epsilon subdivisions; Epsilonproteobacteria; Nautiliales; Nautiliaceae; | ||||
| Genera: Nautilia, Lebetimonas, Caminibacter | ||||
| Caminibacter mediatlanticus | AN | 45–70 (55) | 4.5–7.5 (5.5) | “Rainbow” deep-sea vent field, Mid-Atlantic Ridge |
| CLA | ||||
| Bacteria; Deferribacteres; Deferribacteres; Deferribacterales; Deferribacteraceae; | ||||
| Genera: Deferribacter; Flexistipes also, Caldithrix (unclassified Deferribacteres), Calditerrivibrio | ||||
| Deferribacter desulfuricans | AN | 40–70 (60–65) | 5.0–7.5 (6.5) | From a black smoker vent from the hydrothermal fileds at the Suiyo Seamount in the Izu-Bonin Arc, Japan |
| COH | ||||
| Bacteria; Thermodesulfobacteria; Thermodesulfobacteria; Thermodesulfobacteriales; Thermodesulfobacteriaceae; | ||||
| Genera: Thermodesulfatator, Thermodesulfobacterium, Caldimicrobium, Thermosulfidibacter | ||||
| Thermodesulfatator indicus | AN | 55–80 (70) | 6–6.7 (6.25) | The Kairei deep-sea hydrothermal vent field, Central Indian Ridge |
| CLA | ||||
| Thermodesulfobacterium commune | AN | 50–85 (70) | 6.0–8.0 | Ink Pot Spring, Yellowstone National Park, USA |
| COH | ||||
| Bacteria; Nitrospirae; Nitrospira; Nitrospirales; Nitrospiraceae; | ||||
| Genus: Thermodesulfovibrio | ||||
| Thermodesulfovibrio yellowstonii | AN | 40–70 (65) | (6.8–7) | Thermal vent, Yellowstone National Park, USA |
| COH | ||||
| Bacteria; Dictyoglomi; Dictyoglomi; Dictyoglomales; Dictyoglomaceae; | ||||
| Genus: Dictyoglomus | ||||
| Dictyoglomus thermophilum | AN | 50–80 (73–78) | 5.9–8.3 (7) | Hot spring, Kumamoto Prefecture, Japan |
| COH | ||||
| Bacteria; Chloroflexi; Chloroflexi; Chloroflexales; Chloroflexaceae; | ||||
| Genera: Roseiflexus, Chloroflexus, Heliothrix | ||||
| Roseiflexus castenholzii | FAE | 45–55 (50) | 7–9 (7.5–8) | Hot spring, Nakabusa, Japan |
| PH (anaerobic) | ||||
| Chloroflexus aggregans | FAE | (50–60) | 7.0–9.0 | Hot spring of the Okukinu Meotobuchi hot spring in Tochigi Perfecture, Japan |
| PH (anaerobic) | ||||
| Chloroflexus aurantiacus | FAE | (52–60) | (8) | Hot spring in the canyon at Sokokura, Hakone district, Japan |
| PH (anaerobic) | ||||
| Heliothrix oregonensis | FAE | (40–55) | NR | Hot spring near Warm Springs River, Oregon, USA |
| PH | ||||
| Bacteria; Chloroflexi; Thermomicrobia; Thermomicrobiales; Thermomicrobiaceae; | ||||
| Genus: Thermomicrobium | ||||
| Thermomicrobium roseum | AN | (70–75) | 6–9.4 (8.2–8.5) | Hot spring, Yellowstone National Park, USA |
| COH | ||||
| Bacteria; Aquificae; Aquificae; Aquificales; Aquificaceae; | ||||
| Genera: Hydrogenivirga, Aquifex, Desulfurobacterium (unclassified Aquificales), Balnearium (unclassified Aquificales), Thermovibrio (unclassified Aquificales) | ||||
| Desulfurobacterium thermolithotrophum | AN | 40–75 (70) | 4.4–7.5 (6) | “Snake Pit” vent field, Mid-Atlantic ridge |
| CLA | ||||
| Thermovibrio ammonificans | AN | 60–80 (75) | 5–7 (5.5) | Deep sea hydrothermal vent area, East Pacific Rise |
| CLA | ||||
| Bacteria; Aquificae; Aquificae; Aquificales; Hydrogenothermaceae; | ||||
| Genera: Hydrogenothermus, Sulfurihydrogenibium, Persephonella | ||||
| Sulfurihydrogenibium azorense | FAE | 50–73 (68) | 5.5–7 (6) | Near the Água do Caldeirão, Furnas, on São Miguel Island, Azores |
| CLA | ||||
| Bacteria; Thermotogae; Thermotogae; Thermotogales; Thermotogaceae; | ||||
| Genera: Geotoga, Marinitoga, Petrotoga, Thermosipho, Thermotoga, Fervidobacterium, Thermococcoides, Kosmotoga | ||||
| Marinitoga camini | AN | 25–65 (55) | 5–9 (7) | Deep sea vent fields, Mid-Atlantic ridge |
| COH | ||||
| Petrotoga mobilis | AN | 40–65 (58–60) | 5.5–8.5 (6.5–7) | Oil reservoir production water from off-shore oil platforms, North Sea |
| COH | ||||
| Thermosipho melanesiensis | AN | 45–80 (70) | 3.5–9.5 (6.5–9.5) | Deep sea hydrothermal area, Lau Basin, southwest Pacific Ocean |
| COH | ||||
| Fervidobacterium nodosum | AN | 41–79 (70) | 6–8 (7) | Hot spring in New Zealand |
| COH | ||||
| Thermotoga lettingae | AN | 50–75 (65) | 6–8.5 (7) | Thermophilic, sulfate-reducing, slightly saline bioreactor |
| COH | ||||
| Thermotoga maritima | AN | 55–90 (80) | 5.5–9 (6.5) | Geothermally heated sea floors, Italy and the Azores |
| COH | ||||
| Thermotoga petrophila | AN | 47–88 (80) | 5.2–9 (7) | Production fluid of the Kubiki oil reservoir in Niigata, Japan |
| COH | ||||
| Thermotoga neapolitana | AN | 55–90 (80) | 5.5–9 (7) | Shallow submarine hot springs, Lucrino Bay, Naples, Italy |
| COH | ||||
| Archaea; Crenarchaeota; Thermoprotei; Desulfurococcales; Desulfurococcaceae | ||||
| Genera: Acidilobus, Staphylothermus, Ignicoccus, Desulfurococcus Thermosphaera, Sulfophobococcus, Stetteria, Thermodiscus, (Also Ignisphaera of the Ignisphaera group) | ||||
| Thermosphaera aggregans | AN | 65–90 (85) | 5–7 (6.5) | “Obsidian Pool” Yellowstone National Park, USA |
| COH | ||||
| Staphylothermus marinus | AN | 65–98 (92) | 4.5–8.5 (6.5) | Vulcano Island, Italy, also a deep-sea black smoker of the East Pacific Rise |
| COH | ||||
| Archaea; Crenarchaeota; Thermoprotei; Desulfurococcales; Pyrodictiaceae; | ||||
| Genera: Pyrodictium, Hyperthermus, Pyrolobus | ||||
| Hyperthermus butylicus | AN | (95–107) | (7) | Hydrothermally heated flat-sea sediments off the coast of São Miguel Island, Azores |
| COH | ||||
| Archaea; Crenarchaeota; Thermoprotei; Thermoproteales; Thermofilaceae | ||||
| Genus: Thermofilum | ||||
| Thermofilum pendens | AN | (85–90) | (5) | Icelandic solfataras |
| COH | ||||
| Archaea; Crenarchaeota; Thermoprotei; Thermoproteales; Thermoproteaceae | ||||
| Genera: Thermoproteus, Pyrobaculum, Thermocladium, Caldvirga | ||||
| Thermoproteus neutrophilus | AN | (85) | (6.8) | Hot spring, Iceland |
| F-CLA | ||||
| Pyrobaculum arsenaticum | AN | 68–100 (81) | NR | Hot water pond, Pisciarelli Solfatara, Naples, Italy |
| F-CLA | ||||
| Pyrobaculum islandicum | AN | 74–102 (100) | 5–7 (6) | Boiling solfataras and geothermal waters, Iceland |
| F-CLA | ||||
| Pyrobaculum calidifontis | FAE | 75–100 (90-95) | 5.5–8.0 (7.0) | Terrestrial hot spring Calamba, Laguna, the Philippines |
| COH | ||||
| Pyrobaculum aerophilum | FAE | 75–104 (100) | 5.8–9 (7) | Boiling marine water hole, Maronti Beach, Ischia, Italy |
| F-CLA | ||||
| Caldivirga maquilingensis | FAE | 62–92 (85) | 2.3–6.4 (3.7–4.2) | Acidic hot spring in the Philippines |
| COH | ||||
| Archaea; Euryarchaeota; Thermoplasmata; Thermoplasmatales; Thermoplasmataceae; | ||||
| Genus: Thermoplasma; Acidiplasma | ||||
| Thermoplasma acidophilum | FAE | 45–63 (59) | 0.5–4 (1–2) | Solfatara fields and self heated coal refuse piles |
| COH | ||||
| Thermoplasma volcanium | FAE | 33–67 (60) | 1–4 (2) | Submarine and continental solfataras at Vulcano Island, Italy; also from Java, Iceland and Yellowstone National Park, USA |
| COH | ||||
| Archaea; Euryarchaeota; Methanococci; Methanococcales; Methanocaldococcaceae | ||||
| Genera: Methanocaldococcus, Methanotorris | ||||
| Methanocaldococcus jannaschii | AN | 50–86 (85) | 5.2–7.0 (6.0) | “White smoker” chimney on the 20°N East Pacific Rise |
| CLA | ||||
| Methanocaldococcus vulcanius | AN | 49–89 (80) | 5.2–7 (6.5) | Deep-sea vent, 13°N thermal field, East Pacific Rise |
| CLA | ||||
| Archaea; Euryarchaeota; Thermococci; Thermococcales; Thermococcaceae; | ||||
| Genera: Thermococcus, Pyrococcus, Palaeococcus | ||||
| Thermococcus barophilus | AN | 48–100 (85) | (7) | “Snakepit” hydrothermal vent region of the Mid-Atlantic ridge |
| COH | ||||
| Thermococcus gammatolerans | AN | 55–95 (88) | (6) | Guaymas Basin, Gulf of California |
| COH | ||||
| Thermococcus kodakarensis | AN | 60–100 (85) | 5–9 (6.5) | Solfatara on Kodakara Island, Kagoshima, Japan |
| COH | ||||
| Thermococcus sibiricus | AN | 40–88 (78) | 5.8–9 (7.5) | Samotlor oil reservoir, Western Siberia |
| COH | ||||
| Pyrococcus furiosus | AN | 70–103 (100) | 5–9 (7) | Shallow marine hydrothermal system at Vulcano Island, Italy |
| COH | ||||
| Pyrococcus horikoshii | AN | 80–102 (98) | 5–8 (7) | Hydrothermal fluid samples obtained at the Okinawa Trough vents in the NE Pacific Ocean, at a depth of 1395 m |
| COH | ||||
| Archaea; Euryarchaeota; Archaeoglobi; Archaeoglobales; Archaeoglobaceae; | ||||
| Genera: Archeoglobus, Geoglobus, Ferroglobus | ||||
| Archaeoglobus fulgidus | AN | 64–92 (83) | 5.5–7.5 | Marine hydrothermal systems at Vulcano island and at Stufe di Nerone, Naples, Italy |
| F-CLA | ||||
| Archaea; Euryarchaeota; Methanopyri; Methanopyrales; Methanopyraceae; | ||||
| Genus: Methanopyrus | ||||
| Methanopyrus kandleri | AN | 84–110 (98) | 5.5–7 (6.5) | Deep-sea sediment from the Guaymas Basin, Gulf of California, and from the shallow marine hydrothermal system of the Kolbeinsey ridge, Iceland |
| CLA | ||||
| Archaea; Euryarchaeota; Methanobacteria; Methanobacteriales; Methanobacteriaceae; | ||||
| Genera: Methanobacterium, Methanothermobacter | ||||
| Methanothermobacter thermautotrophicus | AN | 40–75 (65–70) | 6.0–8.8 (7.2–7.6) | Anaerobic sewage sludge digestor |
| CLA | ||||
| Archaea; Euryarchaeota; Methanococci; Methanococcales; Methanococcaceae; | ||||
| Genus: Methanothermococcus | ||||
| Methanothermococcus thermolithotrophicus | AN | 30–70 (65) | 6–8 (7) | Heated sea sediments near Naples, Italy |
| CLA | ||||
| Archaea; Euryarchaeota; Methanomicrobia; Methanosarcinales; Methanosaetaceae; Methanocellales | ||||
| Genus: Methanothrix | ||||
| Methanothrix thermophila | AN | (55) | 6.1–7.5 (6.7) | Mesophilic anaerobic sludge digestors |
| COH | ||||
| Archaea; Euryarchaeota; Methanomicrobia; Methanosarcinales; Methanosaetaceae; Methanocellales | ||||
| Genus: Methanocella | ||||
| Methanocella conradii | AN | 37–60 (55) | 6.4–7.2 (6.8) | Rice field soil |
| CLA | ||||
3. Metabolism and Biotechnological Applications
| Enzymes, organic compounds and processes | Applications and products | Most representing Genera |
|---|---|---|
| Amylases and pullulanases | Glucose, fructose for sweeteners; polymer-degrading additives in detergents | Pyrococcus, Thermococcus, Fervidobacterium, Dictyoglomus, Anaerobranca |
| Cellulases and Xylanases | Paper bleaching | Clostridium, Petrotoga, Thermotoga, Thermosypho, Moorella, Caldicoprobacter, Caldicellulosiruptor |
| Proteases | Amino acid production from keratins, food processing, baking, brewing, detergents | Thermoanaerobacter, Fervidobacterium |
| DNA-polymerases and ligases | Genetic engineering | Thermotoga, Pyrococcus, Thermococcus, Archaeoglobus, Thermoanaerobacter |
| Ethanol | Chemical and food industries | Clostridium, Thermoanaerobacter; Thermoanaerobacterium, Caldanaerobius, Caloramator |
| Hydrogen and/or methane | Energy, fuels | Clostridium, Carboxydocella, Thermincola, Thermosinus, Thermotoga, Carboxydothermus, Carboxydobrachium, Anaerobaculum, Methanotorris, Methanococcus, Methanothermococcus, Methanotermobacter |
| Volatile fatty Acids | Chemical and food industries | Clostridium |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thermophiles: The Keys to Molecular Evolution and the Origin of Life? Wiegel, J.; Michael, A.W.W. (Eds.) CRC Press: Boca Raton, FL, USA, 1998.
- Stahl, D.A.; Lane, D.J.; Olsen, G.J.; Pace, N.R. Characterization of a Yellowstone hot spring microbial community by 5S rRNA sequences. Appl. Environ. Microbiol. 1985, 49, 1379–1384. [Google Scholar]
- Sandbeck, K.A.; Ward, D.M. Temperature adaptations in the terminal processes of anaerobic decomposition of Yellowstone national park and icelandic hot spring microbial mats. Appl. Environ. Microbiol. 1982, 44, 844–851. [Google Scholar]
- Friedmann, E.I. Endolithic microbial life in hot and cold deserts. Orig. Life 1980, 10, 223–235. [Google Scholar] [CrossRef]
- Brock, T.D. High temperature systems. Annu. Rev. Ecol. Syst. 1970, 1, 191–220. [Google Scholar] [CrossRef]
- Kempner, E.S. Upper temperature limit of life. Science 1963, 142, 1318–1319. [Google Scholar] [CrossRef]
- Averhoff, B.; Müller, V. Exploring research frontiers in microbiology: Recent advances in halophilic and thermophilic extremophiles. Res. Microbiol. 2010, 161, 506–514. [Google Scholar] [CrossRef]
- Basu, S.; Sen, S. Turning a mesophilic protein into a thermophilic one: A computational approach based on 3D structural features. J. Chem. Inf. Model 2009, 49, 1741–1750. [Google Scholar] [CrossRef]
- Kim, M.S.; Weaver, J.D.; Lei, X.G. Assembly of mutations for improving thermostability of Escherichia coli AppA2 phytase. Appl. Microbiol. Biotechnol. 2008, 79, 751–758. [Google Scholar] [CrossRef]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef]
- Daniel, R.M.; Toogood, H.S.; Bergquist, P.L. Thermostable proteases. Biotechnol. Genet. Eng. Rev. 1996, 13, 51–100. [Google Scholar] [CrossRef]
- Shaw, A.J.; Podkaminer, K.K.; Desai, S.G.; Bardsley, J.S.; Rogers, S.R.; Thorne, P.G.; Hogsett, D.A.; Lynd, L.R. Metabolic engineering of a thermophilic bacterium to produce ethanol at high yield. Proc. Natl. Acad. Sci. USA 2008, 105, 13769–13774. [Google Scholar] [CrossRef]
- Bustard, M.T.; Burgess, J.G.; Meeyoo, V.; Wright, P.C. Novel opportunities for marine hyperthermophiles in emerging biotechnology and engineering industries. J. Chem. Technol. Biotechnol. 2000, 75, 1095–1109. [Google Scholar] [CrossRef]
- Cai, G.; Jin, B.; Saint, C.; Monis, P. Genetic manipulation of butyrate formation pathways in Clostridium Butyricum. J. Biotechnol. 2011, 155, 269–274. [Google Scholar] [CrossRef]
- Oshima, T.; Moriya, T. A preliminary analysis of microbial and biochemical properties of high-temperature compost. Ann. N. Y. Acad. Sci. 2008, 1125, 338–344. [Google Scholar]
- Nesbø, C.L.; Kumaraswamy, R.; Dlutek, M.; Doolittle, W.F.; Foght, J. Searching for mesophilic Thermotogales bacteria: “Mesotogas” in the wild. Appl. Environ. Microbiol. 2010, 76, 4896–4900. [Google Scholar] [CrossRef]
- Nesbø, C.L.; Bradnan, D.M.; Adebusuyi, A.; Dlutek, M.; Petrus, A.K.; Foght, J.; Doolittle, W.F.; Noll, K.M. Mesotoga prima gen. nov., sp. nov., the first described mesophilic species of the Thermotogales. Extremophiles 2012, 16, 387–393. [Google Scholar] [CrossRef]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 degree C and isotopically heavy CH sub-production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef]
- Summit, M.; Scott, B.; Nielson, K.; Mathur, E.; Baross, J. Pressure enhances thermal stability of DNA polymerase from three thermophilic organisms. Extremophiles 1998, 2, 339–345. [Google Scholar] [CrossRef]
- Pledger, R.J.; Crump, B.C.; Baross, J.A. A barophilic response by two hyperthermophilic, hydrothermal vent archaea: An upward shift in the optimal temperature and acceleration of growth rate at supra-optimal temperatures by elevated pressure. FEMS Microbiol. Ecol. 1994, 14, 233–242. [Google Scholar] [CrossRef]
- Di Giulio, M. A comparison of proteins from Pyrococcus furiosus and Pyrococcus abyssi: Barophily in the physicochemical properties of amino acids and in the genetic code. Gene 2005, 346, 1–6. [Google Scholar] [CrossRef]
- Marteinsson, V.T.; Birrien, J.-L.; Reysenbach, A.-L.; Vernet, M.; Dominique, M.; Gamacorta, A.; Messner, P.; Sleytr, U.B.; Prieur, D. Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 1999, 49, 351–359. [Google Scholar] [CrossRef]
- Canganella, F.; Gonzalez, J.M.; Yanagibayashi, M.; Kato, C.; Horikoshi, K. Pressure and temperature effects on growth and viability of the hyperthermophilic archaeon Thermococcus peptonophilus. Arch. Microbiol. 1997, 168, 1–7. [Google Scholar] [CrossRef]
- Adams, M.W.W. The biochemical diversity of life near and above 100 °C in marine environments. J. Appl. Microbiol. 1999, 85, 108S–117S. [Google Scholar] [CrossRef]
- Segerer, A.; Trincone, A.; Gahrtz, M.; Stetter, K. Stygiolobus azoricus gen. nov., sp. nov. represents a novel genus of anaerobic, extremely thermoacidophilic archaebacteria of the order Suvolobales. Int. J. Syst. Bacteriol. 1991, 41, 495–501. [Google Scholar] [CrossRef]
- Prokofeva, M.I.; Kostrikina, N.A.; Kolganova, T.V.; Tourova, T.P.; Lysenko, A.M.; Lebedinsky, A.V.; Bonch-Osmolovskaya, E.A. Isolation of the anaerobic thermoacidophilic crenarchaeote Acidilobus saccharovorans sp. nov. and proposal of Acidilobales ord. nov., including Acidilobaceae fam. nov. and Caldisphaeraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 3116–3122. [Google Scholar] [CrossRef]
- Itoh, T.; Suzuki, K.; Sanchez, P.C.; Nakase, T. Caldisphaera lagunensis gen. nov., sp. nov., a novel thermoacidophilic crenarchaeote isolated from a hot spring at Mt. Maquiling, Philippines. Int. J. Syst. Evol. Microbiol. 2003, 53, 1149–1154. [Google Scholar] [CrossRef]
- Wiegel, J. Anaerobic alkalithermophiles, a novel group of extremophiles. Extremophiles 1998, 2, 257–267. [Google Scholar] [CrossRef]
- Chung, D.; Cha, M.; Farkas, J.; Westpheling, J. Construction of a stable replicating shuttle vector for Caldicellulosiruptor species: Use for extending genetic methodologies to other members of this genus. PLoS One 2013. [Google Scholar] [CrossRef]
- Fukui, K.; Bessho, Y.; Shimada, A.; Yokoyama, S.; Kuramitsu, S. Thermostable mismatch-recognizing protein MutS suppresses nonspecific amplification during Polymerase Chain Reaction (PCR). Int. J. Mol. Sci. 2013, 14, 6436–6453. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Dissecting and engineering metabolic and regulatory networks of thermophilic bacteria for biofuel production. Biotechnol. Adv. 2013, 31, 827–837. [Google Scholar] [CrossRef]
- Frock, A.D.; Kelly, R.M. Extreme thermophiles: Moving beyond single-enzyme biocatalysis. Curr. Opin. Chem. Eng. 2012, 1, 363–372. [Google Scholar] [CrossRef]
- Merkel, A.Y.; Huber, J.A.; Chernyh, N.A.; Bonch-Osmolovskaya, E.A.; Lebedinsky, A.V. Detection of putatively thermophilic anaerobic methanotrophs in diffuse hydrothermal vent fluids. Appl. Environ. Microbiol. 2013, 79, 915–923. [Google Scholar] [CrossRef]
- Tsubouchi, T.; Shimane, Y.; Mori, K.; Usui, K.; Hiraki, T.; Tame, A.; Uematsu, K.; Maruyama, T.; Hatada, Y. Polycladomyces abyssicola gen. nov., sp. nov., a thermophilic filamentous bacterium isolated from hemipelagic sediment in Japan. Int. J. Syst. Evol. Microbiol. 2013, 63, 1972–1981. [Google Scholar] [CrossRef]
- Dang, H.; Luan, X.W.; Chen, R.; Zhang, X.; Guo, L.; Klotz, M.G. Diversity, abundance and distribution of amoA-encoding archaea in deep-sea methane seep sediments of the Okhotsk Sea. FEMS Microbiol. Ecol. 2010, 72, 370–385. [Google Scholar] [CrossRef]
- Canganella, F.; Wiegel, J. Extremophiles: From abyssal to terrestrial ecosystems and possibly beyond. Naturwissenschaften 2011, 98, 253–279. [Google Scholar] [CrossRef]
- Canganella, F.; Vettraino, A.M.; Trovatelli, L.D. The extremophilic bacteria: Ecology and agroindustrial applications. Ann. Microbiol. Enzimol. 1995, 45, 173–184. [Google Scholar]
- Kobayashi, T.; Koide, O.; Mori, K.; Shimamura, S.; Matsuura, T.; Miura, T.; Takaki, Y.; Morono, Y.; Nunoura, T.; Imachi, H.; et al. Phylogenetic and enzymatic diversity of deep subseafloor aerobic microorganisms in organics- and methane-rich sediments off Shimokita Peninsula. Extremophiles 2008, 12, 519–527. [Google Scholar] [CrossRef]
- Pagé, A.; Tivey, M.K.; Stakes, D.S.; Reysenbach, A.L. Temporal and spatial archaeal colonization of hydrothermal vent deposits. Environ. Microbiol. 2008, 10, 874–884. [Google Scholar]
- Blöchl, E.; Rachel, R.; Burggraf, S.; Hafenbradl, D.; Jannasch, H.W.; Stetter, K.O. Pyrolobus fumarii, gen. and sp. nov., represents a novel group of Archaea, extending the upper temperature limit for life to 113 degrees C. Extremophiles 1997, 1, 14–21. [Google Scholar] [CrossRef]
- Engle, M.; Li, Y.; Rainey, F.; DeBlois, S.; Mai, V.; Reichert, A.; Mayer, F.; Messmer, P.; Wiegel, J. Thermobrachium celere Y., gen. nov., sp. nov., a fast growing thermophilic, alkalitolerant, and proteolytic obligate anaerobe. Int. J. Syst. Bacteriol. 1996, 46, 1025–1033. [Google Scholar] [CrossRef]
- Abed, R.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013. [Google Scholar] [CrossRef]
- Bowers, K.J.; Mesbah, N.M.; Wiegel, J. Biodiversity of poly-extremophilic Bacteria: Does combining the extremes of high salt, alkaline pH and elevated temperature approach a physico-chemical boundary for life? Saline Syst. 2009. [Google Scholar] [CrossRef]
- Mueller, D.R.; Vincent, W.F.; Bonilla, S.; Laurion, I. Extremotrophs, extremophiles and broadband pigmentation strategies in a high arctic ice shelf ecosystem. FEMS Microbiol. Ecol. 2005, 53, 73–87. [Google Scholar]
- Lee, Y.J.; Wagner, I.D.; Brice, M.E.; Kevbrin, V.V.; Wiegel, J. Thermosediminibacter oceani gen. nov., sp. nov. and Thermosediminibacter litoriperuensis sp.nov., new anaerobic thermophilic bacteria isolated form Peru Margin. Extremophiles 2005, 9, 375–383. [Google Scholar] [CrossRef]
- Li, Y.; Mandelco, L.; Wiegel, J. Isolation and characterization of a moderately thermophilic anaerobic alkaliphile, Clostridium paradoxum, sp. nov. Int. J. Syst. Bacteriol. 1993, 43, 450–460. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information Taxonomy Database. Available online: http://www.ncbi.nlm.nih.gov/genomes/MICROBES (accessed on 10 January 2014).
- Amend, J.P.; Shock, E.L. Energetics of overall metabolic reactions of thermophilic and hyperthermophilic Archaea and Bacteria. FEMS Microbiol. Rev. 2001, 25, 175–243. [Google Scholar] [CrossRef]
- Amend, J.P.; Rogers, K.L.; Shock, E.L.; Gurrieri, S.; Inguaggiato, S. Energetics of chemolithoautotrophy in the hydrothermal system of Vulcano Island, southern Italy. Geobiology 2003, 1, 37–58. [Google Scholar] [CrossRef]
- Selig, M.; Xavier, K.B.; Santos, H.; Schönheit, P. Comparative analysis of Embden-Meyerhof and Entner-Doudoroff glycolytic pathways in hyperthermophilic archaea and the bacterium Thermotoga. Arch. Microbiol. 1997, 167, 217–232. [Google Scholar]
- Yang, S.J.; Kataeva, I.; Wiegel, J.; Yin, Y.; Dam, P.; Xu, Y.; Westpheling, J.; Adams, M.W. Classification of Anaerocellum thermophilum strain DSM 6725 as Caldicellulosiruptor bescii sp. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 2011–2015. [Google Scholar] [CrossRef]
- Bhandiwad, A.; Guseva, A.; Lynd, L. Metabolic Engineering of Thermoanaerobacterium thermosaccharolyticum for increased n-Butanol production. Adv. Microbiol. 2013, 1, 46–51. [Google Scholar] [CrossRef]
- Su, X.; Han, Y.; Dodd, D.; Moon, Y.H.; Yoshida, S.; Mackie, R.I.; Cann, I.K. Reconstitution of a thermostable xylan-degrading enzyme mixture from the bacterium Caldicellulosiruptor bescii. Appl. Environ. Microbiol. 2013, 79, 1481–1490. [Google Scholar] [CrossRef]
- Han, Y.; Agarwal, V.; Dodd, D.; Kim, J.; Bae, B.; Mackie, R.I.; Nair, S.K.; Cann, I.K. Biochemical and structural insights into xylan utilization by the thermophilic bacterium Caldanaerobius polysaccharolyticus. J. Biol. Chem. 2012, 287, 34946–34960. [Google Scholar] [CrossRef]
- Sizova, M.V.; Izquierdo, J.A.; Panikov, N.S.; Lynd, L.R. Cellulose- and xylan-degrading thermophilic anaerobic bacteria from biocompost. Appl. Environ. Microbiol. 2011, 77, 2282–2291. [Google Scholar]
- Shao, W.; Wiegel, J. Purification and characterization of two thermostable acetyl xylan esterases from Thermoanaerobacterium sp. strain JW/SL-YS485. Appl. Environ. Microbiol. 1995, 61, 729–733. [Google Scholar]
- Sokolova, T.G.; Henstra, A.M.; Sipma, J.; Parshina, S.N.; Stams, A.J.; Lebedinsky, A.V. Diversity and ecophysiological features of thermophilic carboxydotrophic anaerobes. FEMS Microbiol. Ecol. 2009, 68, 131–141. [Google Scholar] [CrossRef]
- Kimble, L.K.; Mandelco, L.; Woese, C.R.; Madigan, M.T. Heliobacterium modesticaldum, sp. nov., a thermophilic heliobacterium of hot springs and volcanic soils. Arch. Microbiol. 1995, 163, 259–267. [Google Scholar] [CrossRef]
- Huber, H.; Hohn, M.J.; Rachel, R.; Fuchs, T.; Wimmer, V.C.; Stetter, K.O. A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. Nature 2002, 417, 63–67. [Google Scholar] [CrossRef]
- Liu, S.V.; Zhou, J.; Zhang, C.; Cole, D.R.; Gajdarziska-Josifovska, M.; Phelps, T.J. Thermophilic Fe(III)-reducing bacteria from the deep subsurface: The evolutionary implications. Science 1997, 277, 1106–1109. [Google Scholar] [CrossRef]
- Roh, Y.; Liu, S.V.; Li, G.; Huang, H.; Phelps, T.J.; Zhou, J. Isolation and characterization of metal-reducing Thermoanaerobacter strains from deep subsurface environments of the Piceance Basin, Colorado. Appl. Environ. Microbiol. 2002, 68, 6013–6020. [Google Scholar] [CrossRef]
- Mesbah, N.M.; Cook, G.M.; Wiegel, J. The halophilic alkalithermophile Natranaerobius thermophilus adapts to multiple environmental extremes using a large repertoire of Na+(K+)/H+ antiporters. Mol. Microbiol. 2009, 74, 270–281. [Google Scholar] [CrossRef]
- Lucas, S.; Han, J.; Lapidus, A.; Cheng, J.F.; Goodwin, L.A.; Pitluck, S.; Peters, L.; Mikhailova, N.; Teshima, H.; Detter, J.C.; et al. Complete genome sequence of the thermophilic, piezophilic, heterotrophic bacterium Marinitoga piezophila KA3. J. Bacteriol. 2012, 194, 5974–5975. [Google Scholar] [CrossRef]
- Alain, K.; Marteinsson, V.T.; Miroshnichenko, M.L.; Bonch-Osmolovskaya, E.A.; Prieur, D.; Birrien, J.L. Marinitoga piezophila sp. nov., a rod-shaped, thermo-piezophilic bacterium isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2002, 52, 1331–1339. [Google Scholar] [CrossRef]
- Slobodkin, A.I.; Tourova, T.P.; Kostrikina, N.A.; Lysenko, A.M.; German, K.E.; Bonch-Osmolovskaya, E.A.; Birkeland, N.K. Tepidimicrobium ferriphilum gen. nov., sp. nov., a novel moderately thermophilic, Fe(III)-reducing bacterium of the order Clostridiales. Int. J. Syst. Evol. Microbiol. 2006, 56, 369–372. [Google Scholar] [CrossRef]
- Prevost, S.; Andre, S.; Remize, F. PCR detection of thermophilic spore-forming bacteria involved in canned food spoilage. Curr. Microbiol. 2010, 61, 525–533. [Google Scholar] [CrossRef]
- Canganella, F.; Wiegel, J. The Potential of Thermophilic Clostridia in Biotechnology. In The Clostridia and Biotechnology; Woods, D.R., Ed.; Butterworths Publishers: Stoneham, MA, USA, 1993; pp. 391–429. [Google Scholar]
- Wiegel, J.; Ljungdahl, L.G. The importance of thermophilic bacteria in biotechnology. Crit. Rev. Biotechnol. 1986, 3, 39–107. [Google Scholar] [CrossRef]
- Kristjansson, J.K. Thermophilic Bacteria; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Gao, Z.M.; Liu, X.; Zhang, X.Y.; Ruan, L.W. Thermophagus xiamenensis gen. nov., sp. nov., a moderately thermophilic and strictly anaerobic bacterium isolated from hot spring sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 109–113. [Google Scholar] [CrossRef]
- Fardeau, M.L.; Barsotti, V.; Cayol, J.L.; Guasco, S.; Michotey, V.; Joseph, M.; Bonin, P.; Ollivier, B. Caldinitratiruptor microaerophilus, gen. nov., sp. nov. isolated from a French hot spring (Chaudes-Aigues, Massif Central): A novel cultivated facultative microaerophilic anaerobic thermophile pertaining to the Symbiobacterium branch within the Firmicutes. Extremophiles 2010, 14, 241–247. [Google Scholar] [CrossRef]
- Romano, I.; Dipasquale, L.; Orlando, P.; Lama, L.; d’Ippolito, G.; Pascual, J.; Gambacorta, A. Thermoanaerobacterium thermostercus sp. nov., a new anaerobic thermophilic hydrogen-producing bacterium from buffalo-dung. Extremophiles 2010, 14, 233–240. [Google Scholar] [CrossRef]
- Arun, A.B.; Chen, W.M.; Lai, W.A.; Chou, J.H.; Shen, F.T.; Rekha, P.D.; Young, C.C. Lutaonella thermophila gen. nov., sp. nov., a moderately thermophilic member of the family Flavobacteriaceae isolated from a coastal hot spring. Int. J. Syst. Evol. Microbiol. 2009, 8, 2069–2073. [Google Scholar]
- Panda, S.K.; Jyoti, V.; Bhadra, B.; Nayak, K.C.; Shivaji, S.; Rainey, F.A.; Das, S.K. Thiomonas bhubaneswarensis sp. nov., a novel obligately mixotrophic, moderately thermophilic, thiosulfate oxidizing bacterium. Int. J. Syst. Evol. Microbiol. 2009, 59, 2171–2175. [Google Scholar] [CrossRef]
- Shiratori, H.; Sasaya, K.; Ohiwa, H.; Ikeno, H.; Ayame, S.; Kataoka, N.; Miya, A.; Beppu, T.; Ueda, K. Clostridium clariflavum sp. nov. and Clostridium caenicola sp. nov., moderately thermophilic, cellulose-/cellobiose-digesting bacteria isolated from methanogenic sludge. Int. J. Syst. Evol. Microbiol. 2009, 59, 1764–1770. [Google Scholar] [CrossRef]
- Westerholm, M.; Roos, S.; Schnürer, A. Tepidanaerobacter acetatoxydans sp. nov., an anaerobic, syntrophic acetate-oxidizing bacterium isolated from two ammonium-enriched mesophilic methanogenic processes. Syst. Appl. Microbiol. 2011, 34, 260–266. [Google Scholar] [CrossRef]
- Nishiyama, T.; Ueki, A.; Kaku, N.; Ueki, K. Clostridium sufflavum sp. nov., isolated from a methanogenic reactor treating cattle waste. Int. J. Syst. Evol. Microbiol. 2009, 59, 981–986. [Google Scholar] [CrossRef]
- Ueki, A.; Abe, K.; Suzuki, D.; Kaku, N.; Watanabe, K.; Ueki, K. Anaerosphaera aminiphila gen. nov., sp. nov., a glutamate-degrading, Gram-positive anaerobic coccus isolated from a methanogenic reactor treating cattle waste. Int. J. Syst. Evol. Microbiol. 2009, 59, 3161–3167. [Google Scholar] [CrossRef]
- Rezgui, R.; Maaroufi, A.; Fardeau, M.L.; Ben, A.; Gam, Z.; Cayol, J.L.; Ben Hamed, S.; Labat, M. Anaerosalibacter bizertensis gen. nov., sp. nov., a halotolerant bacterium isolated from sludge. Int. J. Syst. Evol. Microbiol. 2012, 62, 2469–2474. [Google Scholar] [CrossRef]
- Lee, Y.J.; Romanek, C.S.; Mills, G.L.; Davis, R.C.; Whitman, W.B.; Wiegel, J. Gracilibacter thermotolerans gen. nov., sp. nov., an anaerobic, thermotolerant bacterium from a constructed wetland receiving acid sulfate water. Int. J. Syst. Evol. Microbiol. 2006, 56, 2089–2093. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Canganella, F.; Wiegel, J. Anaerobic Thermophiles. Life 2014, 4, 77-104. https://doi.org/10.3390/life4010077
Canganella F, Wiegel J. Anaerobic Thermophiles. Life. 2014; 4(1):77-104. https://doi.org/10.3390/life4010077
Chicago/Turabian StyleCanganella, Francesco, and Juergen Wiegel. 2014. "Anaerobic Thermophiles" Life 4, no. 1: 77-104. https://doi.org/10.3390/life4010077