
L-Arginine-Dependent Nitric Oxide Production in the Blood of Patients with Type 2 Diabetes: A Pilot, Five-Year Prospective Study
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Subjects and Study Design
2.3. Sample Processing
2.4. Determining L-Arginine Concentration
2.5. Arginase Activity Assay
2.6. Nitrate and Nitrite Analysis
2.7. HbA1c Assay
2.8. Statistical Analysis
3. Results
3.1. Clinical Characteristics of Diabetic Patients at Baseline and Five-Year Follow-Up
3.2. Levels of L-Arginine in Plasma and Red Blood Cells
3.3. Arginase Activity in the Blood of Diabetic Patients
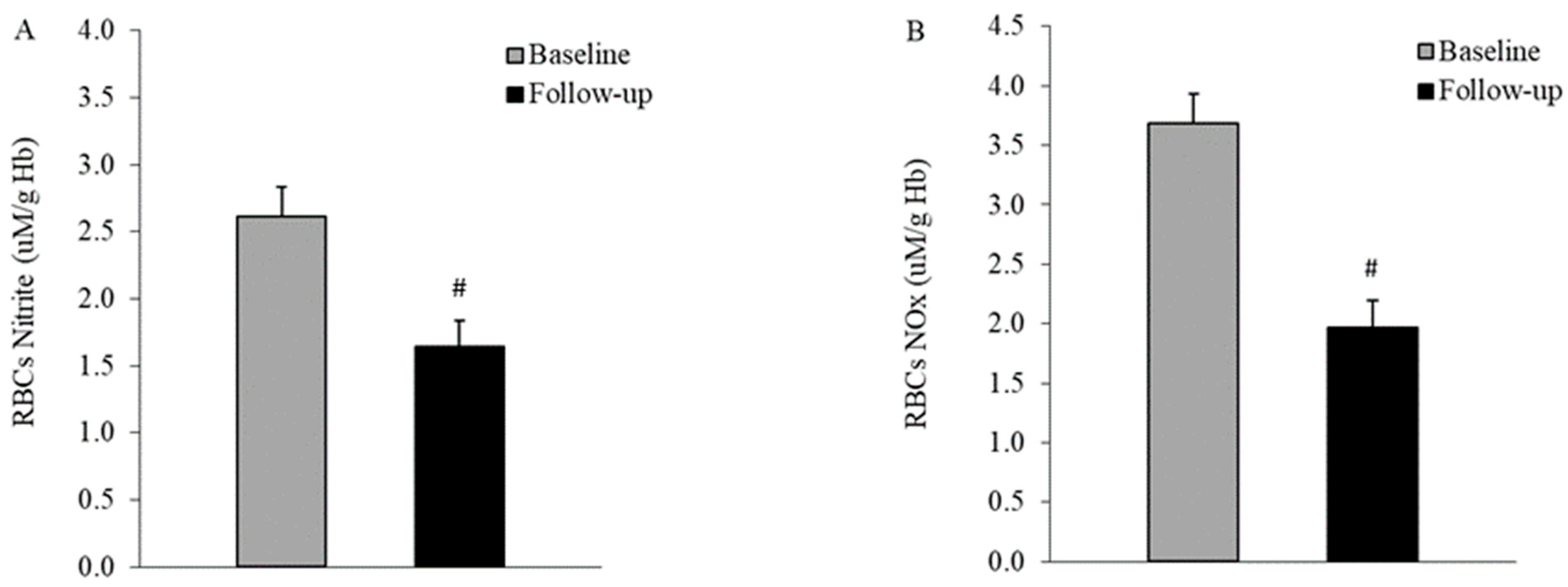
3.4. Nitric Oxide Production in the Blood of Diabetic Patients
3.5. Impact of Statin and ACEI Treatment on Arginase Activity and NO Synthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2021 Diabetes Collaborators. Global, Regional, and National Burden of Diabetes from 1990 to 2021, with Projections of Prevalence to 2050: A Systematic Analysis for the Global Burden of Disease Study 2021. Lancet 2023, 402, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Lagou, V.; Jiang, L.; Ulrich, A.; Zudina, L.; González, K.S.G.; Balkhiyarova, Z.; Faggian, A.; Maina, J.G.; Chen, S.; Todorov, P.V.; et al. Meta-Analysis of Glucose and Insulin-Related Traits Consortium (Magic). Gwas of Random Glucose in 476,326 Individuals Provide Insights into Diabetes Pathophysiology, Complications and Treatment Stratification. Nat. Genet. 2023, 9, 1448–1461. [Google Scholar] [CrossRef]
- Takeda, Y.; Matoba, K.; Sekiguchi, K.; Nagai, Y.; Yokota, T.; Utsunomiya, K.; Nishimura, R. Endothelial Dysfunction in Diabetes. Biomedicines 2020, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S.C.; Mavridis, D.; Nicolucci, A.; Johnson, D.W.; Tonelli, M.; Craig, J.C.; Maggo, J.; Gray, V.; De Berardis, G.; Ruospo, M.; et al. Comparison of Clinical Outcomes and Adverse Events Associated with Glucose-Lowering Drugs in Patients with Type 2 Diabetes: A Meta-Analysis. JAMA 2016, 316, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336 Pt 1, 1–17. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric Oxide Synthases: Structure, Function and Inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cui, H.; Kundu, T.K.; Alzawahra, W.; Zweier, J.L. Nitric Oxide Production from Nitrite Occurs Primarily in Tissues not in the Blood: Critical Role of Xanthine Oxidase and Aldehyde Oxidase. J. Biol. Chem. 2008, 283, 634.1. [Google Scholar] [CrossRef]
- Huang, Z.; Shiva, S.; Kim-Shapiro, D.B.; Patel, R.P.; Ringwood, L.A.; Irby, C.E.; Huang, K.T.; Ho, C.; Hogg, N.; Schechter, A.N.; et al. Enzymatic function of hemoglobin as a nitrite reductase that produces NO under allosteric control. J. Clin. Investig. 2005, 115, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Kleinbongard, P.; Schulz, R.; Rassaf, T.; Lauer, T.; Dejam, A.; Jax, T.; Kumara, I.; Gharini, P.; Kabanova, S.; Özüyaman, B.; et al. Red Blood Cells Express a Functional Endothelial Nitric Oxide Synthase. Blood 2006, 107, 2943–2951. [Google Scholar] [CrossRef]
- Yang, J.; Gonon, A.T.; Sjöquist, P.-O.; Lundberg, J.O.; Pernow, J. Arginase Regulates Red Blood Cell Nitric Oxide Synthase and Export of Cardioprotective Nitric Oxide Bioactivity. Proc. Natl. Acad. Sci. USA 2013, 110, 15049–15054. [Google Scholar] [CrossRef]
- Kim, P.S.; Iyer, R.K.; Lu, K.V.; Yu, H.; Karimi, A.; Kern, R.M.; Tai, D.K.; Cederbaum, S.D.; Grody, W.W. Expression of the liver form of arginase in erythrocytes. Mol. Genet. Metab. 2002, 76, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, S.R.; Lara, A.; Zhang, R.; Park, Y.M.; DeFronzo, R.A. Insulin Reduces Plasma Arginase Activity in Type 2 Diabetic Patients. Diabetes Care 2008, 31, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Shatanawi, A.; Momani, M.S.; Al-Aqtash, R.; Hamdan, M.H.; Gharaibeh, M.N. L-Citrulline Supplementation Increases Plasma Nitric Oxide Levels and Reduces Arginase Activity in Patients with Type 2 Diabetes. Front. Pharmacol. 2020, 11, 584669. [Google Scholar] [CrossRef] [PubMed]
- Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An Old Enzyme with New Tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Bizjak, D.A.; Brinkmann, C.; Bloch, W.; Grau, M. Increase in Red Blood Cell-Nitric Oxide Synthase Dependent Nitric Oxide Production during Red Blood Cell Aging in Health and Disease: A Study on Age Dependent Changes of Rheologic and Enzymatic Properties in Red Blood Cells. PLoS ONE 2015, 10, e0125206. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Zamora, S.; Méndez-Rodríguez, M.L.; Olguín-Martínez, M.; Sánchez-Sevilla, L.; Quintana-Quintana, M.; García-García, N.; Hernández-Muñoz, R. Increased Erythrocytes By-Products of Arginine Catabolism Are Associated with Hyperglycemia and Could Be Involved in the Pathogenesis of Type 2 Diabetes Mellitus. PLoS ONE 2013, 8, e66823. [Google Scholar] [CrossRef] [PubMed]
- Savu, O.; Iosif, L.; Bradescu, O.M.; Serafinceanu, C.; Papacocea, R.; Stoian, I. L-Arginine Catabolism is Driven Mainly towards Nitric Oxide Synthesis in the Erythrocytes of Patients with Type 2 Diabetes at First Clinical Onset. Ann. Clin. Biochem. Int. J. Biochem. Lab. Med. 2014, 52 Pt 1, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Eligini, S.; Porro, B.; Werba, J.P.; Capra, N.; Genovese, S.; Greco, A.; Cavalca, V.; Banfi, C. Oxidative Stress and Arginine/Nitric Oxide Pathway in Red Blood Cells Derived from Patients with Prediabetes. Biomedicines 2022, 10, 1407. [Google Scholar] [CrossRef]
- Piatti, P.; Monti, L.D.; Valsecchi, G.; Magni, F.; Setola, E.; Marchesi, F.; Galli-Kienle, M.; Pozza, G.; Alberti, K.G.M. Long-Term Oral l-Arginine Administration Improves Peripheral and Hepatic Insulin Sensitivity in Type 2 Diabetic Patients. Diabetes Care 2001, 24, 875–880. [Google Scholar] [CrossRef]
- Schulman, S.P.; Becker, L.C.; Kass, D.A.; Champion, H.C.; Terrin, M.L.; Forman, S.; Ernst, K.V.; Kelemen, M.D.; Townsend, S.N.; Capriotti, A.; et al. L-Arginine Therapy in Acute Myocardial Infarction: The Vascular Interaction with Age in Myocardial Infarction (Vintage Mi) Randomized Clinical Trial. JAMA 2006, 295, 58–64. [Google Scholar] [CrossRef]
- McMahon, T.J.; Moon, R.E.; Luschinger, B.P.; Carraway, M.S.; Stone, A.E.; Stolp, B.W.; Gow, A.J.; Pawloski, J.R.; Watke, P.; Singel, D.J.; et al. Nitric oxide in the human respiratory cycle. Nat. Med. 2002, 8, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Pawloski, J.R.; Hess, D.T.; Stamler, J.S. Export by Red Blood Cells of Nitric Oxide Bioactivity. Nature 2001, 409, 622–626. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, X.; Mahdi, A.; Zhou, Z.; Tratsiakovich, Y.; Jiao, T.; Kiss, A.; Kövamees, O.; Alvarsson, M.; Catrina, S.-B.; et al. Red Blood Cells in Type 2 Diabetes Impair Cardiac Post-Ischemic Recovery Through an Arginase-Dependent Modulation of Nitric Oxide Synthase and Reactive Oxygen Species. JACC Basic Transl. Sci. 2018, 3, 450–463. [Google Scholar] [CrossRef] [PubMed]
- Merx, M.W.; Gorressen, S.; Sandt, A.M.; Cortese-Krott, M.M.; Ohlig, J.; Stern, M.; Rassaf, T.; Gödecke, A.; Gladwin, M.T.; Kelm, M. Depletion of circulating blood NOS3 increases severity of myocardial infarction and left ventricular dysfunction. Basic Res. Cardiol. 2013, 109, 398. [Google Scholar] [CrossRef] [PubMed]
- James, P.E.; Lang, D.; Tufnell-Barret, T.; Milsom, A.B.; Frenneaux, M.P. Vasorelaxation by Red Blood Cells and Impairment in Diabetes: Reduced Nitric Oxide and Oxygen Delivery by Glycated Hemoglobin. Circ. Res. 2004, 94, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Ferlito, S.; Gallina, M. Nitrite Plasma Levels in Type 1 and 2 Diabetics with and without Complications. Minerva Endocrinol. 2000, 24, 117–121. [Google Scholar]
- Vanizor, B.; Örem, A.; Karahan, S.; Kiran, E.; Erem, C.; Aliyazicioğlu, R.; Uydu, H.A. Decreased Nitric Oxide End-Products and Its Relationship with High Density Lipoprotein and Oxidative Stress in People with Type 2 Diabetes without Complications. Diabetes Res. Clin. Pract. 2001, 54, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Asl, S.Z.; Ghasemi, A.; Azizi, F. Serum Nitric Oxide Metabolites in Subjects with Metabolic Syndrome. Clin. Biochem. 2008, 41, 1342–1347. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A Rapid, Simple Spectrophotometric Method for Simultaneous Detection of Nitrate and Nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Zinellu, A.; Sotgia, S.; Usai, M.F.; Pintus, G.; Deiana, L.; Carru, C. Improved Method for Plasma ADMA, SDMA, and Arginine Quantification by Field-Amplified Sample Injection Capillary Electrophoresis UV Detection. Anal. Bioanal. Chem. 2010, 399, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, K.; Karai, I.; Nishikawa, Y.; Horiguchi, S. Erythrocyte Arginase Activity as an Indicator of Lead Exposure. Occup. Environ. Med. 1983, 40, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.; Hendgen-Cotta, U.B.; Brouzos, P.; Drexhage, C.; Rassaf, T.; Lauer, T.; Dejam, A.; Kelm, M.; Kleinbongard, P. Recent methodological advances in the analysis of nitrite in the human circulation: Nitrite as a biochemical parameter of the l-arginine/NO pathway. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2007, 851, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Groche, D.; Hoeno, W.; Hoss, G.; Vogt, B.; Herrmann, Z.; Witzigmann, A. Standardization of Two Immunological Hba1c Routine Assays According to the New Ifcc Reference Method. Clin. Lab. 2003, 49, 657–661. [Google Scholar] [PubMed]
- Kosenko, E.; Tikhonova, L.; Suslikov, A.; Kaminsky, Y. Impacts of Lisinopril and Lisinopril Plus Simvastatin on Erythrocyte and Plasma Arginase, Nitrite, and Nitrate in Hypertensive Patients. J. Clin. Pharmacol. 2012, 52, 102–109. [Google Scholar] [CrossRef]
- Gajecki, D.; Gawryś, J.; Wiśniewski, J.; Fortuna, P.; Szahidewicz-Krupska, E.; Doroszko, A. A Cross-Talk between the Erythrocyte L-Arginine/Adma/Nitric Oxide Metabolic Pathway and the Endothelial Function in Subjects with Type 2 Diabetes Mellitus. Nutrients 2021, 13, 2306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Mahdi, A.; Tratsiakovich, Y.; Zahorán, S.; Kövamees, O.; Nordin, F.; Uribe Gonzalez, A.E.; Alvarsson, M.; Östenson, C.-G.; Andersson, D.C.; et al. Erythrocytes from Patients with Type 2 diabetes Induce Endothelial dysfunction Via Arginase I. J. Am. Coll. Cardiol. 2018, 72, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Durante, W.; Johnson, F.K.; A Johnson, R. Arginase: A Critical Regulator of Nitric Oxide Synthesis and Vascular Function. Clin. Exp. Pharmacol. Physiol. 2007, 34, 906–911. [Google Scholar] [CrossRef]
- Qazi, M.A.; Rizzatti, F.; Piknova, B.; Sibmooh, N.; Stroncek, D.F.; Schechter, A.N. Effect of Storage Levels of Nitric Oxide Derivatives in Blood Components. F1000Res 2012, 1, 35. [Google Scholar] [CrossRef]
- Maejima, K.; Nakano, S.; Himeno, M.; Tsuda, S.; Makiishi, H.; Ito, T.; Nakagawa, A.; Kigoshi, T.; Ishibashi, T.; Nishio, M.; et al. Increased Basal Levels of Plasma Nitric Oxide in Type 2 Diabetic Subjects. Relationship to Microvascular Complications. J. Diabetes Complicat. 2001, 15, 135–143. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type 2 Diabetes Mellitus | t-Test (Paired) | ||
|---|---|---|---|
| Baseline | Follow-Up (+5 Years) | ||
| Age (years) | 50.3 ± 3.0 | 56.0 ± 3.1 | |
| Gender (n, women/men) | 5/5 | 5/5 | |
| BMI (kg/m2) | 26.9 ± 1.0 | 27.3 ± 0.9 | 0.53 |
| OAD (n) | 5 | 5 | N/A |
| SBP (mmHg) | 140.1 ± 5.0 | 142.0 ± 4.1 | 0.81 |
| DBP (mmHg) | 87.1 ± 2.1 | 83.1 ± 1.7 | 0.13 |
| HbA1c (%) | 8.4 ± 0.6 | 8.6 ± 0.5 | 0.80 |
| Hb (g/dL) | 14.4 ± 0.30 | 14.5 ± 0.50 | 0.88 |
| RBCs (×105/µL) | 4.95 ± 0.08 | 4.9 ± 0.12 | 0.83 |
| WBCs (×103/µL) | 7.35 ± 0.3 | 7.3 ± 0.3 | 0.35 |
| PLT (×103/µL) | 230.1 ± 14.9 | 221.3 ± 15.2 | 0.68 |
| Cholesterol (mg/dL) | 213.3 ± 12.0 | 212.5 ± 21.1 | 0.97 |
| HDL (mg/dL) | 38.5 ± 3.3 | 46.6 ± 2.9 | 0.09 |
| TG (mg/dL) | 188.6 ± 26.0 | 175.5 ± 19.8 | 0.69 |
| Creatinine (mg/dL) | 0.81 ± 0.1 | 0.73 ± 0.1 | 0.39 |
| ALAT (UI/L) | 28.2 ± 6.7 | 24.7 ± 3.1 | 0.59 |
| ASAT (UI/L) | 51.6 ± 11.7 | 42.4 ± 5.2 | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoian, I.; Iosif, L.; Gilca, M.; Vlad, A.; Tivig, I.; Bradescu, O.M.; Savu, O. L-Arginine-Dependent Nitric Oxide Production in the Blood of Patients with Type 2 Diabetes: A Pilot, Five-Year Prospective Study. Life 2024, 14, 556. https://doi.org/10.3390/life14050556
Stoian I, Iosif L, Gilca M, Vlad A, Tivig I, Bradescu OM, Savu O. L-Arginine-Dependent Nitric Oxide Production in the Blood of Patients with Type 2 Diabetes: A Pilot, Five-Year Prospective Study. Life. 2024; 14(5):556. https://doi.org/10.3390/life14050556
Chicago/Turabian StyleStoian, Irina, Liviu Iosif, Marilena Gilca, Adelina Vlad, Ioan Tivig, Ovidiu Marius Bradescu, and Octavian Savu. 2024. "L-Arginine-Dependent Nitric Oxide Production in the Blood of Patients with Type 2 Diabetes: A Pilot, Five-Year Prospective Study" Life 14, no. 5: 556. https://doi.org/10.3390/life14050556