
Anxiolytic-like Activity, Antioxidant Properties, and Facilitatory Effects on the Short-Term Memory Retention of Molsidomine in Rats
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
- Group I (Control): saline solution, 0.3 mL/100 g of body weight;
- Group II (Mg): 200 mg/kg of body weight of magnesium chloride;
- Group III (MSD): 1 mg/kg of body weight of molsidomine;
- Group IV (V-pyrro/NO): 5 mg/kg of body weightof V-pyrro/NO.
2.3. The Spontaneous Behavior Evaluation
2.4. The Short-Term Memory Evaluation
2.5. Laboratory Investigations
2.6. Data Analysis
3. Results
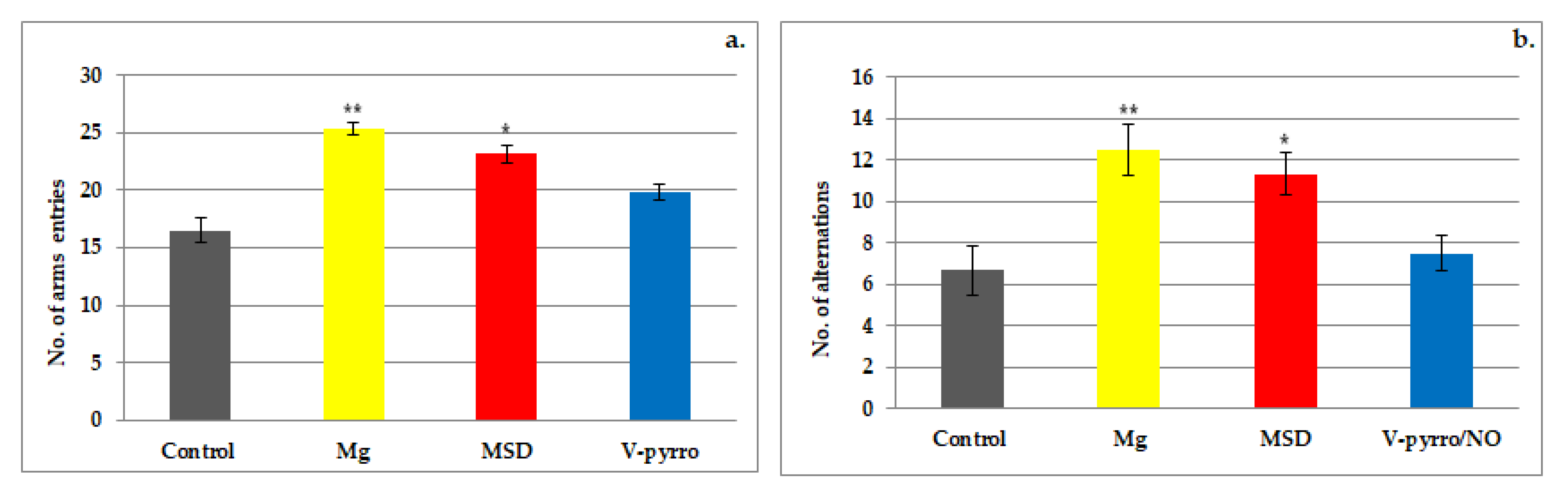
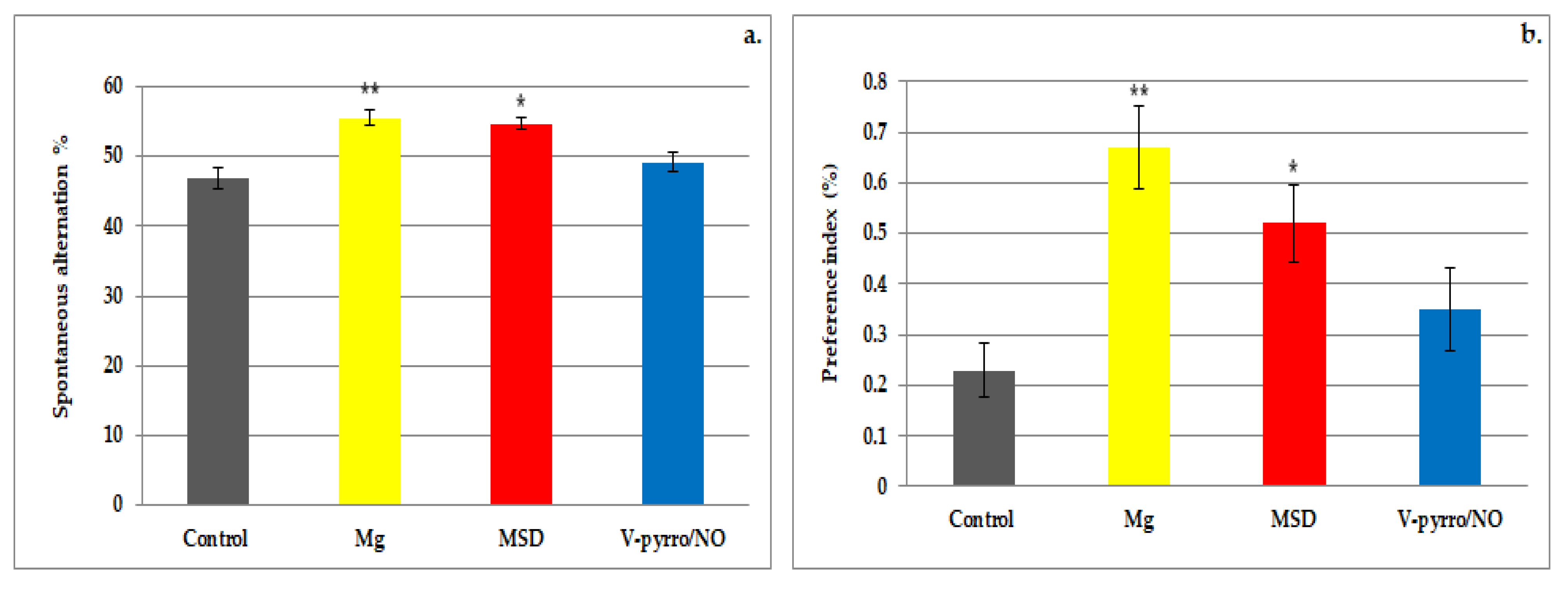
3.1. The Spontaneous Behavior Evaluation
3.2. The Short-Term Memory Evaluation
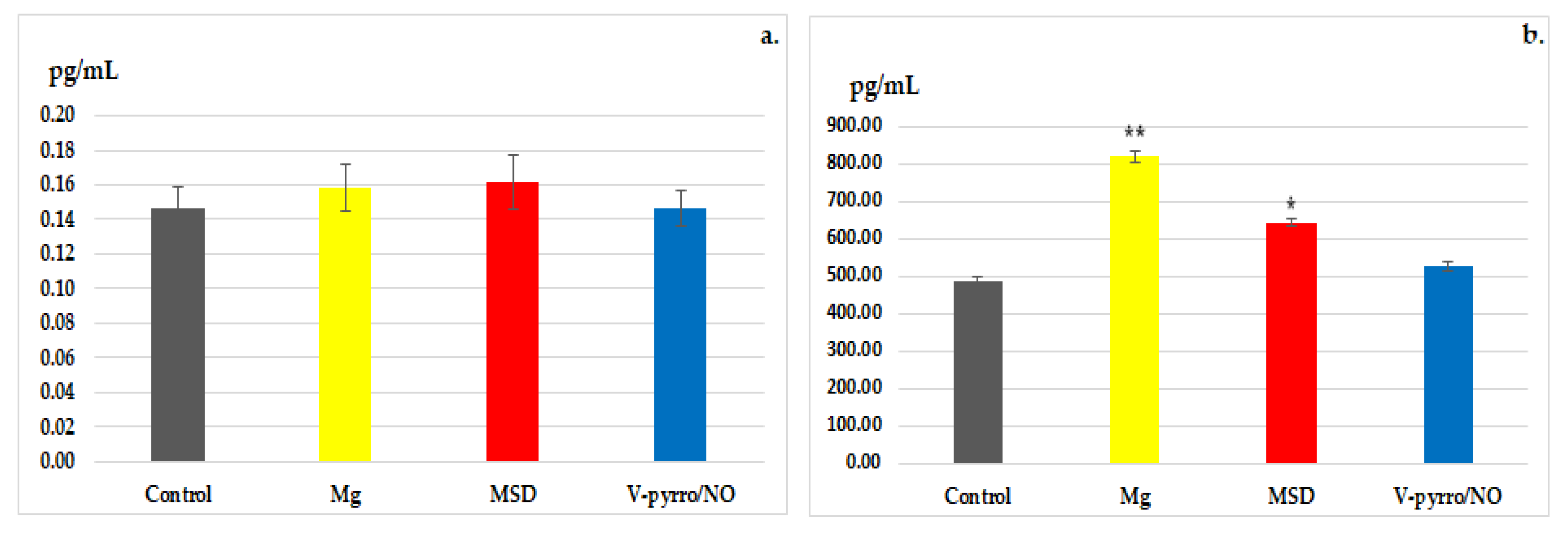
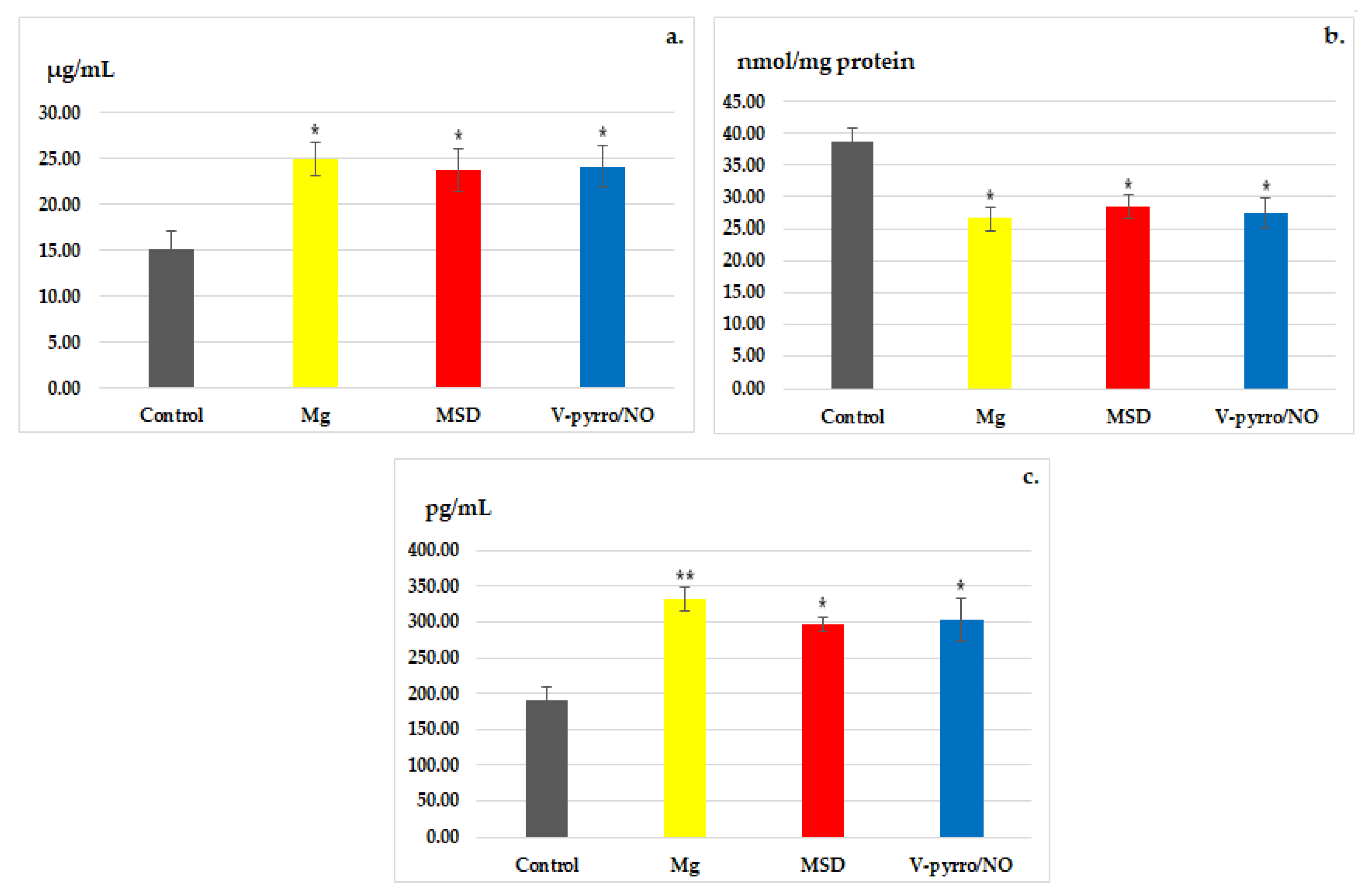
3.3. Laboratory Investigations
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iova, O.M.; Marin, G.E.; Lazar, I.; Stanescu, I.; Dogaru, G.; Nicula, C.A.; Bulboacă, A.E. Nitric oxide/nitric oxide synthase system in the pathogenesis of neurodegenerative disorders-an overview. Antioxidants 2023, 12, 753. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.B.; Punihaole, D.; Levine, T.B. Characterization of the role of nitric oxide and its clinical applications. Cardiology 2012, 122, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.-L.; Ritter, J.K. Role of nitric oxide in the cardiovascular and renal systems. Int. J. Mol. Sci. 2018, 19, 2605. [Google Scholar] [CrossRef] [PubMed]
- Barreto, A.S.; Macedo, F.N.; Fontes, M.T.; Santana-Filho, W.J. Chapter 3—Nitric oxide as a vascular modulator to resistance training. In Endothelial Signaling in Vascular Dysfunction and Disease; Chatterjee, S., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 29–35. [Google Scholar] [CrossRef]
- Bayarri, M.A.; Milara, J.; Estornut, C.; Cortijo, J. Nitric oxide system and bronchial epithelium: More than a barrier. Front. Physiol. 2021, 12, 687381. [Google Scholar] [CrossRef]
- da Silva, G.M.; da Silva, M.C.; Gomes Nascimento, D.V.; Lima Silva, E.M.; Gouvêa, F.F.F.; de França Lopes, L.G.; Araújo, A.V.; Ferraz Pereira, K.N.; de Queiroz, T.M. Nitric oxide as a central molecule in hypertension: Focus on the vasorelaxant activity of new nitric oxide donors. Biology 2021, 10, 1041. [Google Scholar] [CrossRef]
- Jiang, W.; Dong, W.; Li, M.; Guo, Z.; Wang, Q.; Liu, Y.; Bi, Y.; Zhou, H.; Wang, Y. Nitric oxide induces immunogenic cell death and potentiates cancer immunotherapy. ACS Nano 2022, 16, 3881–3894. [Google Scholar] [CrossRef]
- Hays, E.; Bonavida, B. Nitric oxide-mediated enhancement and reversal of resistance of anticancer therapies. Antioxidants 2019, 8, 407. [Google Scholar] [CrossRef]
- Thomas, D.D. Breathing new life into nitric oxide signaling: A brief overview of the interplay between oxygen and nitric oxide. Redox Biol. 2015, 5, 225–233. [Google Scholar] [CrossRef]
- Oliveira, C.; Benfeito, S.; Fernandes, C.; Cagide, F.; Silva, T.; Borges, F. NO and HNO donors, nitrones, and nitroxides: Past, present, and future. Med. Res. Rev. 2018, 38, 1159–1187. [Google Scholar] [CrossRef]
- Maccallini, C.; Amoroso, R. Neuronal nitric oxide synthase and post-translational modifications in the development of central nervous system diseases: Implications and regulation. Molecules 2023, 28, 6691. [Google Scholar] [CrossRef]
- Amal, H.; Steinert, J.R. Editorial: Nitric oxide signaling from synapse to disease. Free Radic. Biol. Med. 2023, 196, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Sagi, Y.; Heiman, M.; Peterson, J.D.; Musatov, S.; Scarduzio, M.; Logan, S.M.; Kaplitt, M.G.; Surmeier, D.J.; Heintz, N.; Greengard, P. Nitric oxide regulates synaptic transmission between spiny projection neurons. Proc. Natl. Acad. Sci. USA 2014, 111, 17636–17641. [Google Scholar] [CrossRef]
- Koriyama, Y.; Furukawa, A. S-nitrosylation regulates cell survival and death in the central nervous system. Neurochem. Res. 2018, 43, 50–58. [Google Scholar] [CrossRef]
- Janaszak-Jasiecka, A.; Płoska, A.; Wierońska, J.M.; Dobrucki, L.W.; Kalinowski, L. Endothelial dysfunction due to eNOS uncoupling: Molecular mechanisms as potential therapeutic targets. Cell Mol. Biol. Lett. 2023, 28, 21. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.M.; Townsend, J.R.; Pinzone, A.G.; Hoffman, J.R. Supplementation with nitric oxide precursors for strength performance: A review of the current literature. Nutrients 2023, 15, 660. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Laranjinha, J. Nitric oxide production from nitrite plus ascorbate during ischemia upon hippocampal glutamate NMDA receptor stimulation. BioChem 2023, 3, 78–90. [Google Scholar] [CrossRef]
- Olufunmilayo, E.O.; Gerke-Duncan, M.B.; Holsinger, R.M.D. Oxidative stress and antioxidants in neurodegenerative disorders. Antioxidants 2023, 12, 517. [Google Scholar] [CrossRef]
- Ghatak, S.; Nakamura, T.; Lipton, S.A. Aberrant protein S-nitrosylation contributes to hyperexcitability-induced synaptic damage in Alzheimer's disease: Mechanistic insights and potential therapies. Front. Neural Circuits 2023, 17, 1099467. [Google Scholar] [CrossRef]
- Behl, T.; Rana, T.; Sehgal, A.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Meraya, A.M.; Bhatia, S.; Sachdeva, M. Phytochemicals targeting nitric oxide signaling in neurodegenerative diseases. Nitric Oxide 2023, 130, 1–11. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of neuroinflammation in neurodegeneration development. Signal Transduct. Target. Ther. 2023, 8, 267. [Google Scholar] [CrossRef]
- Zhu, L.J.; Li, F.; Zhu, D.Y. nNOS and neurological, neuropsychiatric disorders: A 20-year story. Neurosci. Bull. 2023, 39, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Sah, A.N.; Bawari, S.; Nabavi, S.F.; Dehpour, A.R.; Shirooie, S.; Braidy, N.; Fiebich, B.L.; Vacca, R.A.; Nabavi, S.M. Role of nitric oxide in neurodegeneration: Function, regulation, and inhibition. Curr. Neuropharmacol. 2021, 19, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, V.B.; Bhatnagar, D.; Rubin, H.; Reynolds, M.M. Chapter 6—Nitric oxide donors in nerve regeneration. In Nitric Oxide Donors. Novel Biomedical Applications and Perspectives; Seabra, A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 141–168. [Google Scholar]
- Locascio, A.; Annona, G.; Caccavale, F.; D’Aniello, S.; Agnisola, C.; Palumbo, A. Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates. Int. J. Mol. Sci. 2023, 24, 11182. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Nacharaju, P.; Friedman, A.; Friedman, J.M. Nitric oxide generating/releasing materials. Future Sci. OA. 2015, 1, FSO54. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yoon, B.; Dey, A.; Nguyen, V.Q.; Park, J.H. Recent progress in nitric oxide-generating nanomedicine for cancer therapy. J. Control. Release 2022, 352, 179–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, A.; Yang, Z.; Huang, W. Advanced nitric oxide generating nanomedicine for therapeutic applications. ACS Nano. 2023, 17, 8935–8965. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Fu, J.; Zhang, Y. Nitric oxide donor-based cancer therapy: Advances and prospects. J. Med. Chem. 2017, 60, 7617–7635. [Google Scholar] [CrossRef]
- Li, C.Y.; Anuraga, G.; Chang, C.P.; Weng, T.Y.; Hsu, H.P.; Ta, H.D.K.; Su, P.F.; Chiu, P.H.; Yang, S.J.; Chen, F.W.; et al. Repurposing nitric oxide donating drugs in cancer therapy through immune modulation. J. Exp. Clin. Cancer Res. 2023, 42, 22. [Google Scholar] [CrossRef]
- Roth, L.; Van der Donckt, C.; Emini Veseli, B.; Van Dam, D.; De Deyn, P.P.; Martinet, W.; Herman, A.G.; De Meyer, G.R.Y. Nitric oxide donor molsidomine favors features of atherosclerotic plaque stability and reduces myocardial infarction in mice. Vascul. Pharmacol. 2019, 118–119, 106561. [Google Scholar] [CrossRef]
- Boerhout, C.K.M.; Feenstra, R.G.T.; van de Hoef, T.P.; Piek, J.J.; Beijk, M.A.M. Pharmacotherapy in patients with vasomotor disorders. Int. J. Cardiol. Heart Vasc. 2023, 48, 101267. [Google Scholar] [CrossRef]
- Katsanou, L.; Fragkiadaki, E.; Kampouris, S.; Konstanta, A.; Vontzou, A.; Pitsikas, N. The nitric oxide (NO) donor molsidomine counteract social withdrawal and cognition deficits induced by blockade of the NMDA receptor in the rat. Int. J. Mol. Sci. 2023, 24, 6866. [Google Scholar] [CrossRef] [PubMed]
- Vartzoka, F.; Ozenoglu, E.; Pitsikas, N. The nitric oxide (NO) donor molsidomine attenuates memory impairments induced by the D1/D2 dopaminergic receptor agonist apomorphine in the rat. Molecules 2023, 28, 6861. [Google Scholar] [CrossRef] [PubMed]
- Pitsikas, N.; Rigamonti, A.E.; Cella, S.G.; Muller, E.E. Effects of the nitric oxide donor molsidomine on different memory components as assessed in the object-recognition task in the rat. Psychopharmacology 2002, 162, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Pitsikas, N.; Sakellaridis, N. Memantine and recognition memory: Possible facilitation of its behavioral effects by the nitric oxide (NO) donor molsidomine. Eur. J. Pharmacol. 2007, 571, 174–179. [Google Scholar] [CrossRef]
- Kalouda, T.; Pitsikas, N. The nitric oxide donor molsidomine induces anxiolytic-like behaviour in two different rat models of anxiety. Pharmacol. Biochem. Behav. 2015, 138, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, J.; Saavedra, J.E.; Keefer, L.K.; Waalkes, M.P. The nitric oxide donor, V-PYRRO/NO, protects against acetaminophen-induced nephrotoxicity in mice. Toxicology 2003, 189, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Zuckerbraun, B.S.; Billiar, T.R.; Otterbein, S.L.; Kim, P.K.; Liu, F.; Choi, A.M.; Bach, F.H.; Otterbein, L.E. Carbon monoxide protects against liver failure through nitric oxide-induced heme oxygenase 1. J. Exp. Med. 2003, 198, 1707–1716. [Google Scholar] [CrossRef]
- Kus, E.; Jasiński, K.; Skórka, T.; Cichon, I.-C.; Chlopicki, S. Short-term treatment with hepatoselective NO donor V-PYRRO/NO improves blood flow in hepatic microcirculation in liver steatosis in mice. Pharmacol. Rep. 2018, 70, 463–469. [Google Scholar] [CrossRef]
- European Union. DIRECTIVE 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; European Union: Brussels, Belgium, 2010; Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 15 March 2018).
- Poleszak, E.; Szewczyk, B.; Kedzierska, E.; Wlaź, P.; Pilc, A.; Nowak, G. Antidepressant- and anxiolytic-like activity of magnesium in mice. Pharmacol. Biochem. Behav. 2004, 78, 7–12. [Google Scholar] [CrossRef]
- Slutsky, I.; Abumaria, N.; Wu, L.J.; Huang, C.; Zhang, L.; Li, B.; Zhao, X.; Govindarajan, A.; Zhao, M.G.; Zhuo, M.; et al. Enhancement of learning and memory by elevating brain magnesium. Neuron 2010, 65, 165–177. [Google Scholar] [CrossRef]
- Klein, C.J.M.I.; Budiman, T.; Homberg, J.R.; Verma, D.; Keijer, J.; van Schothorst, E.M. Measuring locomotor activity and behavioral aspects of rodents living in the home-cage. Front. Behav. Neurosci. 2022, 16, 877323. [Google Scholar] [CrossRef] [PubMed]
- Ilchibaeva, T.; Tsybko, A.; Lipnitskaya, M.; Eremin, D.; Milutinovich, K.; Naumenko, V.; Popova, N. Brain-derived neurotrophic factor (BDNF) in mechanisms of autistic-like behavior in BTBR mice: Crosstalk with the dopaminergic brain system. Biomedicines 2023, 11, 1482. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Taylor, C.; Williamson, M.; Newton, S.S.; Qin, L. Diminished activity-dependent BDNF signaling differentially causes autism-like behavioral deficits in male and female mice. Front. Psychiatry 2023, 14, 1182472. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, H.; Lee, Y.B.; Lee, B.; Lee, D. A quantitative analysis of spontaneous alternation behaviors on a Y-maze reveals adverse effects of acute social isolation on spatial working memory. Sci. Rep. 2023, 13, 14722. [Google Scholar] [CrossRef] [PubMed]
- Navarro, D.; Gasparyan, A.; Martí Martínez, S.; Díaz Marín, C.; Navarrete, F.; García Gutiérrez, M.S.; Manzanares, J. Methods to identify cognitive alterations from animals to humans: A translational approach. Int. J. Mol. Sci. 2023, 24, 7653. [Google Scholar] [CrossRef] [PubMed]
- Melbiarta, P.P.; Kalanjati, V.P.; Herawati, L.; Salim, Y.; Othman, Z. Analysis of spatial working memory using the Y-maze on rodents treated with high-calorie diet and moderate-intensity exercise. Folia Medica Indones. 2023, 59, 40–45. [Google Scholar] [CrossRef]
- Schiavo, A.; Martins, L.A.; Wearick-Silva, L.E.; Orso, R.; Xavier, L.L.; Mestriner, R.G. Can anxiety-like behavior and spatial memory predict the extremes of skilled walking performance in mice? An exploratory, preliminary study. Front. Behav. Neurosci. 2023, 17, 1059029. [Google Scholar] [CrossRef]
- Dronse, J.; Ohndorf, A.; Richter, N.; Bischof, G.N.; Fassbender, R.; Behfar, Q.; Gramespacher, H.; Dillen, K.; Jacobs, H.I.L.; Kukolja, J.; et al. Serum cortisol is negatively related to hippocampal volume, brain structure, and memory performance in healthy aging and Alzheimer's disease. Front. Aging Neurosci. 2023, 15, 1154112. [Google Scholar] [CrossRef]
- Tripathi, A.; Nasrallah, H.A.; Pillai, A. Pimavanserin treatment increases plasma brain-derived neurotrophic factor levels in rats. Front. Neurosci. 2023, 17, 1237726. [Google Scholar] [CrossRef]
- Peskin, A.V.; Winterbourn, C.C. Assay of superoxide dismutase activity in a plate assay using WST-1. Free Radic. Bio. Med. 2017, 103, 188–191. [Google Scholar] [CrossRef]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of oxidative stress in biological samples using the thiobarbituric acid reactive substances assay. J. Vis. Exp. 2020, 159, e61122. [Google Scholar] [CrossRef]
- Ola, O.S.; Odunola, O.A. Eulophia gracilis pseudobulb extract mitigates cyclophosphamide-induced genotoxicity and oxidative stress on murine hepatic tissue. J. Umm Al-Qura Univ. Appll Sci. 2023, 9, 426–435. [Google Scholar] [CrossRef]
- Samdanci, E.T.; Huz, M.; Ozhan, O.; Tanbek, K.; Pamukcu, E.; Akatli, A.N.; Parlakpinar, H. Cytoprotective effects of molsidomine against methotrexate-induced hepatotoxicity: An experimental rat study. Drug Des. Devel Ther. 2018, 13, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Karakoc, H.T.; Altintas, R.; Parlakpinar, H.; Polat, A.; Samdanci, E.; Sagir, M.; Duran, Z.R. Protective Effects of Molsidomine Against Cisplatin-Induced Nephrotoxicity. Adv. Clin. Exp. Med. 2015, 24, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.R.; Megson, I.L. Recent developments in nitric oxide donor drugs. Br. J. Pharmacol. 2007, 151, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Kostov, K.; Halacheva, L. Role of magnesium deficiency in promoting atherosclerosis, endothelial dysfunction, and arterial stiffening as risk factors for hypertension. Int. J. Mol. Sci. 2018, 19, 1724. [Google Scholar] [CrossRef] [PubMed]
- Houston, M. The Role of Magnesium in Hypertension and Cardiovascular Disease. J. Clin. Hypertens. 2011, 13, 843–847. [Google Scholar] [CrossRef]
- Joris, P.J.; Plat, J.; Bakker, S.J.L.; Mensink, R.P. Effects of long-term magnesium supplementation on endothelial function and cardiometabolic risk markers: A randomized controlled trial in overweight/obese adults. Sci. Rep. 2017, 7, 106. [Google Scholar] [CrossRef]
- Cunha, A.R.; Umbelino, B.; Correia, M.L.; Neves, M.F. Magnesium and vascular changes in hypertension. Int. J. Hypertens. 2012, 2012, 754250. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Locatelli, L.; Fedele, G.; Cazzaniga, A.; Mazur, A. Magnesium and the brain: A focus on neuroinflammation and neurodegeneration. Int. J. Mol. Sci. 2022, 24, 223. [Google Scholar] [CrossRef]
- Mathew, A.A.; Panonnummal, R. Magnesium’—The master cation-as a drug—Possibilities and evidences. Biometals 2021, 34, 955–986. [Google Scholar] [CrossRef] [PubMed]
- de Baaij, J.H.F.; Hoenderop, J.G.J.; Bindels, R.J.M. Magnesium in man: Implications for health and disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Shen, Y.; Chopp, M.; Zacharek, A.; Venkat, P.; Chen, Z.; Li, W.; Qian, Y.; Landschoot-Ward, J.; Chen, J. Deficiency of endothelial nitric oxide synthase (enos) exacerbates brain damage and cognitive deficit in a mouse model of vascular dementia. Aging Dis. 2021, 12, 732–746. [Google Scholar] [CrossRef] [PubMed]
- Picón-Pagès, P.; Garcia-Buendia, J.; Muñoz, F.J. Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys. Acta. Mol. Basis Dis. 2019, 1865, 1949–1967. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Guan, P.P.; Zhu, D.; Liang, Y.Y.; Wang, T.; Wang, Z.Y.; Wang, P. Magnesium ions inhibit the expression of tumor necrosis factor α and the activity of γ-secretase in a β-amyloid protein-dependent mechanism in APP/PS1 transgenic mice. Front. Mol. Neurosci. 2018, 11, 172. [Google Scholar] [CrossRef]
- Veronese, N.; Pizzol, D.; Smith, L.; Dominguez, L.J.; Barbagallo, M. Effect of magnesium supplementation on inflammatory parameters: A meta-analysis of randomized controlled trials. Nutrients 2022, 14, 679. [Google Scholar] [CrossRef] [PubMed]
- Himanshu, D.; Sarkar, D.N. A review of behavioral tests to evaluate different types of anxiety and anti-anxiety effects. Clin. Psychopharmacol. Neurosci. 2020, 18, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.J.; Castagne, V.; Moser, P.C.; Mittelstadt, S.W. Comparison of methods for the assessment of locomotor activity in rodent safety pharmacology studies. J. Pharmacol. Toxicol. Method. 2011, 64, 74–80. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-maze for assessment of spatial working and reference memory in mice. Methods Mol. Biol. 2019, 1916, 105–111. [Google Scholar] [CrossRef]
- Guo, W.; Nagappan, G.; Lu, B. Differential effects of transient and sustained activation of BDNF-TrkB signaling. Dev. Neurobiol. 2018, 78, 647–659. [Google Scholar] [CrossRef]
- Casagrande, B.P.; Ribeiro, A.M.; Pisani, L.P.; Estadella, D. Hippocampal BDNF mediated anxiety-like behaviours induced by obesogenic diet withdrawal. Behav. Brain Res. 2023, 436, 114077. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The role of BDNF on neural plasticity in depression. Front. Cell Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.; Hill, J.; Calcaterra, N. Functional role of BDNF production from unique promoters in aggression and serotonin signaling. Neuropsychopharmacol 2016, 41, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Martinowich, K.; Manji, H.; Lu, B. New insights into BDNF function in depression and anxiety. Nat. Neurosci. 2007, 10, 1089–1093. [Google Scholar] [CrossRef]
- Martinowich, K.; Lu, B. Interaction between BDNF and serotonin: Role in mood disorders. Neuropsychopharmacology 2008, 33, 73–83. [Google Scholar] [CrossRef]
- Meaney, M.J. Epigenetics and the biology of gene environment interactions. In Gene-Environment Transactions in Developmental Psychopathology; Tolan, P.H., Leventhal, B.L., Eds.; Springer: New York, NY, USA, 2017. [Google Scholar]
- Notaras, M.; Van deen Buuse, M. Neurobiology of BDNF in fear memory, sensitivity to stress, and stress-related disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef]
- Meyer, R.C.; Spangler, E.L.; Patel, N.; London, E.D.; Ingram, D.K. Impaired learning in rats in a 14-unit T-maze by 7-nitroindazole, a neuronal nitric oxide synthase inhibitor, is attenuated by the nitric oxide donor, molsidomine. Eur. J. Pharmacol. 1998, 341, 17–22. [Google Scholar] [CrossRef]
- Pitsikas, N. The nitric oxide (NO) donor molsidomine antagonizes scopolamine and L-NAME-induced performance deficits in a spatial memory task in the rat. Behav. Brain Res. 2009, 200, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Pitsikas, N.; Zisopoulou, S.; Sakellaridis, N. Nitric oxide donor molsidomine attenuates psychotomimetic effects of the NMDA receptor antagonist MK-801. J. Neurosci. Res. 2006, 84, 299–305. [Google Scholar] [CrossRef]
- Argyrousi, E.K.; Prickaerts, J. Nitrinergic signalling in object novelty recognition, Chapter 37. In Handbook of Behavioral Neuroscience; Ennaceur, A., de Souza Silva, M.A., Eds.; Elsevier: New York, NY, USA, 2018; pp. 561–565. [Google Scholar]
- Vanaja, P.; Ekambaram, P. Involvement of nitric oxide in learning & memory processes. Indian. J. Med. Res. 2011, 133, 471–478. [Google Scholar]
- Faria, M.; Miguel, T.; Gomes, K.; Nunes-de-Souza, R. Anxiety-like responses induced by nitric oxide within the BNST in mice: Role of CRF1 and NMDA receptors. Horm. Behav. 2016, 79, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Zarrabian, S.; Jamali, S.; Fazli-Tabaei, S.; Haghparast, A. The cross-talk between dopaminergic and nitric oxide systems in the medial septal nucleus, and their distinct effects on anxiety-like behaviors in male rats. J. Psychiatr. Res. 2021, 141, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Kurt, M.; Bilge, S.S.; Aksoz, E.; Kukula, O.; Celik, S.; Kesim, Y. Effect of sildenafil on anxiety in the plus-maze test in mice. Pol. J. Pharmacol. 2004, 56, 353–358. [Google Scholar] [PubMed]
- Roohbakhsh, A.; Moghaddam, A.H.; Massoudi, R.; Zarrindast, M.R. Role of dorsal hippocampal cannabinoid receptors and nitric oxide in anxiety like behaviours in rats using the elevated plus-maze test. Clin. Exp. Pharmacol. Physiol. 2007, 34, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Calixto, A.; Duarte, F.; Duzzioni, M.; Häckl, L.N.; Faria, M.; De Lima, T. Role of ventral hippocampal nitric oxide/cGMP pathway in anxiety-related behaviors in rats submitted to the elevated T-maze. Behav. Brain Res. 2010, 207, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Papageorgoulis, A.; Fallon, P.; Mpalantes, N.; Papageorgouli, D.; Pitsikas, N. Repeated but not acute exposure with a low dose range of the nitric oxide (NO) donor sodium nitroprusside (SNP) induces anxiolytic-like behaviour in a dose-independent manner in two different rat models of anxiety. Nitric Oxide 2020, 99, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.; Vicente, M.; Cipriano, A.; Miguel, T.; Nunes-de-Souza, R. Functional lateralization of the medial prefrontal cortex in the modulation of anxiety in mice: Left or right? Neuropharmacology 2016, 108, 82–90. [Google Scholar] [CrossRef]
- Volke, V.; Wegener, G.; Vasar, E. Augmentation of the NO-cGMP cascade induces. J. Physiol. Pharmacol. 2003, 54, 653–660. [Google Scholar]
- Orfanidou, M.A.; Lafioniatis, A.; Trevlopoulou, A.; Touzlatzi, N.; Pitsikas, N. Acute and repeated exposure with the nitric oxide (NO) donor sodium nitroprusside (SNP) differentially modulate responses in a rat model of anxiety. Nitric Oxide 2017, 69, 56–60. [Google Scholar] [CrossRef]
- Hernández-Melesio, M.A.; Alcaraz-Zubeldia, M.; Jiménez-Capdeville, M.E.; Martínez-Lazcano, J.C.; Santoyo-Pérez, M.E.; Quevedo-Corona, L.; Gerónimo-Olvera, C.; Sánchez-Mendoza, A.; Ríos, C.; Pérez-Severiano, F. Nitric oxide donor molsidomine promotes retrieval of object recognition memory in a model of cognitive deficit induced by 192 IgG-saporin. Behav. Brain Res. 2019, 366, 108–117. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Distance Traveled (cm) | Global Motor Activity | |
|---|---|---|
| Control | 248.5 ± 42.35 | 165.4 ± 28.75 |
| Mg | 149.6 ± 18.52 ** | 107.7 ± 10.82 ** |
| MSD | 192.2 ± 34.67 * | 122.5 ± 16.55 * |
| V-pyrro/NO | 227.8 ± 39.33 | 149.3 ± 31.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mititelu-Tartau, L.; Bogdan, M.; Pavel, L.L.; Rezus, C.; Foia, C.I.; Dima, N.; Gurzu, I.L.; Pelin, A.-M.; Buca, B.R. Anxiolytic-like Activity, Antioxidant Properties, and Facilitatory Effects on the Short-Term Memory Retention of Molsidomine in Rats. Life 2024, 14, 306. https://doi.org/10.3390/life14030306
Mititelu-Tartau L, Bogdan M, Pavel LL, Rezus C, Foia CI, Dima N, Gurzu IL, Pelin A-M, Buca BR. Anxiolytic-like Activity, Antioxidant Properties, and Facilitatory Effects on the Short-Term Memory Retention of Molsidomine in Rats. Life. 2024; 14(3):306. https://doi.org/10.3390/life14030306
Chicago/Turabian StyleMititelu-Tartau, Liliana, Maria Bogdan, Liliana Lăcrămioara Pavel, Ciprian Rezus, Cezar Ilie Foia, Nicoleta Dima, Irina Luciana Gurzu, Ana-Maria Pelin, and Beatrice Rozalina Buca. 2024. "Anxiolytic-like Activity, Antioxidant Properties, and Facilitatory Effects on the Short-Term Memory Retention of Molsidomine in Rats" Life 14, no. 3: 306. https://doi.org/10.3390/life14030306