
Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber

1
Zoological Museum, Moscow Lomonosov State University, Bol’shya Nikitskaya 2, Moscow 125009, Russia
2
I. I. Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Bogdan Khmielnitsky Str. 15, 01030 Kiev, Ukraine
3
Natural History Museum of Denmark, Universitetsparken 15, 2100 Copenhagen, Denmark
4
A.A. Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow 117647, Russia
5
Paleontological Laboratory, Cherepovets State University, Cherepovets 162600, Russia
*
Authors to whom correspondence should be addressed.
Life 2023, 13(3), 636; https://doi.org/10.3390/life13030636
Submission received: 12 January 2023
/
Revised: 21 February 2023
/
Accepted: 21 February 2023
/
Published: 24 February 2023
(This article belongs to the Special Issue Recent Research on Palaeontology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Xenophagus simutniki sp. n. is described from a late Eocene Rovno amber specimen. The new species is similar to the fossil Xenophagus popovi Lyubarsky et Perkovsky, 2017 from the late Eocene Baltic amber (W Russia), differing in the medially notched anterior margin of the pronotum. The Rovno xenosceline fauna is the richest among both extant and extinct faunas. This fauna includes the extinct genera Xenophagus Lyubarsky & Perkovsky, 2017 and Xenohimatium Lyubarsky & Perkovsky, 2012, which are closest to the extant Mediterranean Xenoscelis Wollaston 1864 and the representative of the extant boreal genus Zavaljus Reitter, 1880. A key to extinct species of the subfamily Xenoscelinae is presented. The possible reasons of xenoscelines abundance in European amber forests are discussed.
1. Introduction
Erotylidae, pleasing fungus beetles and lizard beetles, are distributed worldwide and include approximately 3200 species in 300 genera and six subfamilies [1,2]. The classification of the group was most recently revised by Leschen [2], who combined the families Languriidae and Erotylidae based on a parsimony analysis [1,3,4]. One of the major problems remaining with the classification of erotylids is the paraphyly of Xenoscelinae and Loberinae (details are discussed in Leschen [2]). The family contains phytophagous, mycophagous, and saprophagous species, and a few other taxa that feed on pollen and dead wood. Most species are free living, but–as noted by Leschen and Buckley [5]—Chasmatodera Arrow, 1943 and Bancous Pic, 1946 (Erotylinae from Afrotropical and Oriental region) are known from nests of the fungus-cultivating termites [6], and wingless species of Loberopsyllus Martinez & Barrera, 1966 (Cryptophilinae from southern Mexico and Costa Rica) occur phoretic on neotomine rodents [7]. Lepidotoramus Leschen, 1997 (Cryptophilini from Amazon Basin) may be endoparasitic on Lepidoptera pupae [8]. The data mainly refer to the biology of adult beetles; there is much less information on the development of larvae.
Several extant genera have been recently erected in the subfamily Xenoscelinae with the list given in Lyubarsky and Perkovsky [9]. In last the last few years, four new extinct Xenoscelinae genera have been described (Microzavaljus Lyubarsky & Perkovsky, 2018; Warnis Lyubarsky, Perkovsky & Alekseev, 2016; Xenophagus Lyubarsky & Perkovsky, 2017; Xenohimatium Lyubarsky & Perkovsky, 2012) resulting in a total of 12.
Several Erotylidae fossils are known from the late Eocene amber faunas: erotyline Triplax contienensis Alekseev, 2014, languriine Serramorphus rasnitsyni Lyubarsky & Perkovsky, 2017, and xenosceline Microzavaljus saxonicum Lyubarsky & Perkovsky, 2018 in Bitterfeld amber [9,10,11], languriine Thallisellites olgae Kupryjanowicz, Lyubarsky & Perkovsky, 2021 in Baltic amber, Cycadophila mumia Alekseev, 2017 (Pharaxonothinae), xenoscelines Warnis tvanksticus Lyubarsky, Perkovsky & Alekseev, 2016 and Xenophagus popovi Lyubarsky & Perkovsky, 2017 in Baltic amber [12,13,14,15], xenoscelines Xenohimatium rovnense Lyubarsky & Perkovsky, 2012 [16], the new species of Xenophagus Lyubarsky & Perkovsky, 2017 described in the present paper, and Zavaljus lyubarskyi Alekseev & Bukejs, 2022 [17] in Rovno amber.
Rovno amber is the southern coeval of the famous Baltic amber from Rovno, Kiev (western part), Volyn and Zhitomir regions of Ukraine [18,19,20] and adjacent districts of Belarus [21]. Rovno amber biota was reviewed in [22,23,24], [25] and references therein). More than 70 beetle species from 35 families [17,19,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53] are reported; less than 15% of Rovno beetle species are known from Baltic amber [53,54,55] and references therein.
The purpose of this work was to describe the third erotylid species from Rovno amber and analyze the reasons for the relict nature of the modern distribution of the tribe.
2. Material and Methods
The new Rovno amber pleasing fungus beetle specimen was purchased from an amber dealer, who obtained the amber mostly in the Varash district of the Rovno region, the most important source of the new Rovno amber taxa in the last few years [30,32,33,38,40,41,45,50,51,52,56,57,58,59]. It was included in the clear piece of amber, weighting 7.6 g after the primary treatment. The specimen was cut and polished by Anatoly P. Vlaskin (SIZK) for the best visibility of the fossil to the subrectangular piece 10 × 6.5 × 1.6 mm.
Morphological terminology for the body parts of the beetles is following the work of Leschen [2].
Photographs were taken at the Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine (Kiev, SIZK) using the microscope Leica Z16 APO stereomicroscope equipped with a Leica DFC 450 camera. Dimensions were measured by 10 × 23 Leica 10450023Widefield Adjustable Eyepiece on Leica MZ APO.
Eleven extant specimens cited in the text were collected by late Alexey Yu. Isaev (Ulyanovsk State University, Russia) and are deposited in the Zoological Museum of Lomonosov State University (Moscow, ZMSU).
The holotype is deposited in collection of Schmalhausen Institute of Zoology.
3. Results
Systematic Paleontology
- Order Coleoptera Linnaeus, 1758
- Superfamily Cucujoidea Latreille, 1802
- Family Erotylidae Latreille, 1802
- Subfamily Xenoscelinae Ganglbauer, 1899
- Genus Xenophagus Lyubarsky & Perkovsky, 2017
- Type species: Xenophagus popovi Lyubarsky & Perkovsky, 2017
- Species composition. Type species, Xenophagus simutniki sp.n.
- Xenophagus simutniki Lyubarsky & Perkovsky, sp.n.
- http://zoobank.org/urn:lsid:zoobank.org:act:97E43669-56D8-4103-BC4E-D273399E242A (accessed on 20 February 2023)
- Etymology. Named in honor of our friend, hymenopterist Dr. Sergej A. Simutnik.
- MATERIAL. Holotype: SIZK UA-28134, Rovno amber, late Eocene.
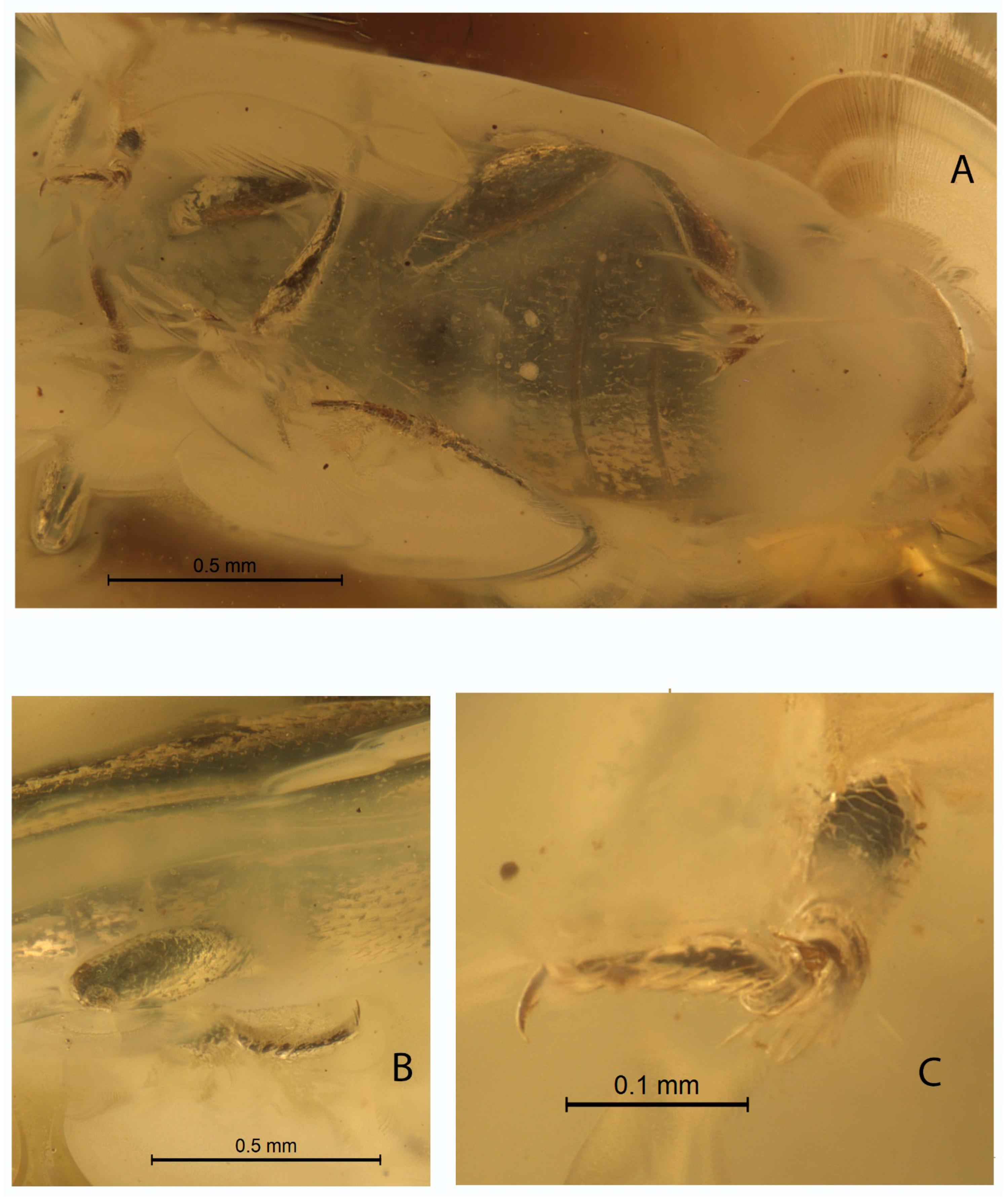
- DESCRIPTION. Body parallel-sided, 2.5 times longer than wide, dorsal vestiture short and decumbent (Figure 1A), dorsum weakly convex. Body length 2.7 mm, width 0.9 mm.
Head width equals 0.8 of the pronotal width. Compound eye hemispherical, comparatively large. Facets moderate in size, approximately equal to diameter of dorsal head puncture. Dorsal punctuation of head: punctures medium in size, distance between neighboring punctures equal to 1.5 diameter of puncture. Antennae 11-merous with three-segmented club, comparatively short, not extending beyond posterior margin of pronotum when directed posteriad. Antennal club slightly flattened. Antennomeres 9–10 transverse (Figure 1A). Antennomeres 3–5 very slightly elongate, 6th antennomere as long as wide, 7th and 8th antennomeres slightly transverse. Terminal antennomere as long as wide, with rounded tip. Antennomeres 3–7 subequal in length. Antennal furrows not present.
Pronotum parallel-sided, without lateral callosity and denticles, pronotal length 0.7 times the width, about 0.4 the elytral length. Anterior margin with median notch (Figure 1A). Lateral pronotal margin slightly flattened at anterolateral angles. Lateral margins and base of pronotum bordered. Base of pronotum with shallow, transverse depression; basal pits not present, basal furrow not present (Figure 1C). Posterior pronotal margin with basal lobe. Pronotum strongly and densely punctured, intervening spaces about 1.5 times as wide as puncture diameters. Posterolateral angles rectangular.
Intercoxal distance between mesocoxae 1.5 times shorter than width of mesocoxa. Intercoxal distance between metacoxae greater than diameter of metacoxa. Pre-, meso-, and metasternum strongly punctured. Legs slender, tibia slightly dilated apically. Tibia with bristle-like “crown” at apex, with short terminal spurs. Tarsomeres elongated, not lobate (Figure 2B,C). Tarsomeres 1–4 about same length, fifth tarsomere the longest, nearly as long as combined length of remaining tarsomeres (Figure 2B). Claw without notches, smooth, about 1/4 length of tarsomere 5.
Scutellar shield transverse, 2.5 times as long as wide. Elytral length 1.9 times as long as wide. Elytra narrowed and rounded apically. Elytra strongly punctured, intervening spaces about twice as wide as puncture diameters. Much smaller, secondary punctures are visible on the intervening spaces. Elytral surface not shagreened. Lateral margin of elytra is weakly distinguishable from above because of the boundaries between the layers of amber.
Abdominal ventrite 1 twice as long as ventrite 2 (Figure 2A). Submetacoxal lines present (Figure 2A). Abdominal punctures with bristles.
Key to extinct species of subfamily Xenoscelinae
- 1.
- —
- Submetacoxal lines not present……………………………………………………………….……4
- 2.
- Elytral punctures irregular. .…………………………………………………………………………3
- —
- Elytral punctures arranged in rows. Baltic amber ………… Warnis tvanksticus Lyubarsky, Perkovsky & Alekseev, 2016
- 3.
- Anterior margin of pronotum notched (Figure 1A). Rovno amber..……Xenophagus simutniki Lyubarsky & Perkovsky, sp. n.
- —
- Anterior margin of pronotum without notch. Baltic amber …………Xenophagus popovi Lyubarsky & Perkovsky, 2017
- 4.
- Pronotum widest across apical one third. ..……………………………………………………5
- —
- Pronotum almost completely parallel-sided. Rovno amber..………… Xenohimatium rovnense Lyubarsky & Perkovsky, 2012
- 5.
- Antenna long, extending beyond posterior edge of pronotum when directed posteriad. Rovno amber ..………Zavaljus lyubarskyi Alekseev & Bukeis, 2022
- —
- Antenna short, extending towards midlength of pronotum when directed posteriad. Bitterfeld amber ………Microzavaljus saxonicus Lyubarsky & Perkovsky, 2018
4. Discussion
Hitherto, no erotylid genera were known simultaneously from all three studied late Eocene European amber faunas. In that respect, this family differs, for instance, from cryptophagids, where all yet known Rovno (and Bitterfeld) amber genera are extant and known from the Baltic amber as well [54].
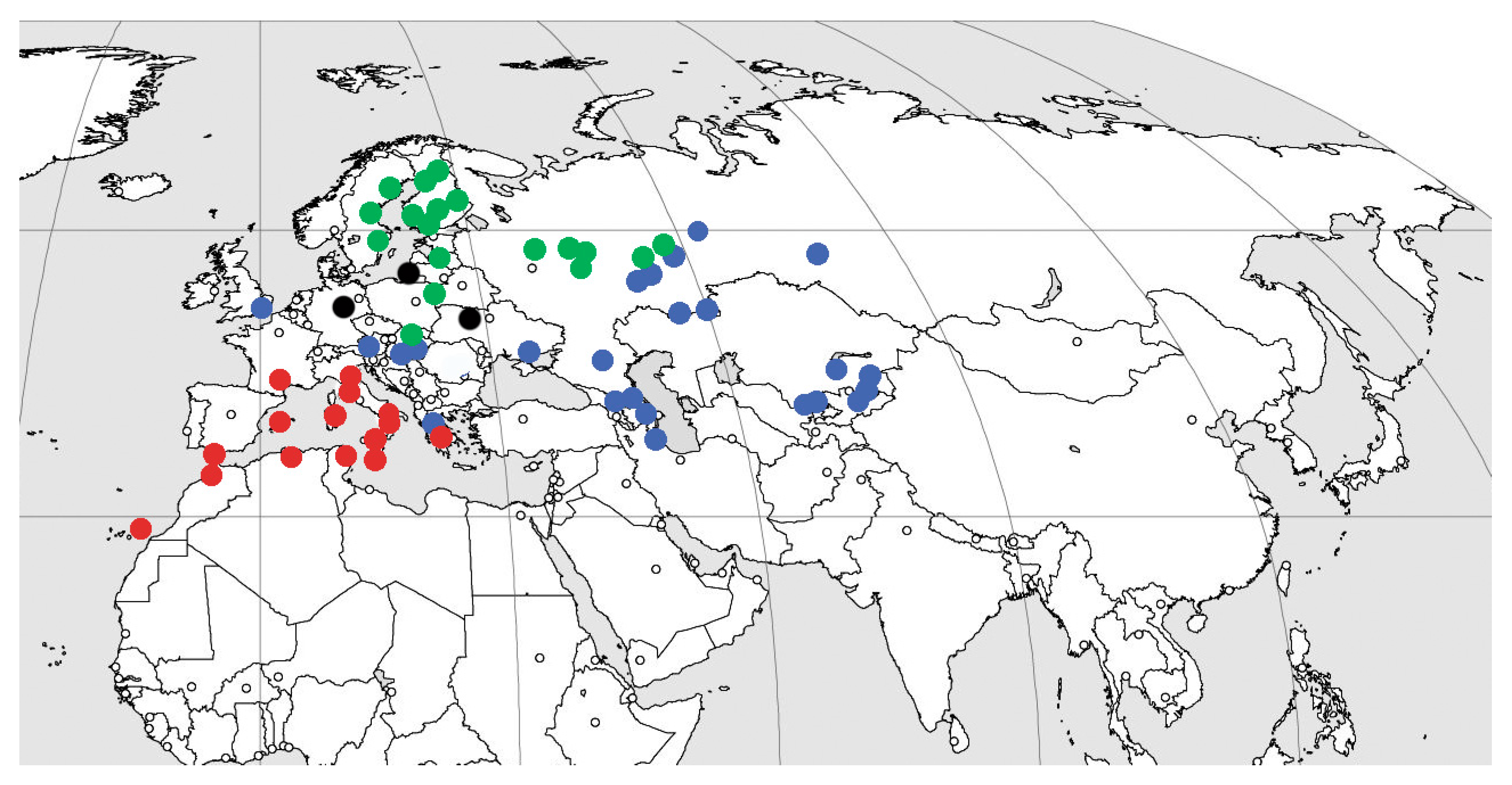
Subfamily Xenoscelinae comprises the following extant genera: New Zealand: Loberonotha Sen Gupta & Crowson, 1969 (one species); Neotropical: Othniocryptus Sharp, 1900 (one species; Panama, South America); Australian: Protoloberus Leschen, 2003 (one species; Queensland, New South Wales); Afrotropical: Arrowcryptus Leschen & Wegrzynowicz, 2008 (two species, tropical Africa), Xenocryptus Arrow, 1929 (two species; South Africa and Western Australia); Palaearctic (Figure 3): Xenoscelis Wollaston, 1864 (Mediterranean, one species); Zavaljus Reitter, 1880 (one extant species from Northern and north of Eastern Europe) and Macrophagus Motschulsky, 1845 (one species; from Austria, Hungary and Slovakia to Gilan and Kyrgyzstan). Xenoscelinae also includes four extinct genera (Xenohimatium from Rovno amber, Warnis from Baltic amber, Xenophagus from Baltic and Rovno amber, and Microzavaljus from Bitterfeld amber).
In each region and each amber Lagerstätte, one–two species of each genus are present. Three genera and three species that occur sympatrically in Rovno amber is the maximum yet observed local diversity for this subfamily, while the six species from European amber already represent 60% of the extant global diversity of the group and 71% of the genus-rank one.
The highest present-day diversity of the subfamily (three species in three genera) is recorded in Western Palaearctic. Yet even there, no distribution area overlapping is observed for different genera and species. Even in the eastern part of European Russia with the closest recorded distance between the populations of Zavaljus and Macrophagus, their nearest known localities are at least 150 km distant (Ulyanovsk Region, our data: Zavaljus from Palatovo village, Valgusskoye rural settlement, Inza District, under alder bark; Macrophagus from chalk steppe: Arskoye village, Ulyanovsk city district, 12 August 1990; Zykovo village, Novospasskoye urban settlement, 23 August 1990; Akulovka, Nikolaevsky District, 7 June 1990 and steppe (“mowing”): Shilovka village, Tushninskoye rural settlement, Sengileevsky District, 15 June 1987). This can be explained by the different ecological preferences of the mentioned taxa in present-day environments, i.e., dead wood association for Zavaljus brunneus (Gyllenhal, 1808) [60].
In contrast, adults of Xenoscelis deplanatus (Wollaston, 1862) are known to occur in dead stems of the Euphorbia shrubs, under decaying vegetation and under stones (especially in winter). It should be noted that Macrophagus is anthophilous, collected from bee nests (Anthophora Latreille 1803 and Halictus Latreille, 1804), and Zavaljus is more associated with wasps [61].
On the contrary, in European late Eocene amber faunas xenosceline genera obviously coexisted. Equally in contrast with the present times, Xenoscelinae dominated clearly in the late Eocene amber faunas of Europe, comprising a third of all known Erotylidae in Bitterfeld, half in Baltic, and being the only erotylid subfamily known from Rovno amber. In other words, xenoscelines encompassed half of all Erotylidae at the northern coast of Subparatethys and two thirds of them at its southern coast.
While the area of Russoscandia Island was comparable with Greenland, and the allopatry of the Baltic xenoscelines is theoretically possible, Volyn land with Rovno amber forest was rather small [62], so the xenoscelines sympatry was practically inevitable.
Extant fauna of xenoscelines includes three tropical species (33%) from the genera Othniocryptus and Arrowcryptus, two (22%) species from temperate regions (Figure 3), but the largest diversity of species—four (44%) are known from the subtropics, except for Xenoscelis, which is distributed all through the Southern Hemisphere. It is natural to assume that increasing seasonality of European post-Eocene climate led to a sharp decline in the abundance and diversity of xenoscelines, as well as to their transition to allopatric distribution. The transition to allopatric distribution was likely driven by narrowing and decrease in available ecological niches. Xenoscelines, adapted to equable climate, were likely quite preadapted for the areas with a Mediterranean climate, and part of them moved further south (genera Xenophagus and Xenohimatium most close to the extant Mediterranean genus Xenoscelis Wollaston, 1864; of course the existence of this lineage in the Eocene of Southern Europe could not be excluded as well); genus Zavaljus was able, on the contrary, to adapt in a similar way as Formica Linnaeus, 1758 and Lasius Fabricius, 1804 [63,64] to the strongly seasonal climate possible thanks to the symbiosis: for ants with aphids [65] and for Zavaljus with wasps (see below). The genus Zavaljus remains unknown even from Baltic amber deposits yielding immeasurably more beetle fossils than Rovno amber [18]. In any case, we do not consider the Eocene distribution of Zavaljus boreal. Extant species clearly prefer deciduous trees (when available) to gymnosperms. The adaptation by Zavaljus to boreal climate may be as well a result of its symbiotic association with wasp nests, which is not common in other xenoscelines [66], and its absence in the more southern regions could hypothetically have resulted from the stronger competition there. In turn, representatives of genus Macrophagus collected predominantly in steppe regions as well as in forests in the south of the temperate zone, with the single Asian subtropical record from Gilan (Figure 3).
An additional explanation of the extinction of xenoscelines could be their potential competition with Asian erotylids. Distribution of these taxa towards the west was limited by the presence of the large water bodies in the Eocene time. Perhaps not coincidentally, the Oriental (and Nearctic) region are the only zoogeographic units with no available xenosceline records; the single Eastern Palaearctic xenosceline species occurs only in regions adjacent to Western Palaearctic (Figure 3). At the same time, extant Oriental and Nearctic erotylid faunas are quite diverse, and the Oriental Region appears as one of the most important diversity centers of extant erotylids. Erotylidae evolved from a microfungal diet to saprophagy (then anthophyly) or phytophagy (then feeding on dead wood) [5]. The Macrophagus lineage switched from a microfungal to a pollen diet (perhaps through saprophagy), and the Zavaljus lineage from a microfungal to a dead wood diet (perhaps through phytophagy). Leschen and Buckley [5] (p. 108) indicated: “There could have been widespread extinction in Pharaxonothinae and Xenoscelinae, resulting in scant numbers of species per genus, leaving a hypodiverse present-day remnant of a vast radiation”. Perhaps, the extinction of Xenoscelinae was caused partially by their ancient relationship with cycads [5].
Xenoscelinae is one of the stem groups of the family Erotylidae. It is possible that the younger lineages of this family (Languriinae, Cryptophilinae) forced out the older lineage into the subtropics and temperate zone: the maximum diversity of Languriinae and Cryptophilinae is in the tropical regions, in the Oriental region and the Neotropics. Cryptophilinae is not present in European amber, Languriinae is as common as Xenoscelinae (Bitterfeld amber), twice less abundant (Baltic amber) or not present (Rovno amber); on the southern coast of Subparathetys, Languriinae likely were four times less abundant than Xenoscelinae.
It appears highly likely that the comparatively warmer climate of the Rovno amber forest [67] was more favourable to xenoscelines and many other thermophile forms [53] and references therein than the Baltic one, and this is the reason for their comparatively high diversity in the Rovno coleopterofauna [37,38,42,45,52,53,57,59,68,69,70].
All in all, the present distribution of Xenoscelinae appears evidently a relic. With similar extant distribution (not counting the three Western Palearctic species) these relic taxa are habitually named “Gondwanan” (see [71,72] and references therein). Still, the domination of xenoscelines in European amber forests was so evident that the adaptations they acquired there likely contributed to the further survival of the three extant Western Palearctic genera of the subfamily.
Author Contributions
G.Y.L. and E.E.P. designed the study. G.Y.L. and E.E.P. prepared the systematic placement of the new taxa, as well as prepared new taxa descriptions and plates. G.Y.L., D.V.V. and E.E.P. drafted the manuscript and contributed to the writing and discussion. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Alexandr P. Rasnitsyn (Paleontological Institute, Moscow, Russia) for his continuous, great support, Anatoly P. Vlaskin (SIZK) for cutting and polishing the amber pieces, Mykola R. Khomych (Rovno, Ukraine) for the help with obtaining the discussed specimen, Sergej A. Simutnik (SIZK) for imaging. We are obliged to Dmitry Telnov (Natural History Museum, London, United Kingdom) for proofreading the manuscript and the anonymous reviewers for their valuable comments. The work of the first co-author has been supported by a grant from the Zoological Museum AAAA A16-116021660077-3, the second was supported by Scholars at Risk Ukraine (SARU) fellowship at The University of Copenhagen.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Węgrzynowicz, P. Morphology, phylogeny and classification of the family Erotylidae based on adult characters (Coleoptera: Cucujoidea). Genus 2002, 13, 435–504. [Google Scholar]
- Leschen, R.A.B. Erotylidae (Insecta: Coleoptera: Cucujoidea): Phylogeny and review. Fauna New Zealand 2003, 47, 1–108. [Google Scholar]
- Leschen, R.A.B.; Węgrzynowicz, P. Generic catalogue and taxonomic status of Languriidae (Coleoptera: Cucujoidea). Ann. Zool. 1998, 48, 221–243. [Google Scholar]
- Robertson, J.A.; McHugh, J.V.; Whiting, M.F. A molecular phylogenetic analysis of the pleasing fungus beetles (Coleoptera: Erotylidae): evolution of colour patterns, gregariousness, and mycophagy. Syst. Entomol. 2004, 29, 173–187. [Google Scholar] [CrossRef]
- Leschen, R.A.B.; Buckley, T.R. Multistate characters and diet shifts: Evolution of Erotylidae (Coleoptera). Syst. Biol. 2007, 56, 97–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelley, P.E. Termitophilous pleasing fungus beetles: Chasmatodera Arrow and Rhamphidera Skelley, gen. nov. (Coleoptera: Erotylidae: Tritominae). Ann. Zool. 1999, 49, 255–263. [Google Scholar]
- Leschen, R.A.B.; Ashe, J.S. New species, phylogenetic placement, and mammal associations of Loberopsyllus (Languriidae: Xenoscelinae). In Entomological Contributions in Memory of Byron Alexander; Byers, G.W., Hagen, R.H., Brooks, R.W., Eds.; University of Kansas Natural History Museum Special Publication: Lawrence, KS, USA, 1999; Volume 24, pp. 171–177. [Google Scholar]
- Leschen, R.A.B. The Empocryptus-group (Languriidae: Toraminae) and a new genus associated with a lepidopteran cocoon. Coleopt. Bull. 1997, 51, 303–318. [Google Scholar]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Microzavaljus, a second extinct genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Bitterfeld Amber. Russ. Entomol. J. 2018, 27, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Alekseev, V.I. A new species of Triplax Herbst, 1793 (Coleoptera: Erotylidae) from Bitterfeld amber. Balt. J. Coleopterol. 2014, 14, 171–177. [Google Scholar]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Serramorphus, a new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia) from Late Eocene Bitterfeldian amber. Russ. Entomol. J. 2017, 26, 37–40. [Google Scholar] [CrossRef]
- Kupryjanowicz, J.; Lyubarsky, G.Yu.; Perkovsky, E.E. A new genus of the subfamily Languriinae (Coleoptera: Erotylidae) from the Late Eocene Baltic amber. Far East. Entomol. 2022, 447, 1–9. [Google Scholar] [CrossRef]
- Alekseev, V.; Bukejs, A. First fossil representatives of Pharaxonothinae Crowson (Coleoptera: Erotylidae): indirect evidence for cycads existence in Baltic amber forest. Zootaxa 2017, 4337, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Lyubarsky, G.Yu.; Perkovsky, E.E.; Alekseev, V.I. The first record of the subfamily Xenoscelinae (Coleoptera, Erotylidae) from the Baltic amber. Paleontol. J. 2016, 50, 963–969. [Google Scholar] [CrossRef]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Xenophagus, a new genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Baltic amber. Russ. Entomol. J. 2017, 26, 147–150. [Google Scholar] [CrossRef]
- Lyubarsky, G.Yu.; Perkovsky, E.E. A new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia). Russ. Entomol. J. 2012, 21, 35–38. [Google Scholar] [CrossRef]
- Alekseev, V.I.; Bukejs, A. A new extinct species of Zavaljus Reitter (Coleoptera: Erotylidae) from Rovno amber: boreal distribution range since the Eocene. Hist. Biol. 2022, 35, 315–319. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Zosimovich, V.Y.; Vlaskin, A.P. Chapter 7: Rovno amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Legalov, A.A.; Nazarenko, V.Yu.; Perkovsky, E.E. A new species of the genus Dorytomus Germar, 1817 (Coleoptera, Curculionidae) from Rovno amber. Zootaxa 2021, 5006, 95–100. [Google Scholar] [CrossRef]
- Martynov, A.V.; Vasilenko, D.V.; Perkovsky, E.E. First Odonata from Upper Eocene Rovno amber (Ukraine). Hist. Biol. 2022, 34, 2182–2187. [Google Scholar] [CrossRef]
- Perkovsky, E.E. Rovno amber caddisflies (Insecta, Trichoptera) from different localities, with information about three new sites. Vestn. Zool. 2017, 51, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Jałoszyński, P.; Perkovsky, E.E. Diversity of Scydmaeninae (Coleoptera: Staphylinidae) in Upper Eocene Rovno amber. Zootaxa 2016, 4157, 1–85. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Ignatov, M.S.; Remizowa, M.V.; Nuraliev, M.S.; Blagoderov, V.; Garbout, A.; Perkovsky, E.E. Staminate flower of Prunus s. l. (Rosaceae) from Eocene Rovno amber (Ukraine). J. Plant. Res. 2018, 131, 925–943. [Google Scholar] [CrossRef]
- Perkovsky, E.E. Only a half of species of Hymenoptera in Rovno amber is common with Baltic amber. Vestn. Zool. 2018, 52, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Mitov, P.G.; Perkovsky, E.E.; Dunlop, J.A. Harvestmen (Arachnida: Opiliones) in Eocene Rovno amber (Ukraine). Zootaxa 2021, 4984, 43–72. [Google Scholar] [CrossRef]
- Bukejs, A.; Háva, J.; Alekseev, V.I. A new fossil species of Attagenus Latreille (Coleoptera: Dermestidae) in Rovno and Baltic ambers, with a brief review of known fossil beetles from the Rovno amber Lagerstätte. Foss. Rec. 2020, 23, 95–104. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Kairišs, K. The first record of Cupedidae (Coleoptera: Archostemata) from Eocene Rovno amber: Cupes groehni Kirejtshuk, 2005 examined using X-ray microtomography. Balt. J. Coleopterol. 2021, 21, 111–116. [Google Scholar]
- Sokolov, A.V.; Perkovsky, E.E. The first Eocene species of Bacanius (Coleoptera: Histeridae: Dendrophilinae) from Rovno amber. Russ. Entomol. J. 2020, 29, 157–160. [Google Scholar] [CrossRef]
- Kazantsev, S.V.; Perkovsky, E.E. The first Cacomorphocerus species (Coleoptera, Cantharidae) from Rovno amber: A second species of the genus with 11-segmented antennae. Zootaxa 2020, 4751, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazantsev, S.V.; Perkovsky, E.E. Imprint of a Helcophorus Fairmaire, 1881: The first net-winged beetle (Coleoptera: Lycidae) from Rovno amber. Zootaxa 2022, 5128, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Kupryjanowicz, J.; Lyubarsky, G.Y.; Perkovsky, E.E. Heterhelus buzina sp.n. (Coleoptera: Kateretidae) from Rovno amber: The first proxy for Sambucus in the Eocene of Eastern Europe. Invertebr. Zool. 2021, 18, 16–24. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Perkovsky, E.E. Protomauroania mikhailovi—A new species of malachite beetles (Coleoptera, Dasytidae) in Rovno amber. Zootaxa 2021, 5006, 189–194. [Google Scholar] [CrossRef]
- Jałoszyński, P.; Perkovsky, E.E. A new bizarre species of Euconnus (Cladoconnus) in Upper Eocene Rovno amber (Coleoptera: Staphylinidae: Scydmaeninae). Zootaxa 2021, 5004, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Alekseev, V.I. Cathartosilvanus perkovskyi sp. nov.—first record of Silvanidae (Coleoptera: Cucujoidea) from Eocene Rovno amber. Balt. J. Coleopterol. 2021, 21, 19–24. [Google Scholar]
- Alekseev, V.I.; Bukejs, A. A new fossil genus of minute-clubbed beetles (Coleoptera: Cucujoidea: Monotomidae) from Paleogene amber of Europe. Hist. Biol. 2022, 35, 430–435. [Google Scholar] [CrossRef]
- Alekseev, V.I.; Bukejs, A. Extant genus of flat bark beetle (Coleoptera: Silvanidae) with a present-day Australian-southern South American disjunction discovered in Eocene Rovno amber. Zootaxa 2022, 5129, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Telnov, D.; Perkovsky, E.E.; Vasilenko, D.V.; Yamamoto, S. The first fossil Coleoptera record from the Volyn Region, Ukraine, with description of a new Glesoconomorphus (Coleoptera, Mycteridae) in syninclusion with Winterschmidtiidae (Acari) and a key to species. ZooKeys 2021, 1068, 189–201. [Google Scholar] [CrossRef]
- Telnov, D.; Perkovsky, E.E.; Kundrata, R.; Kairišs, K.; Vasilenko, D.V.; Bukejs, A. Revealing Palaeogene distribution of the Ptilodactylidae (Insecta: Coleoptera): The first Ptilodactyla Illiger, 1807 records from Rovno amber of Ukraine. Hist Biol. 2022, in press. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new species of the genus Glaesotropis Gratshev and Zherikhin, 1995 (Coleoptera, Anthribidae) from Rovno amber. Foss. Rec. 2021, 24, 1–7. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. Ceutorhynchus Germar (Coleoptera, Curculionidae) as proxy for Eocene Brassicaceae: First record of the genus from Rovno amber. J. Paleontol. 2022, 96, 379–386. [Google Scholar] [CrossRef]
- Legalov, A.A.; Vasilenko, D.V.; Perkovsky, E.E. The American tribes Anypotactini and Eudiagogini (Coleoptera, Curculionidae) in Eocene of Europe as indicators of Eocene climate with description a new species. Diversity 2022, 14, 767. [Google Scholar] [CrossRef]
- Legalov, A.A.; Vasilenko, D.V.; Perkovsky, E.E. New proxy for Moraceae in Priabonian of Europe: First record of the genus Demimaea Pascoe, 1870 (Coleoptera: Curculionidae) from Eocene Rovno amber. Hist. Biol. 2022, 1–7. [Google Scholar] [CrossRef]
- Legalov, A.A. Fossil history of bark-beetles (Coleoptera: Scolytidae) with descriptions of two new species. Hist. Biol. 2022, 1–11. [Google Scholar] [CrossRef]
- Lyubarsky, G.Y.; Bukejs, A. Two new fossil species of the genus Atomaria Stephens (Coleoptera: Cryptophagidae) from Eocene European amber with a key to species described from fossil resins. Zootaxa 2022, 5188, 283–289. [Google Scholar] [CrossRef]
- Matalin, A.V.; Perkovsky, E.E.; Vasilenko, D.V. First record of tiger beetles (Coleoptera, Cicindelidae) from Rovno amber with the description of a new genus and species. Zootaxa 2021, 5016, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, S.V.; Bocak, L. New genus of erotine net-winged beetles, Damzenium gen. nov. (Coleoptera: Lycidae), from Eocene Rovno amber. Zootaxa 2022, 5154, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Kirichenko-Babko, M.; Perkovsky, E.E.; Vasilenko, D.V. A new genus and species of Lebiini (Coleoptera: Carabidae) from late Eocene Rovno amber. Hist. Biol. 2022, 34, 436–442. [Google Scholar] [CrossRef]
- Kirichenko-Babko, M.; Perkovsky, E.E.; Vasilenko, D.V. Antephilorhizus zerovae sp. nov. (Carabidae: Lebiini), the second ground beetle species from Rovno amber. Hist. Biol. 2022, 34, 2424–2429. [Google Scholar] [CrossRef]
- Schmidt, J.; Scholz, S.; Will, K. Character analysis and descriptions of Eocene sphodrine fossils (Coleoptera, Carabidae) using light microscopy, micro-CT scanning, and 3D imaging. Dtsch. Entomol. Z. 2022, 69, 19–44. [Google Scholar] [CrossRef]
- Vitali, F.; Perkovsky, E.E. Poliaenus europaeus n. sp., the first cerambycid from Rovno amber (Coleoptera Cerambycidae). Hist. Biol. 2022, in press. [Google Scholar] [CrossRef]
- Haug, C.; Baranov, V.A.; Hörnig, M.K.; Gauweiler, J.; Hammel, J.U.; Perkovsky, E.E.; Haug, J.T. 35 million-year-old solid-wood-borer beetle larvae support the idea of stressed Eocene amber forests. Palaeobiodivers. Palaeoenviron. 2022, 1–10. [Google Scholar] [CrossRef]
- Yamamoto, S.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. First fossil species of ship-timber beetles (Coleoptera: Lymexylidae) from Eocene Rovno amber (Ukraine). Foss. Rec. 2022, 25, 65–74. [Google Scholar] [CrossRef]
- Nabozhenko, M.V.; Perkovsky, E.E. Oracula campbelli sp. n.—The first fossil darkling beetle (Coleoptera: Tenebrionidae: Alleculinae) from the late Eocene Rovno amber (Ukraine). Zootaxa 2023, 5230, 245–250. [Google Scholar] [CrossRef]
- Lyubarsky, G.Y.; Perkovsky, E.E. New findings of Cryptophagidae (Coleoptera: Clavicornia) from Baltic amber in the unbiased collection of the Paleontological Institute of RAS. Russ. Entomol. J. 2021, 30, 282–287. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Odnosum, V.K.; Nazarenko, V.Yu.; Vasilenko, D.V. First record of the genus Cyrtanaspis Emery (Insecta: Coleoptera: Scraptiidae) from Baltic amber. Paleontol. J. 2022, 56, 208–212. [Google Scholar] [CrossRef]
- Simutnik, S.A.; Perkovsky, E.E.; Vasilenko, D.V. Protaphycus shuvalikovi Simutnik gen. et sp. n. (Chalcidoidea, Encyrtidae, Encyrtinae) from Rovno amber. J. Hymenopt. Res. 2022, 91, 1–9. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Nel, A. A new Rovno amber termite genus (Isoptera, Rhinotermitidae) from Styr River basin. Palaeont. Electron. 2021, 24, a05. [Google Scholar] [CrossRef] [PubMed]
- Giłka, W.; Harbach, R.E.; Perkovsky, E.E. Mosquitoes (Diptera: Culicidae) in Eocene amber from the Rovno region, Ukraine. Zootaxa 2021, 5016, 257–270. [Google Scholar] [CrossRef]
- Olmi, M.; Guglielmino, A.; Vasilenko, D.V.; Perkovsky, E.E. Discovery of the first apterous pincer wasp from amber, with description of a new tribe, genus and species of Apodryininae (Hymenoptera, Dryinidae). Zootaxa 2022, 5162, 54–66. [Google Scholar] [CrossRef]
- Gimmel, M.L.; Ferro, M.L. General overview of saproxylic Coleoptera. In Saproxylic Insects: Diversity, Ecology and Conservation; Ulyshen, M.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 51–128. [Google Scholar]
- Horion, A. Faunistik der Mitteleuropäischer Käfer. Band VII.; Clavicornia, I. Teil (Sphaeritidae bis Phalacridae); Feyel: Überlingen, Germany, 1960; p. 346. [Google Scholar]
- Ivanov, V.D.; Melnitsky, S.I.; Perkovsky, E.E. Caddisflies from Cenozoic resins of Europe. Paleontol. J. 2016, 50, 485–493. [Google Scholar] [CrossRef]
- Radchenko, A.G.; Perkovsky, E.E. Wheeler’s dilemma revisited: First Oecophylla–Lasius syninclusion and other ant syninclusions in the Bitterfeld amber (late Eocene). Invertebr. Zool. 2021, 18, 47–65. [Google Scholar] [CrossRef]
- Radchenko, A.G.; Perkovsky, E.E.; Vasilenko, D.V. Formica species (Hymenoptera, Formicidae, Formicinae) in late Eocene Rovno amber. J. Hymen. Res. 2021, 82, 237–251. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Wegierek, P. Aphid-Buchnera-Ant symbiosis; or why are aphids rare in the tropics and very rare further south? Earth. Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 297–310. [Google Scholar] [CrossRef]
- Hilszczański, J.; Jaworski, T.; Plewa, R.; Ługowoj, J. Zavaljus brunneus (Gyllenhal, 1808)—a beetle species new to the Polish fauna (Coleoptera: Erotylidae). Genus 2014, 25, 421–424. [Google Scholar]
- Mänd, K.; Muehlenbachs, K.; McKellar, R.C.; Wolfe, A.P.; Konhauser, K.O. Distinct origins for Rovno and Baltic ambers: Evidence from carbon and hydrogen stable isotopes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 265–273. [Google Scholar] [CrossRef]
- Perkovsky, E.E. A new species of Micromalthidae (Coleoptera) from the Rovno amber: 1. Adult morphology. Paleontol. J. 2016, 50, 293–296. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new genus of fungus weevils (Coleoptera: Anthribidae) in Rovno amber. Foss. Rec. 2018, 21, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Olmi, M.; Eggs, B.; Capradossi, L.; van de Kamp, T.; Perkovsky, E.E.; Guglielmino, A.; Vasilenko, D.V. A new species of Bocchus from upper Eocene Rovno amber (Hymenoptera: Dryinidae). J. Hymenopt. Res. 2022, 92, 257–272. [Google Scholar] [CrossRef]
- Gumovsky, A.; Perkovsky, E.; Rasnitsyn, A. Laurasian ancestors and “Gondwanan” descendants of Rotoitidae (Hymenoptera: Chalcidoidea): What a review of Late Cretaceous Baeomorpha revealed. Cretac. Res. 2018, 84, 286–322. [Google Scholar] [CrossRef]
- Kolesnikov, V.B.; Turbanov, I.S.; Eskov, K.Yu.; Propistsova, E.A.; Bashkuev, A.S. First non-amber Mesozoic pseudoscorpion from Upper Triassic deposits of eastern Europe, with a description of two new fossil subfamilies (Arachnida, Pseudoscorpiones, Feaellidae). Pap. Palaeontol. 2022, 8, e1466. [Google Scholar] [CrossRef]
Figure 1.
Holotype Xenophagus simutniki sp.n., images: (A)—dorsal view, arrow indicates notch of pronotum; (B)—ventral view, arrow indicates submetacoxal line; (C)—left lateral view.
Figure 1.
Holotype Xenophagus simutniki sp.n., images: (A)—dorsal view, arrow indicates notch of pronotum; (B)—ventral view, arrow indicates submetacoxal line; (C)—left lateral view.

Figure 2.
Holotype Xenophagus simutniki sp.n., images: (A)—ventral view of abdomen with submesocoxal lines; (B)—lateral view of abdomen with metatarsus; (C)—metatarsus.
Figure 2.
Holotype Xenophagus simutniki sp.n., images: (A)—ventral view of abdomen with submesocoxal lines; (B)—lateral view of abdomen with metatarsus; (C)—metatarsus.

Figure 3.
Distribution of Palearctic xenosceline genera. Green dots—Zavaljus; blue dots—Macrophagus; red dots—Xenoscelis; black dots—Priabonian amber deposits: Baltic, Rovno, and Bitterfeld amber.
Figure 3.
Distribution of Palearctic xenosceline genera. Green dots—Zavaljus; blue dots—Macrophagus; red dots—Xenoscelis; black dots—Priabonian amber deposits: Baltic, Rovno, and Bitterfeld amber.

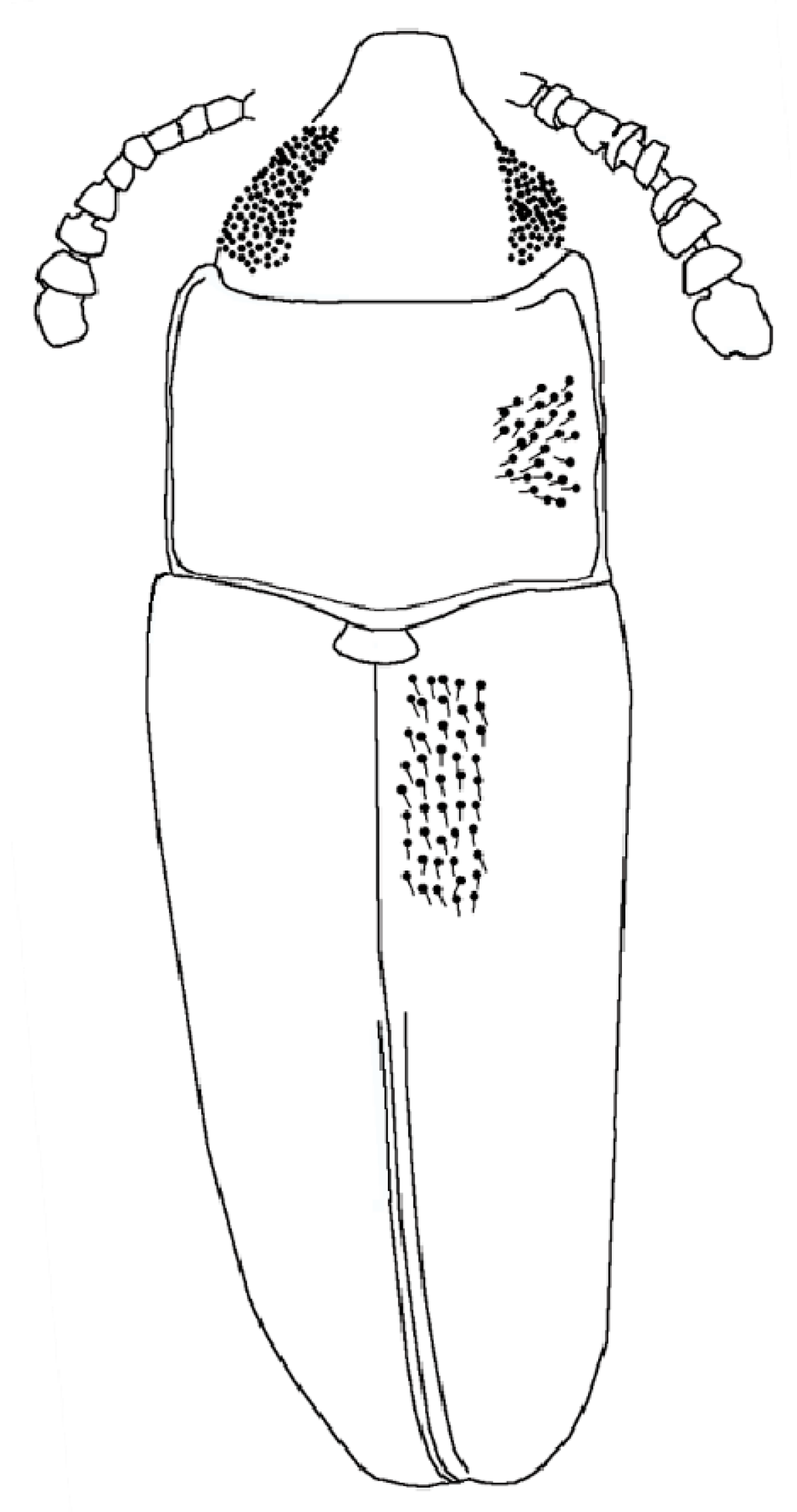
Figure 4.
Xenophagus simutniki sp.n., drawing, dorsal view.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lyubarsky, G.Y.; Perkovsky, E.E.; Vasilenko, D.V. Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber. Life 2023, 13, 636. https://doi.org/10.3390/life13030636
AMA Style
Lyubarsky GY, Perkovsky EE, Vasilenko DV. Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber. Life. 2023; 13(3):636. https://doi.org/10.3390/life13030636
Chicago/Turabian StyleLyubarsky, Georgy Yu., Evgeny E. Perkovsky, and Dmitry V. Vasilenko. 2023. "Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber" Life 13, no. 3: 636. https://doi.org/10.3390/life13030636
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.