
Exogenous Nitric Oxide and Silicon Applications Alleviate Water Stress in Apricots

 ,
,  and
and 
Abstract
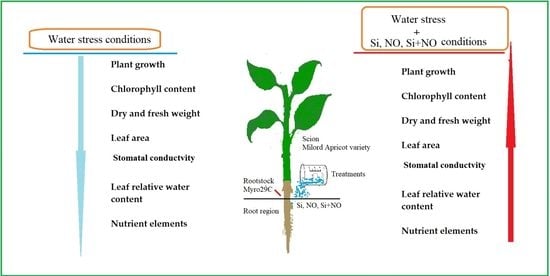
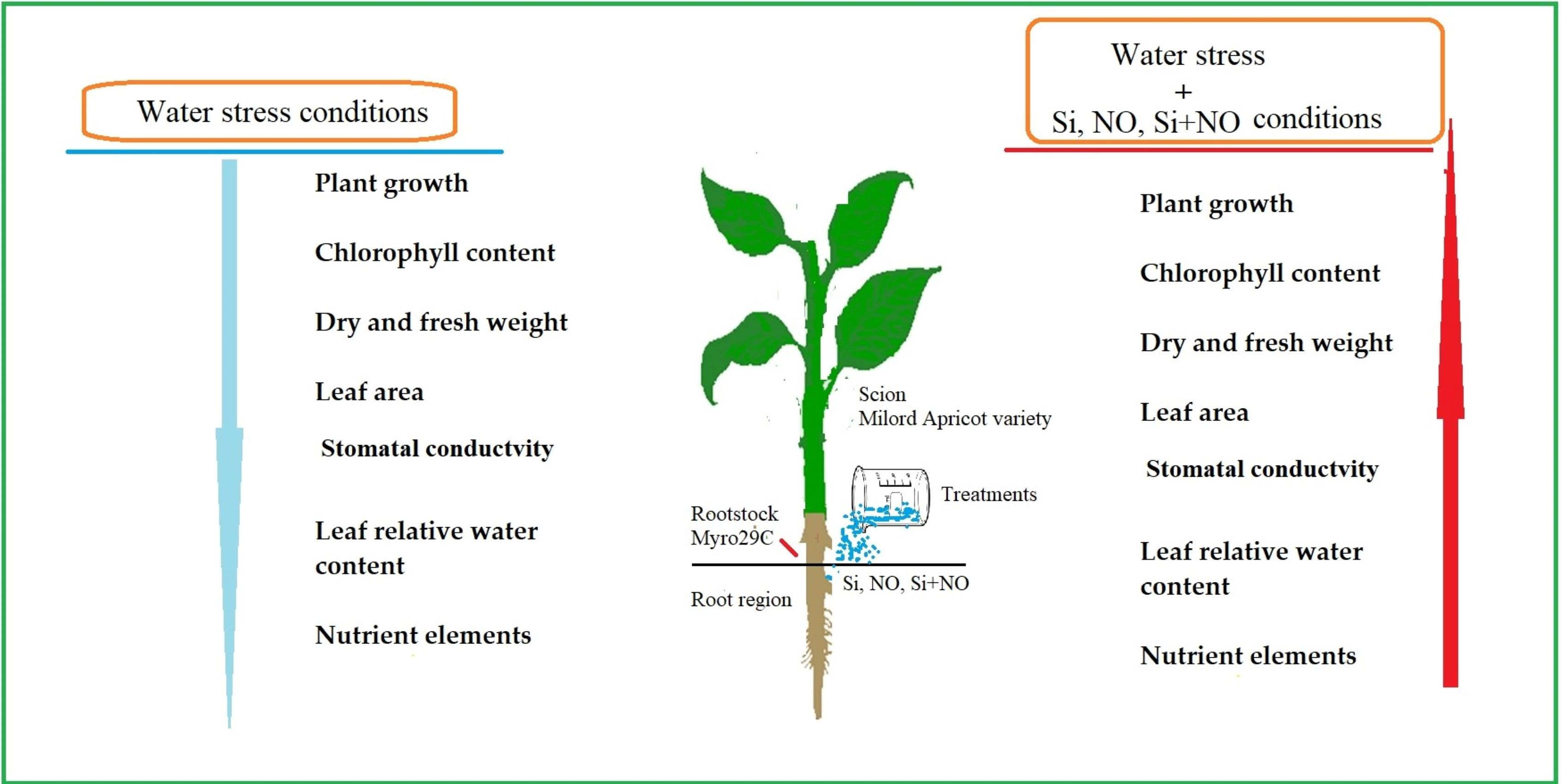
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Applications
2.2. Harvest in Saplings and Their Growth Parameters
2.3. Membrane Permeability (MP)
2.4. Leaf Relative Water Content (LRWC)
2.5. Stomatal Conductivity (SC)
2.6. Chlorophyll Content (Chl)
2.7. Leaf Temperature (LT), Proline Content and Lipid Peroxidation Analysis (MDA)
2.8. Mineral Analysis in Leaves
2.9. Statistical Analysis
3. Results
3.1. Plant Growth and Physiological Parameters
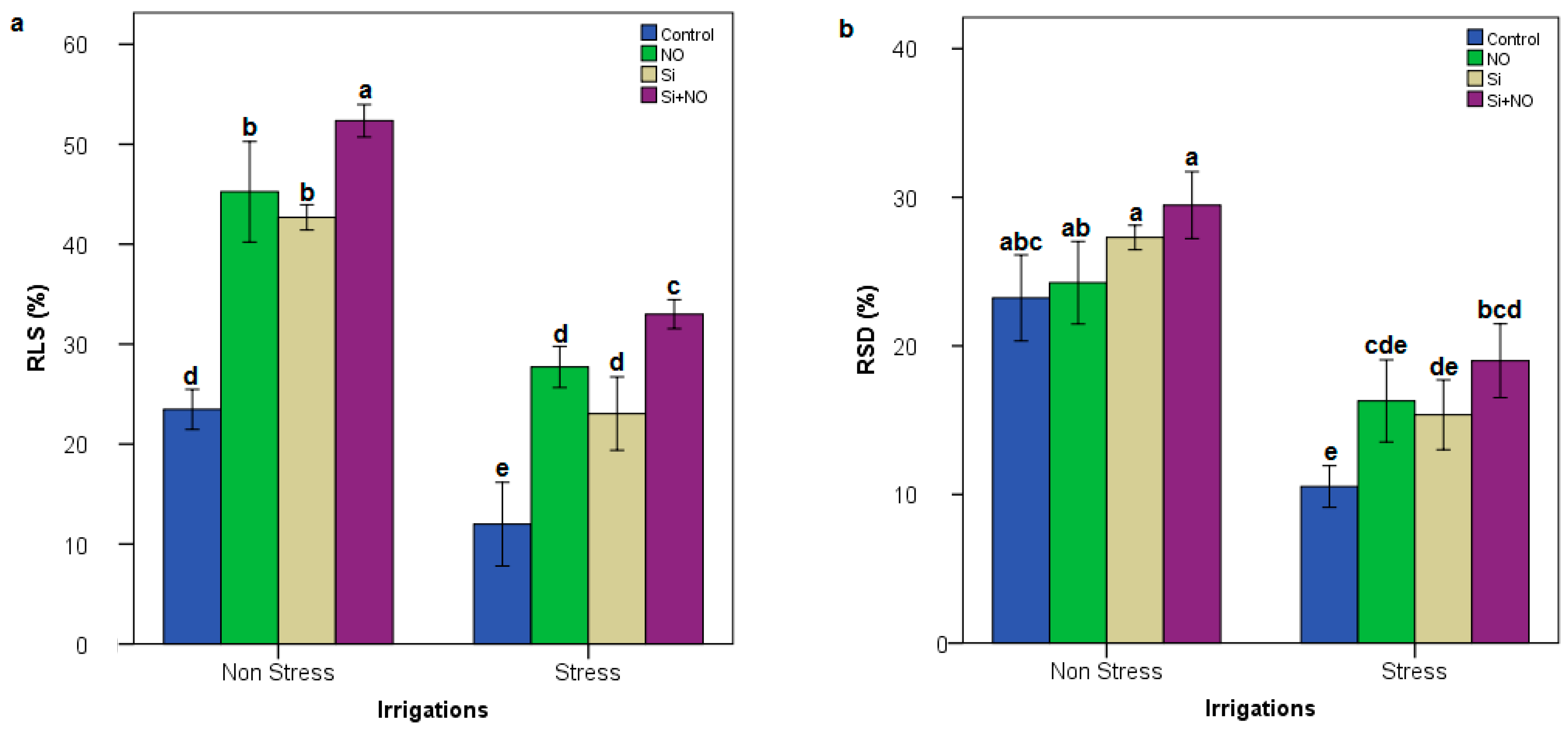
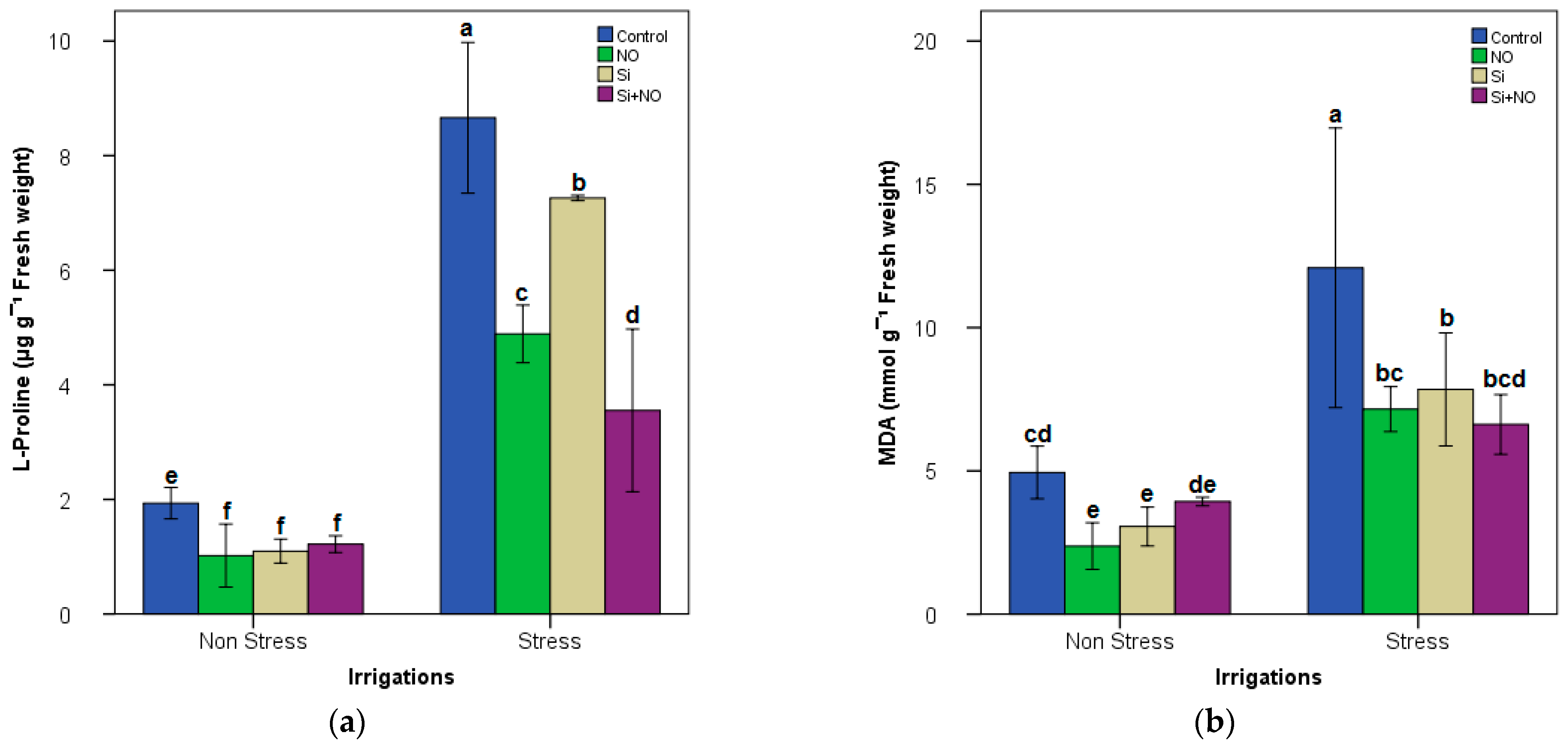
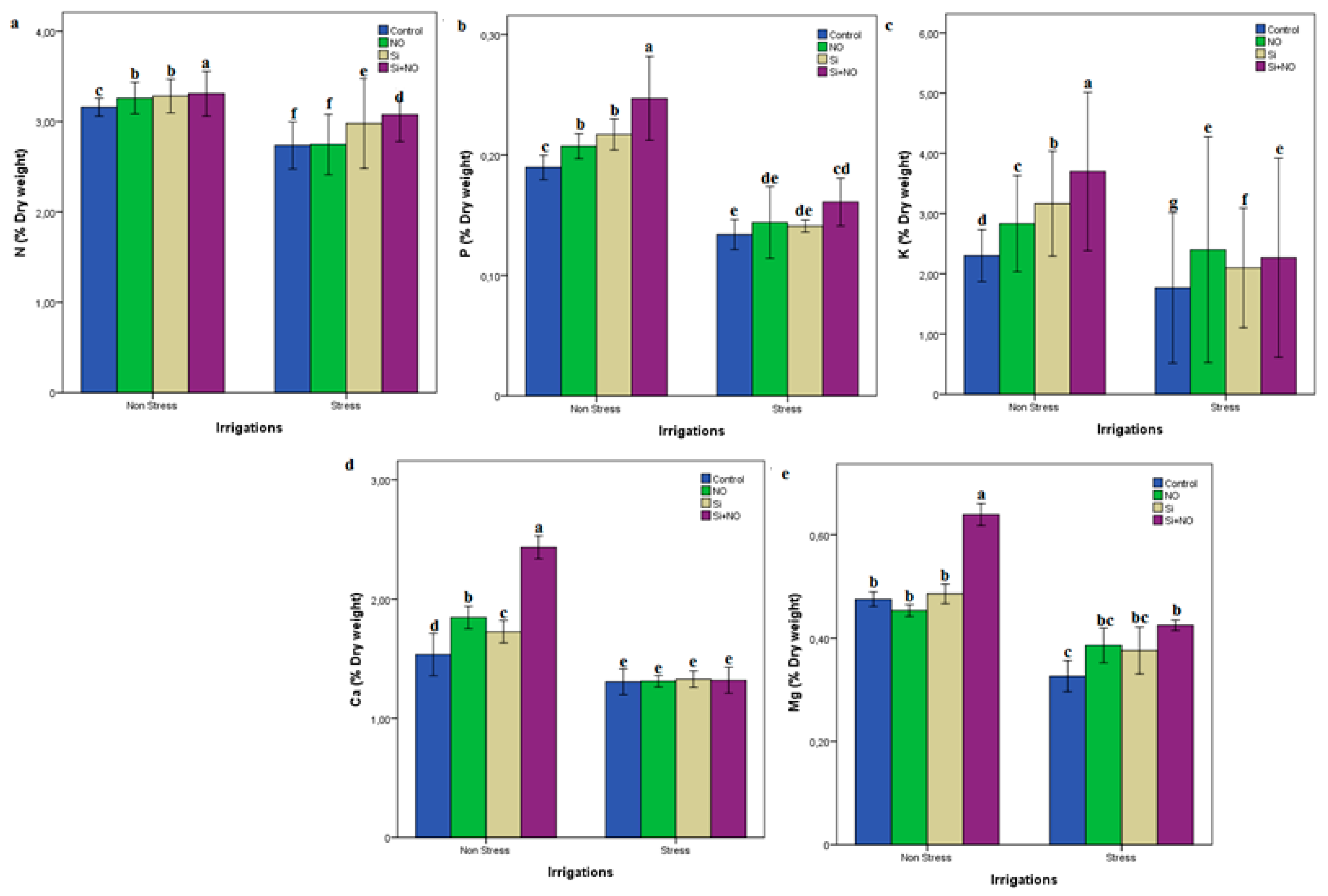
3.2. Biochemical Parameters
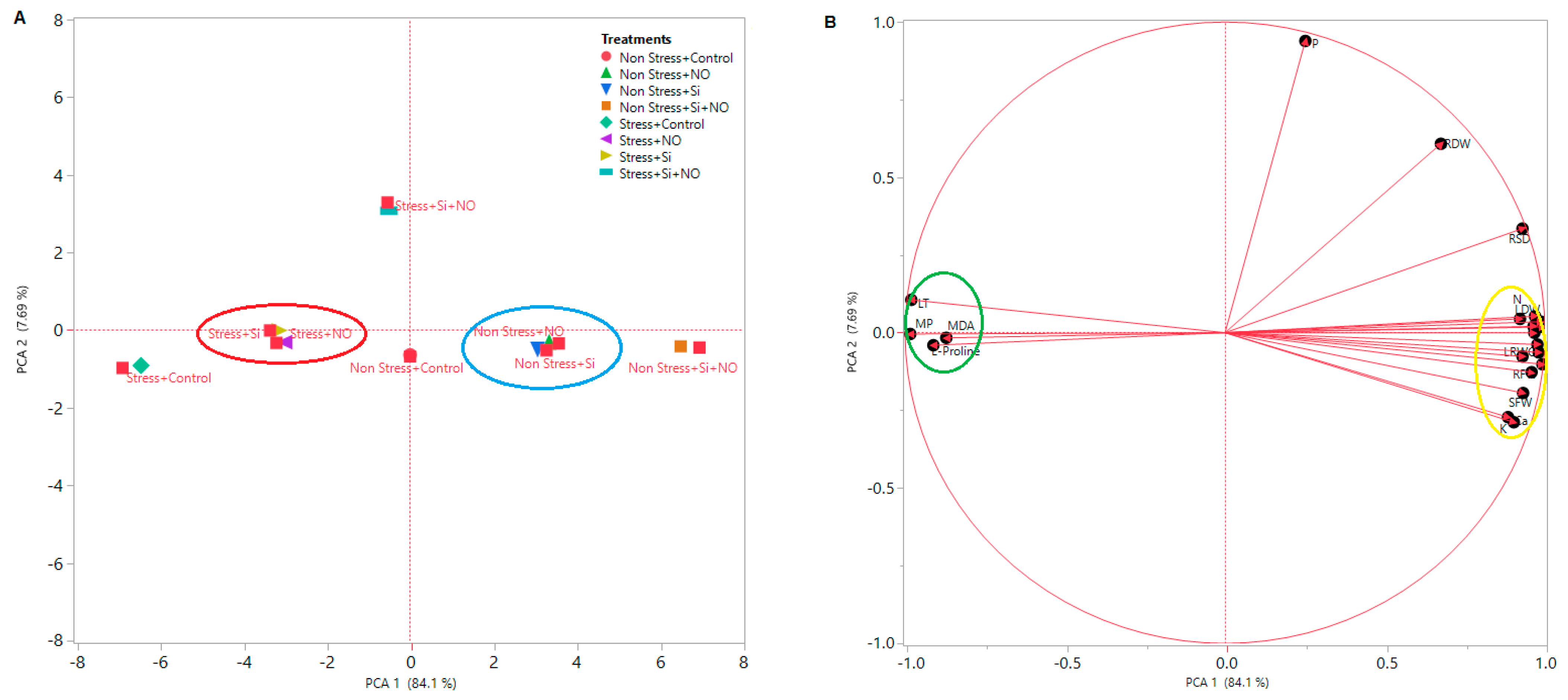
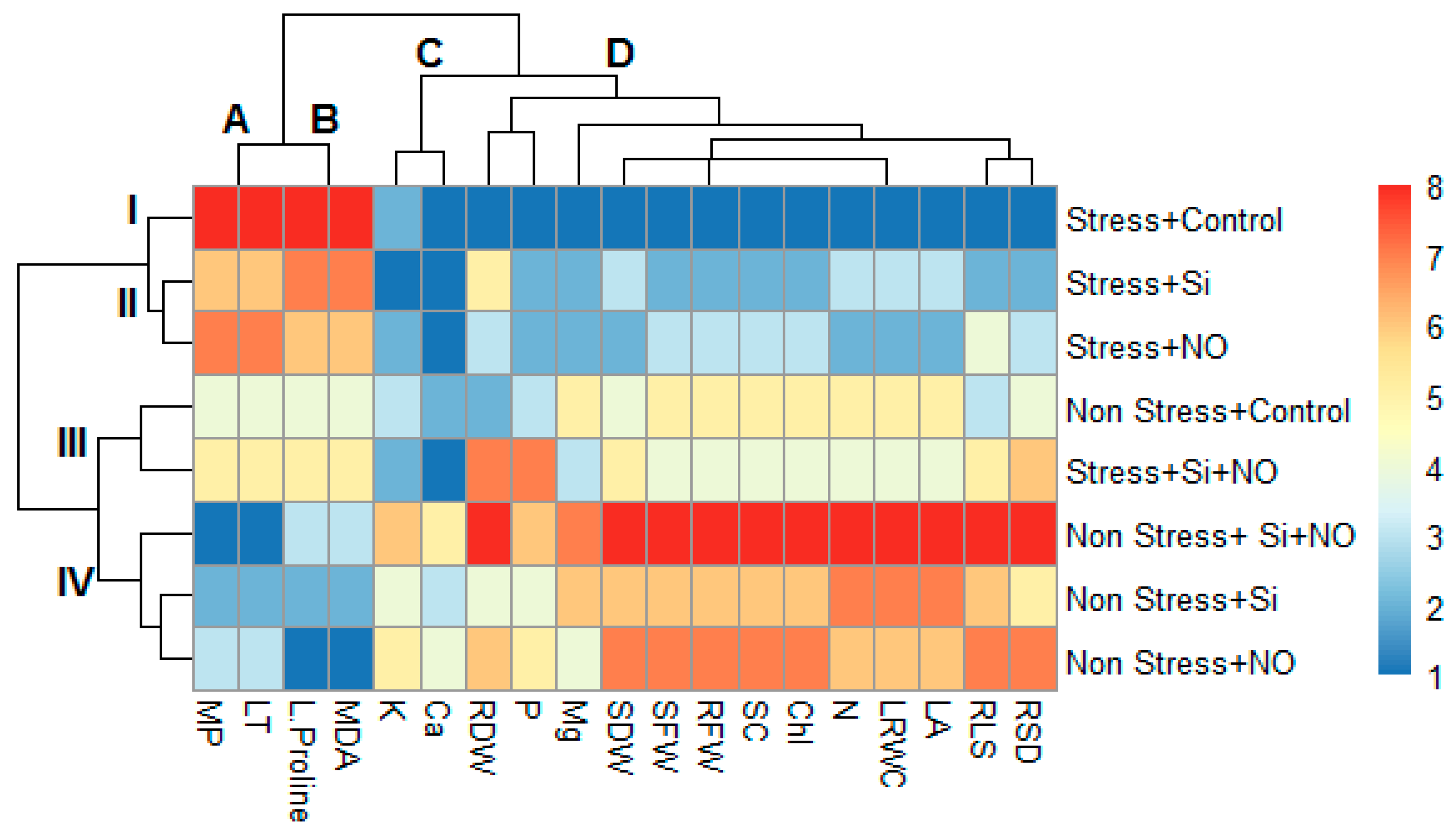
3.3. Multivariate Data Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The Role of Silicon in Higher Plants under Salinity and Drought Stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef] [PubMed]
- García-Mata, C.; Lamattina, L. Nitric Oxide Induces Stomatal Closure and Enhances the Adaptive Plant Responses against Drought Stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Biju, S.; Fuentes, S.; Gupta, D. The Use of Infrared Thermal Imaging as a Non-Destructive Screening Tool for Identifying Drought-Tolerant Lentil Genotypes. Plant Physiol. Biochem. 2018, 127, 11–24. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Partitioning of Assimilates to Deeper Roots is Associated with Cooler Canopies and Increased Yield under Drought in Wheat. Funct. Plant Biol. 2010, 37, 147–156. [Google Scholar] [CrossRef]
- Doncheva, S.; Poschenrieder, C.; Stoyanova, Z.; Velichkova, M.; Barceló, J. Silicon Amelioration of Manganese Toxicity in Mn-Sensitive and Mn-Tolerant Maize Varieties. Environ. Exp. Bot. 2009, 65, 189–197. [Google Scholar] [CrossRef]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity Stress Mitigation by Humic Acid Treatment in Strawberry (Fragaria x ananassa Duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- Alzahrani, Y.; Kuşvuran, A.; Hesham, F.; Alharby, H.F.; Kuşvuran, Ş.; Rady, M.M. The Defensive Role of Silicon in Wheat Against Stress Conditions Induced by Drought, Salinity or Cadmium. Ecotoxicol. Environ. Saf. 2018, 154, 187–196. [Google Scholar] [CrossRef]
- Bolat, I.; Bakır, A.G.; Korkmaz, K.; Gutiérrez-Gamboa, G.; Kaya, O. Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.). Agriculture 2022, 12, 1273. [Google Scholar] [CrossRef]
- Hernandez-Apaolaza, L. Can Silicon Partially Alleviate Micronutrient Deficiency in Plants? Springer Planta 2014, 240, 447–458. [Google Scholar] [CrossRef]
- Zhu, X.F.; Zhu, C.Q.; Wang, C.; Dong, X.Y.; Shen, R.F. Nitric Oxide Acts Upstream of Ethylene in Cell Wall Phosphorus Reutilization in Phosphorus-Deficient Rice. J. Exp. Bot. 2017, 68, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Torrecillas, A.; Galego, R.; Perez-Pastor, A.; Ruız-Sanchez, M. Gas Exchange and Water Relations of Young Apricot Plants under Drought Conditions. J. Agric. Sci. 1999, 132, 445–452. [Google Scholar] [CrossRef]
- Torrecillas, A.; Corell, M.; Galindo, A.; López, D.P.; Rodríguez, P.; Cruz, Z.N.; Centeno, A. Agronomical Effects of Deficit Irrigation in Apricot, Peach, and Plum Trees. In Water Scarcity and Sustainable Agriculture in Semiarid Environment; Academic Press: Cambridge, MA, USA, 2018; pp. 87–109. [Google Scholar]
- Mohammadkhani, N.; Heidari, R. Drought-induced Accumulation of Soluble Sugars and Proline in Two Maize Varieties. World Appl. Sci. J. 2008, 3, 448–453. [Google Scholar]
- El Mokh, F.; Nagaz, K.; Masmoudi, M.M.; Ben Mechlia, N.; Ghiglieri, G. Deficit Irrigation Using Saline Water of Fruit Trees under Water Scarcity Conditions of Southern Tunisia. Atmosphere 2021, 12, 864. [Google Scholar] [CrossRef]
- Hasan, M.M.; Alharby, H.F.; Hajar, A.S.; Hakeem, K.R.; Alzahrani, Y. The effect of magnetized water on the growth and physiological conditions of Moringa species under drought stress. Pol. J. Environ. Stud. 2019, 28, 1145–1155. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Calif. Agric. Exp. Stn. 1990, 347, 32. [Google Scholar]
- Klamkowskı, K.; Treder, W. Response to drought stress of three strawberry cultivars grown under greenhouse condition. J. Fruit Ornam. Plant Res. 2008, 16, 179–188. [Google Scholar]
- Bolat, I.; Dikilitas, M.; Ikinci, A.; Ercisli, S.; Tonkaz, T. Morphological, Physiological, Biochemical Characteristics and Bud Success Responses of Myrobolan 29C Plum Rootstock Subjected to Water Stress. Can. J. Plant Sci. 2016, 96, 485–493. [Google Scholar] [CrossRef]
- Lutts, S.; Kınet, J.M.; Bouharmont, J. NaCl-induced Senescence in Leaves of Rice (Oryza sativa L.) Varieties Differing in Salinity Resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Yamasaki, S.; Dillenburg, L.R. Measurements of Leaf Relative Water Content in Araucaria Angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Acevedo-Opazo, C.; Valdés-Gómez, H.; Taylor, J.A.; Avalo, A.; Verdugo-Vásquez, N.; Araya, M.; Jara-Rojas, F.; Tisseyre, B. Assessment of an Empirical Spatial Prediction Model of Vine Water Status for Irrigation Management in a Grapevine Field. Agric. Water Manag. 2013, 124, 58–68. [Google Scholar] [CrossRef]
- Khan, A.N.; Qureshi, R.H.; Ahmad, N. Salt Tolerance of Cotton Cultivars in Relation to Relative Growth Rate in Saline Environments. Int. J. Agric. Biol. 2004, 6, 786–787. [Google Scholar]
- Chen, C. Determining the Leaf Emissivity of Three Crops by Infrared Thermometry. Sensors 2015, 15, 11387–11401. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kacar, B.; Inal, A. Plant Analysis; Nobel Press: Ankara, Turkey, 2008. [Google Scholar]
- Shao, H.B.; Chu, L.Y.; Shao, M.A.; Abdul Jaleel, C.; Hong-Mei, M. Higher Plant Antioxidants and Redox Signaling under Environmental Stresses. C. R. Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef]
- Meeteren, U.V.; Kaiser, E.; Matamoros, P.M.; Verdonk, J.C.; Aliniaeifard, S. Is Nitric Oxide a Critical Key Factor in Aba-Induced Stomatal Closure? J. Exp. Bot. 2020, 71, 399–410. [Google Scholar] [CrossRef]
- Hasan, M.M.; Skalicky, M.; Jahan, M.S.; Hossain, M.N.; Anwar, Z.; Nie, Z.-F.; Alabdallah, N.M.; Brestic, M.; Hejnak, V.; Fang, X.-W. Spermine: Its Emerging Role in Regulating Drought Stress Responses in Plants. Cells 2021, 10, 261. [Google Scholar] [CrossRef]
- Alabdallah, N.M.; Hasan, M.M.; Hammami, I.; Alghamdi, A.I.; Alshehri, D.; Alatawi, H.A. Green synthesized metal oxide nanoparticles mediate growth regulation and physiology of crop plants under drought stress. Plants 2021, 10, 1730. [Google Scholar] [CrossRef]
- Hasan, M.M.; Gong, L.; Nie, Z.; Feng, X.; Ahammed, G.J.; Fang, X.W. ABA-induced stomatal movements in vascular plants during dehydration versus rehydration. Environ. Exp. Bot. 2021, 186, 104436. [Google Scholar] [CrossRef]
- Hasan, M.M.; Alharby, H.F.; Uddin, M.N.; Ali, M.A.; Anwar, Y.; Fang, X.W.; Hakeem, K.R.; Alzahrani, Y.; Hajar, A.S. Magnetized water confers drought stress tolerance in Moringa biotype via modulation of growth, gas exchange, lipid peroxidation and antioxidant activity. Pol. J. Environ. Stud. 2020, 29, 1625–1636. [Google Scholar] [CrossRef]
- Waseem, M.; Nie, N.F.; Yao, G.Q.; Hasan, M.M.; Xiang, Y.; Fang, X.W. Dew absorption by leaf trichomes in Caragana korshinskii: An alternative water acquisition strategy to withstand drought in arid environment. Physiol. Plant. 2021, 172, 528–539. [Google Scholar] [CrossRef]
- Hasan, M.M.; Ali, M.A.; Soliman, M.H.; Alqarawi, A.A.; Abd Allah, E.F.; Fang, X.W. Insights into 28-homobrassinolide (HBR)-mediated redox homeostasis, AsA–GSH cycle, and methylglyoxal detoxification in soybean under drought-induced oxidative stress. J. Plant Inter. 2020, 15, 371–385. [Google Scholar] [CrossRef]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Shiranibidabadi, S. Effect of Silicon on Growth and Development of Strawberry under Water Deficit Conditions. Hortic. Plant J. 2018, 4, 226–232. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric Oxide Regulates Plant Responses to Drought, Salinity, and Heavy Metal Stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Hasan, M.M.; Alharbi, B.M.; Alhaithloul, H.A.S.; Abdulmajeed, A.M.; Alghanem, S.M.; Al-Mushhin, A.A.M.; Jahan, M.S.; Corpas, F.J.; Fang, X.-W.; Soliman, M.H. Spermine-Mediated Tolerance to Selenium Toxicity in Wheat (Triticum aestivum L.) Depends on Endogenous Nitric Oxide Synthesis. Antioxidants 2021, 10, 1835. [Google Scholar] [CrossRef]
- Hasan, M.M.; Liu, X.D.; Waseem, W.; Guang-Qian, Y.; Alabdallah, N.M.; Jahan, M.S.; Fang, X.W. ABA activated SnRK2 kinases: An emerging role in plant growth and physiology. Plant Signal. Behav. 2022, 17, 2071024. [Google Scholar] [CrossRef] [PubMed]
- Servet, A.; Eşitken, A. Effects of Silicon to Salt Stress on Strawberry Plant. Harran Tarım ve Gıda Bilimleri Dergisi 2018, 22, 478–483. [Google Scholar]
- Sharif, P.; Amirnia, R.; Bidabadi, S.S. Role of silicon in mediating heat shock tolerance in soybean. Gesunde Pflanz. 2022, 74, 397–411. [Google Scholar] [CrossRef]
- Ipek, M. Determination of Garnem and Myrobolan 29C Rootstock Reactions against Drought Stress In Vitro Conditions. Ph.D. Thesis, Selçuk University, Konya, Turkey, 2015. Unpublished. [Google Scholar]
- Balotf, S.; Islam, S.; Kavoosi, G.; Kholdebarin, B.; Juhasz, A.; Ma, W. How Exogenous Nitric Oxide Regulates Nitrogen Assimilation in Wheat Seedlings under Different Nitrogen Sources and Levels. PLoS ONE 2018, 13, e0190269. [Google Scholar] [CrossRef]
- Ramos-Artuso, F.; Galatro, A.; Buet, A.; Santa-María, G.E.; Simontacchi, M. Key Acclimation Responses to Phosphorus Deficiency in Maize Plants Are Influenced by Exogenous Nitric Oxide. J. Plant Physiol. 2018, 222, 51–58. [Google Scholar] [CrossRef]
- Majeed, S.; Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Ejaz, S.; Ahmad, K.S.; Tauseef, S.; Farid, G.; Khalid, I.; Mehmood, K. Nitric oxide regulates water status and associated enzymatic pathways to inhibit nutrients imbalance in maize (Zea mays L.) under drought stress. Plant Physiol. Biochem. 2020, 155, 147–160. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Kapoor, D. Fascinating Regulatory Mechanism of Silicon for Alleviating Drought Stress in Plants. Plant Physiol. Biochem. 2021, 166, 1044–1053. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation Level | SFW (g/Plant) | SDW (g/Plant) | RFW (g/Plant) | RDW (g/Plant) | LA (cm2) |
|---|---|---|---|---|---|
| Non-Stress | 168.57 a | 88.92 a | 153.50 a | 43.61 a | 62.03 a |
| Stress | 108.35 b | 51.01 b | 88.04 b | 39.68 b | 41.76 b |
| t-test | * | * | * | * | * |
| Treatments | |||||
| Control | 110.80 d | 52.15 c | 89.41 c | 28.47 c | 40.11 b |
| Si | 137.04 c | 70.61 b | 122.05 b | 40.92 b | 56.03 a |
| NO | 142.50 b | 73.97 b | 123.82 b | 40.20 b | 51.48 a |
| Si + NO | 163.70 a | 83.13 a | 147.80 a | 56.99 a | 59.96 a |
| Tukey test | * | * | * | * | * |
| Irrigation Level | SC (mmol m−2 s−1) | MP (%) | Chl | LRWC (%) | LT (°C) |
|---|---|---|---|---|---|
| Non Stress | 112.96 a | 15.01 b | 43.47 a | 79.30 a | 31.92 b |
| Stress | 49.41 b | 19.83 a | 38.83 b | 73.32 b | 34.57 a |
| t-test | * | * | * | * | * |
| Treatments | |||||
| Control | 61.85 d | 19.80 a | 38.53 c | 73.26 d | 34.63 a |
| Si | 79.57 c | 16.85 c | 42.40 b | 77.32 b | 32.87 c |
| NO | 87.12 b | 17.73 b | 42.40 b | 75.80 c | 33.32 b |
| Si + NO | 96.19 a | 15.32 d | 43.05 a | 78.86 a | 32.15 c |
| Tukey test | * | * | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakır, A.G.; Bolat, I.; Korkmaz, K.; Hasan, M.M.; Kaya, O. Exogenous Nitric Oxide and Silicon Applications Alleviate Water Stress in Apricots. Life 2022, 12, 1454. https://doi.org/10.3390/life12091454
Bakır AG, Bolat I, Korkmaz K, Hasan MM, Kaya O. Exogenous Nitric Oxide and Silicon Applications Alleviate Water Stress in Apricots. Life. 2022; 12(9):1454. https://doi.org/10.3390/life12091454
Chicago/Turabian StyleBakır, Asuman Gundogdu, Ibrahim Bolat, Kubra Korkmaz, Md. Mahadi Hasan, and Ozkan Kaya. 2022. "Exogenous Nitric Oxide and Silicon Applications Alleviate Water Stress in Apricots" Life 12, no. 9: 1454. https://doi.org/10.3390/life12091454
APA StyleBakır, A. G., Bolat, I., Korkmaz, K., Hasan, M. M., & Kaya, O. (2022). Exogenous Nitric Oxide and Silicon Applications Alleviate Water Stress in Apricots. Life, 12(9), 1454. https://doi.org/10.3390/life12091454