


Outcomes of Low-Temperature Stress on Biological Alterations within Pothos (Epipremnum aureum) Leaves

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Chloroplast Pigment Contents Measurement
2.3. ROS Determination
2.4. Leaf Water Content Measurement
2.5. Relative Electrical Conductivity Measurement
2.6. Antioxidant Enzymatic Activities Determination
2.7. Chlorophyll Fluorescence Parameters Measurement
2.8. Anatomical Observation
2.9. RNA Isolation, cDNA Library Generation, and Sequencing
2.10. Sequence Assembly, Annotation, Identification, and Enrichment Analysis
2.11. qRT-PCR Validation
2.12. Statistical Analysis
3. Results
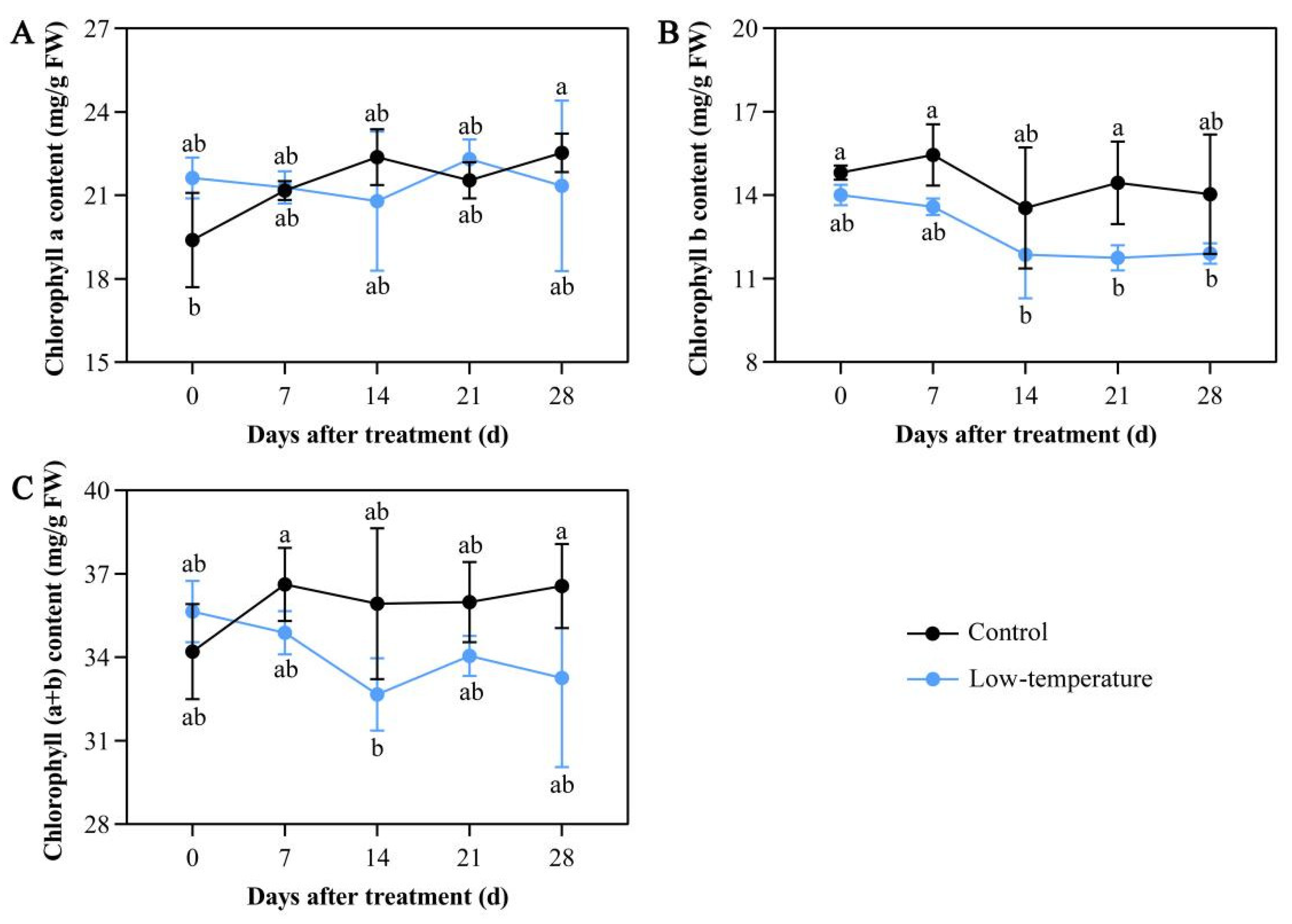
3.1. Influence of LTS on Chloroplast Pigment Concentrations
3.2. Influence of LTS on ROS Concentrations
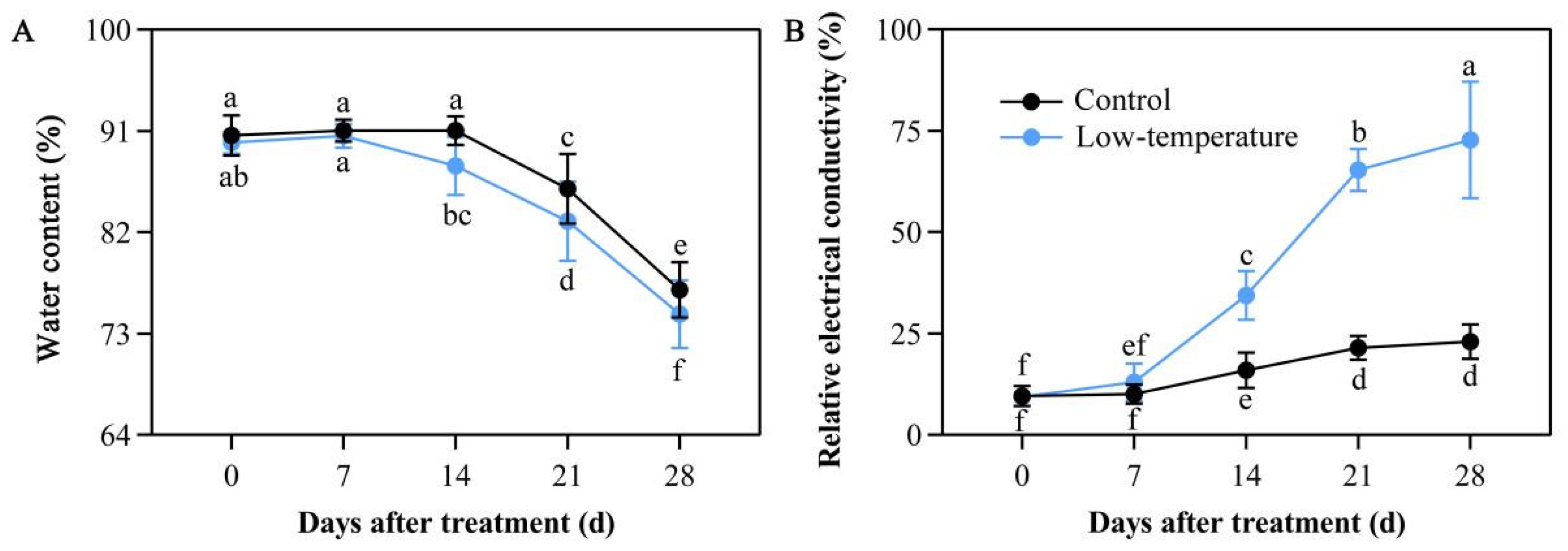
3.3. Influence of LTS on Leaf Water Content
3.4. Influence of LTS on Relative Electrical Conductivity
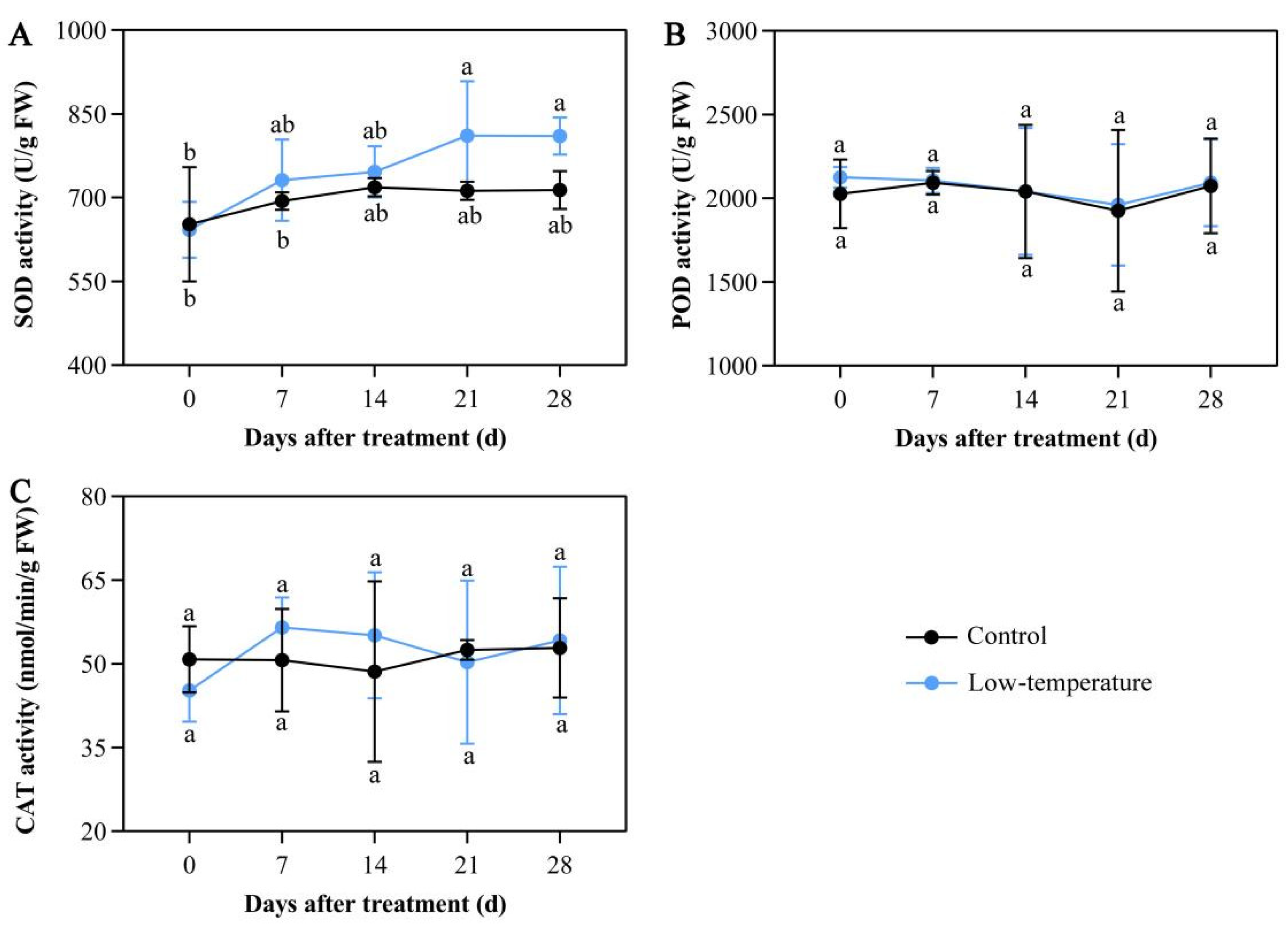
3.5. Influence of LTS on Antioxidant Enzymatic Activities
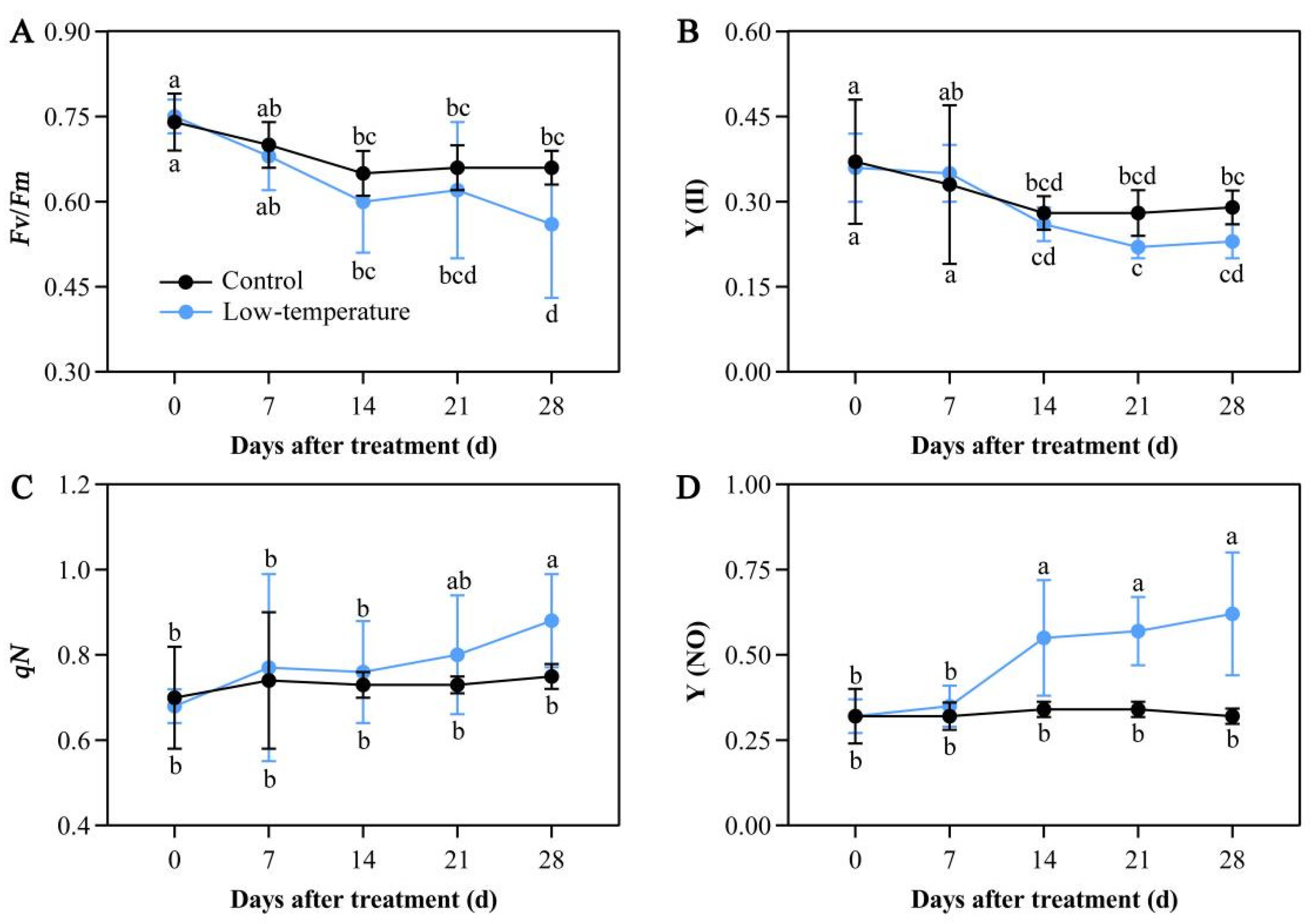
3.6. Influence of LTS on Chlorophyll Fluorescence Parameters
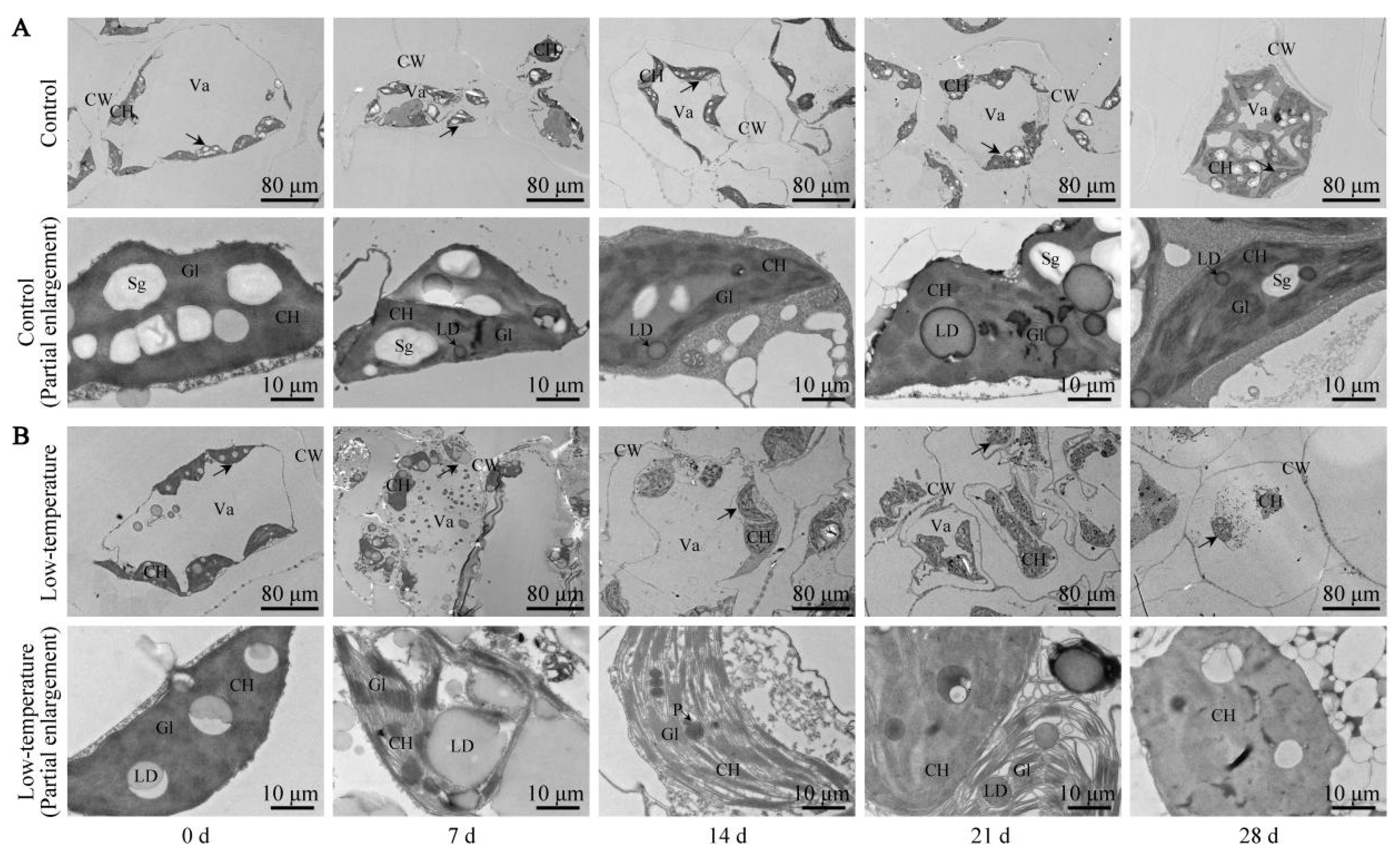
3.7. Influence of LTS on Leaf Ultrastructure
3.8. Mapping and Quantitative Evaluation of Illumina Sequences
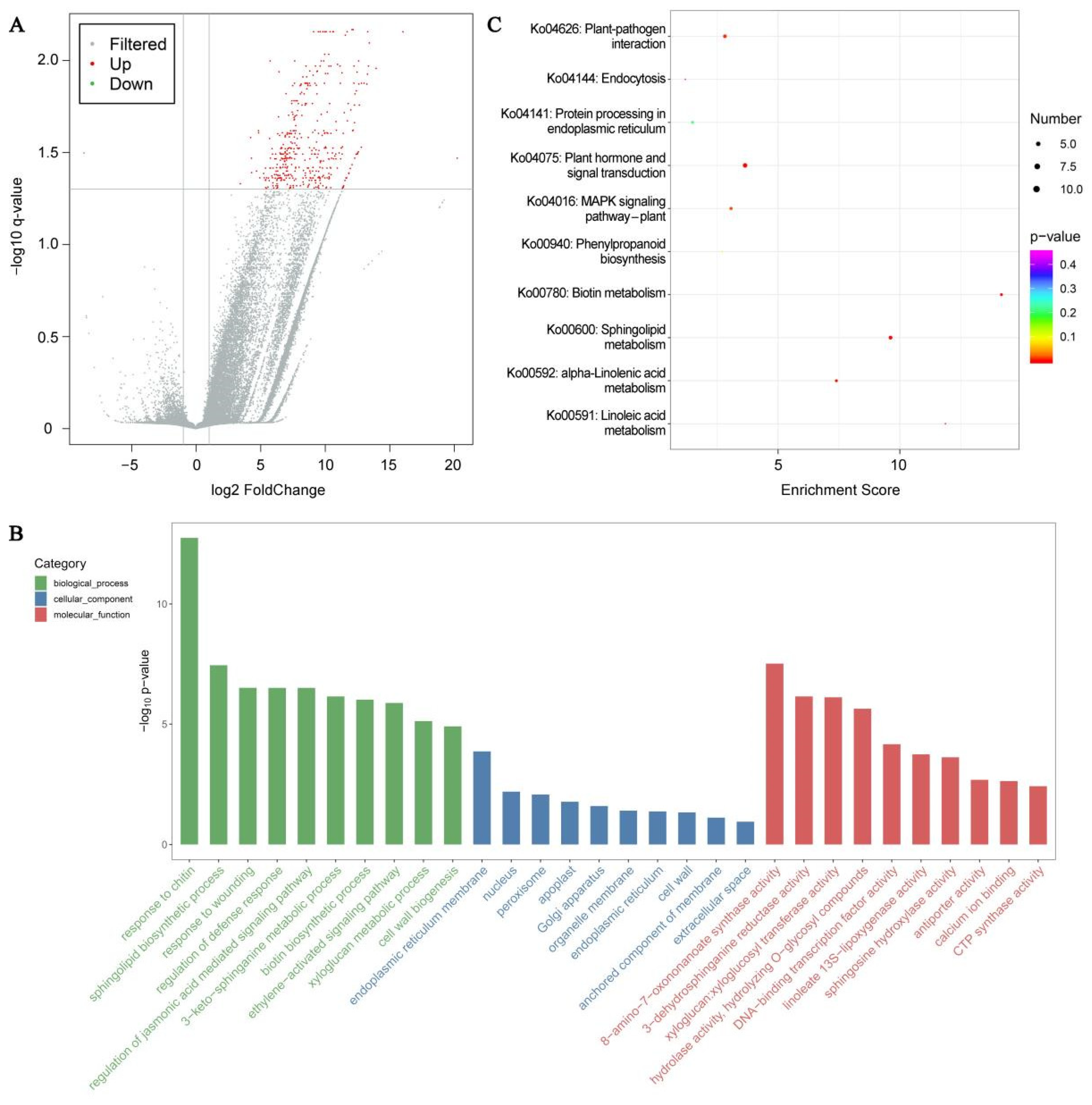
3.9. DEGs Identification and Enrichment Analyses
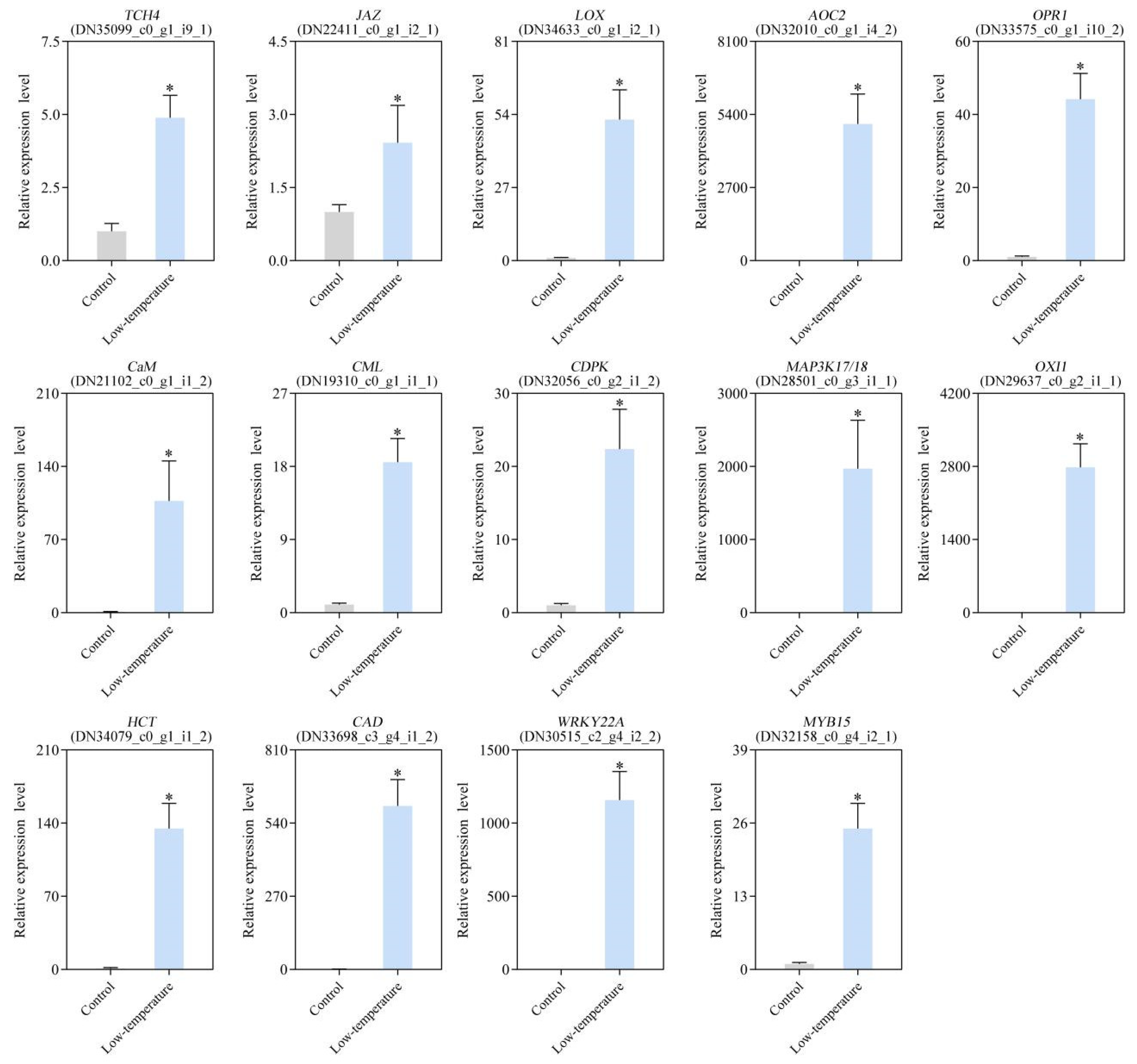
3.10. Verification of DEG Expression Profile via qRT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AOC2 | allene oxide cyclase |
| BP | biological process |
| BR | brassinosteroid |
| CAD | cinnamyl alcohol dehydrogenase |
| CaM | calmodulin |
| CAT | catalase |
| CDPK | calcium-dependent protein kinase |
| CH | chloroplast |
| CML | calmodulin-like protein |
| CW | cell wall |
| DAB | diaminobenzidine |
| DEGs | differentially expressed genes |
| DHE | dihydroethidium |
| DW | dry weight |
| Fm | maximum fluorescence |
| Fv | variable fluorescence |
| FW | fresh weight |
| Gl | basal lamellae |
| GO | gene ontology |
| HCT | hydroxycinnamoyl CoA: shikimate hydroxycinnamoyl transferase |
| JA | jasmonic acid |
| JAZ | jasmonate ZIM domain-containing protein |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| H2O2 | hydrogen peroxide |
| KOG | clusters of orthologous groups for eukaryotic complete genomes |
| LD | lipid droplet |
| LOX | lipoxygenase |
| LTS | low-temperature stress |
| MAP3K17/18 | mitogen-activated protein kinase kinase kinase 17/18 |
| NR | nonredundant |
| O2·− | superoxide radicals |
| OPR1 | 12-oxophytodienoate acid reductase |
| OXI1 | serine/threonine-protein kinase |
| P | plastid pellet |
| POD | peroxidase |
| qN | nonphotochemical quenching coefficient |
| ROS | reactive oxygen species |
| RT | room temperature |
| SD | standard deviation |
| Sg | starch granule |
| SOD | superoxide dismutase |
| SRA | sequence read archive |
| TCH4 | xyloglucan:xyloglucosyl transferase |
| TEM | transmission electron microscope |
| TF | transcription factors |
| V | volume |
| Va | central large vacuole |
| VIGS | virus-induced gene silencing |
| Y(II) | PSII photochemistry |
| Y(NO) | quantum yield of nonregulated energy dissipation |
References
- Walne, C.H.; Reddy, K.R. Temperature Effects on the Shoot and Root Growth, Development, and Biomass Accumulation of Corn (Zea mays L.). Agriculture 2022, 12, 443. [Google Scholar] [CrossRef]
- Gu, X.; Yang, C.; Zhao, H.; Hu, N.; Krauss, K.W.; Deng, C.; Chen, L. Sap flow evidence of chilling injury and recovery in mangroves following a spring cold spell. Trees 2021, 35, 907–917. [Google Scholar] [CrossRef]
- Szalai, G.; Majlath, I.; Pal, M.; Gondor, O.K.; Rundnoy, S.; Olah, C.; Vankova, R.; Kalapos, B.; Janda, T. Janus-Faced Nature of Light in the Cold Acclimation Processes of Maize. Front. Plant Sci. 2018, 9, 850. [Google Scholar] [CrossRef]
- Wei, C.W.; Huang, J.F.; Wang, X.Z.; Blackburn, G.A.; Zhang, Y.; Wang, S.S.; Mansaray, L.R. Hyperspectral characterization of freezing injury and its biochemical impacts in oilseed rape leaves. Remote Sens. Environ. 2017, 195, 56–66. [Google Scholar] [CrossRef]
- Zheng, G.; Li, L.; Li, W. Glycerolipidome responses to freezing-and chilling-induced injuries: Examples in Arabidopsis and rice. BMC Plant Biol. 2016, 16, 70. [Google Scholar] [CrossRef]
- Heidari, P.; Amerian, M.R.; Barcaccia, G. Hormone Profiles and Antioxidant Activity of Cultivated and Wild Tomato Seedlings under Low-Temperature Stress. Agronomy 2021, 11, 1146. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Pandit, E.; Nayak, D.K.; Behera, L.; Mohapatra, T. Genes, pathways and transcription factors involved in seedling stage chilling stress tolerance in indica rice through RNA-Seq analysis. BMC Plant Biol. 2019, 19, 352. [Google Scholar] [CrossRef]
- Shivashankar, K.S.; Geetha, G.A.; Ravishankar, K.V.; Rao, D.V.S.; Pavithra, K.C.G.; Roy, T.K. Temperature gradient storage induced biochemical and molecular changes in mango (Mangifera indica L. cv. Alphonso) fruits. Acta Physiol. Plant. 2022, 44, 46. [Google Scholar] [CrossRef]
- Chen, W.L.; KO, Y.T. Exogenous hydrogen peroxide induces chilling tolerance in Phalaenopsis seedlings through glutathione-related antioxidant system. Sci. Hortic. 2021, 289, 110421. [Google Scholar] [CrossRef]
- Hung, C.Y.; Qiu, J.; Sun, Y.H.; Chen, J.J.; Kittur, F.S.; Henny, R.J.; Jin, G.L.; Fan, L.J.; Xie, J.H. Gibberellin deficiency is responsible for shy-flowering nature of Epipremnum aureum. Sci. Rep. 2016, 6, 28598. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Sengupta, P.; Mustapha, M.S.; Sarker, M.R. An Experimental Evaluation of Adaptogenic Potential of Standardized Epipremnum Aureum Leaf Extract. J. Pharm. Bioallied Sci. 2016, 9, 88–93. [Google Scholar] [CrossRef]
- Shyamala, S.; Manikandan, N.A.; Pakshirajan, K.; Tang, V.T.; Rene, E.R.; Park, H.S.; Behera, S.K. Phytoremediation of nitrate contaminated water using ornamental plants. J. Water Supply Res. Technol. 2019, 68, 731–743. [Google Scholar] [CrossRef]
- Hou, H.; Xu, Z. Effect Of Benzene On Formaldehyde Removal By Shoots Of Three Indoor Plant Species. Environ. Eng. Manag. J. 2015, 14, 2849–2854. [Google Scholar]
- Lewis, D.H.; Arathoon, H.S.; Swinny, E.E.; Huang, S.C.; Funnell, K.A. Anthocyanin and carotenoid pigments in spathe tissue from selected Zantedeschia Hybrids. Acta Hortic. 2003, 624, 147–154. [Google Scholar] [CrossRef]
- Tian, F.; Gong, J.; Zhang, J.; Zhang, M.; Wang, G.; Li, A.; Wang, W. Enhanced stability of thylakoid membrane proteins and antioxidant competence contribute to drought stress resistance in the tasg1 wheat stay-green mutant. J. Exp. Bot. 2013, 64, 1509–1520. [Google Scholar] [CrossRef]
- Fang, Z.W.; Wang, X.X.; Zhang, X.Y.; Zhao, D.Q.; Tao, J. Effects of fulvic acid on the photosynthetic and physiological characteristics of Paeonia ostii under drought stress. Plant Signal. Behav. 2020, 15, 1774714. [Google Scholar] [CrossRef]
- Xu, J.; Jin, J.; Zhao, H.; Li, K. Drought stress tolerance analysis of Populus ussuriensis clones with different ploidies. J. For. Res. 2019, 30, 1267–1275. [Google Scholar] [CrossRef]
- Zhao, D.; Fang, Z.; Tang, Y.; Tao, J. Graphene Oxide as an Effective Soil Water Retention Agent Can Confer Drought Stress Tolerance to Paeonia ostii without Toxicity. Environ. Sci. Technol. 2020, 54, 8269–8279. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kanehisa, M.; Arak, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2019, 28, 511–515. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2013, 10, 71–73. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level—The DESeq package. Eur. Mol. Biol. Lab. 2012, 10, 1–23. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Korkmaz, A.; Deger, O.; Szafranska, K.; Koklu, S.; Karaca, A.; Yakupoglu, G.; Kocacinar, F. Melatonin effects in enhancing chilling stress tolerance of pepper. Sci. Hortic. 2021, 289, 110434. [Google Scholar] [CrossRef]
- Heidarvand, L.; Maali-Amiri, R. Physio-biochemical and proteome analysis of chickpea in early phases of cold stress. J. Plant Physiol. 2013, 170, 459–469. [Google Scholar] [CrossRef]
- Dong, J.; Zhao, J.; Zhang, S.; Yang, T.; Liu, Q.; Mao, X.; Fu, H.; Yang, W.; Liu, B. Physiological and genome-wide gene expression analyses of cold-induced leaf rolling at the seedling stage in rice (Oryza sativa L.). Crop J. 2019, 7, 431–443. [Google Scholar] [CrossRef]
- Erdal, S. Androsterone-induced molecular and physiological changes in maize seedlings in response to chilling stress. Plant Physiol. Biochem. 2012, 57, 1–7. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Shi, K.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q. Hydrogen peroxide is involved in the cold acclimation-induced chilling tolerance of tomato plants. Plant Physiol. Biochem. 2012, 60, 141–149. [Google Scholar] [CrossRef]
- John, R.; Anjum, N.A.; Sopory, S.K.; Akram, N.A.; Ashraf, M. Some key physiological and molecular processes of cold acclimation. Biol. Plant. 2016, 60, 603–618. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; EL-Khateeb, A.Y. Antioxidant enzyme activities and exogenous ascorbic acid treatment of ‘Williams’ banana during long-term cold storage stress. Sci. Hortic. 2018, 234, 210–219. [Google Scholar] [CrossRef]
- Esim, N.; Atici, O.; Mutlu, S. Effects of exogenous nitric oxide in wheat seedlings under chilling stress. Toxicol. Ind. Health 2014, 30, 268–274. [Google Scholar] [CrossRef]
- Wang, Y.X.; Hu, Y.; Chen, B.H.; Zhu, Y.F.; DaWuda, M.M.; Svetla, S. Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks. J. Integr. Agric. 2018, 17, 857–866. [Google Scholar] [CrossRef]
- Fu, J.; Miao, Y.; Shao, L.; Hu, T.; Yang, P. De novo transcriptome sequencing and gene expression profiling of Elymus nutans under cold stress. BMC Genom. 2016, 17, 870. [Google Scholar] [CrossRef]
- Popova, A.V.; Borisova, P.; Mihailova, G.; Georgieva, K. Antioxidative response of Arabidopsis thalianato combined action of low temperature and high light illumination when lutein is missing. Acta Physiol. Plant 2021, 44, 10. [Google Scholar] [CrossRef]
- Balabusta, M.; Szafranska, K.; Posmyk, M.M. Exogenous Melatonin Improves Antioxidant Defense in Cucumber Seeds (Cucumis sativus L.) Germinated under Chilling Stress. Front. Plant Sci. 2016, 7, 575. [Google Scholar] [CrossRef]
- Kaouthar, F.; Ameny, F.K.; Yosra, K.; Walid, S.; Ali, G.; Faical, B. Responses of transgenic Arabidopsis plants and recombinant yeast cells expressing a novel durum wheat manganese superoxide dismutase TdMnSOD to various abiotic stresses. J. Plant Physiol. 2016, 198, 56–68. [Google Scholar] [CrossRef]
- Ueda, Y.; Uehara, N.; Sasaki, H.; Kobayashi, K.; Yamakawa, T. Impacts of acute ozone stress on superoxide dismutase (SOD) expression and reactive oxygen species (ROS) formation in rice leaves. Plant Physiol. Biochem. 2013, 70, 396–402. [Google Scholar] [CrossRef]
- Che, Y.; Zhang, N.; Zhu, X.; Li, S.; Wang, S.; Si, H. Enhanced tolerance of the transgenic potato plants overexpressing Cu/Zn superoxide dismutase to low temperature. Sci. Hortic. 2020, 261, 108949. [Google Scholar] [CrossRef]
- Jan, N.; Majeed, U.; Andrabi, K.I.; John, R. Cold stress modulates osmolytes and antioxidant system in Calendula officinalis. Acta Physiol. Plant. 2018, 40, 73. [Google Scholar] [CrossRef]
- Sun, C.X.; Zhang, R.N.; Yuan, Z.Y.; Cao, H.X.; Martin, J.J.J. Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress. Horticulturae 2021, 7, 234. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, C.; Chen, J.; Qiu, J.; Huang, Z.; Wang, Q.; Ye, Y. Cold acclimation improves photosynthesis by regulating the ascorbate-glutathione cycle in chloroplasts of Kandelia obovata. J. For. Res. 2019, 30, 755–765. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence and Photosynthesis: The Basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.X.; Yu, Q.; Ma, Y.; Gu, W.R.; Yang, D.G. Physiological changes associated with enhanced cold resistance during maize (Zea mays) germination and seedling growth in response to exogenous calcium. Crop Pasture Sci. 2020, 71, 529–538. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Yang, Y.Q.; Wang, M.; Hu, S.J.; Wu, J.R.; Yu, Z.X. Differences in lipid homeostasis and membrane lipid unsaturation confer differential tolerance to low temperatures in two Cycas species. BMC Plant Biol. 2021, 21, 377. [Google Scholar] [CrossRef]
- Hola, D.; Kutik, J.; Kocova, M.; Rothova, O. Low-temperature induced changes in the ultrastructure of maize mesophyll chloroplasts strongly depend on the chilling pattern/intensity and considerably differ among inbred and hybrid genotypes. Photosynthetica 2008, 46, 329–338. [Google Scholar] [CrossRef]
- Zhang, F.; Lu, K.; Gu, Y.; Zhang, L.; Li, W.; Li, Z. Effects of low-temperature stress and brassinolide application on the photosynthesis and leaf structure of tung tree seedlings. Front. Plant Sci. 2020, 10, 1767. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 2003, 6, 410–417. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Zhou, J.; Gao, X.; Zhu, S.; Yuan, L.; Hou, X.; Liu, T.; Chen, G.; Tang, X.; et al. Transcriptome analysis and differential gene expression profiling of wucai (Brassica campestris L.) in response to cold stress. BMC Genom. 2022, 23, 137. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Ban, Q.; Zhu, X.; Jiang, C.; Wei, C.; Bennetzen, J.L. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genom. 2019, 20, 624. [Google Scholar] [CrossRef]
- Zhang, J.W.; Liang, L.; Xie, Y.D.; Zhao, Z.; Su, L.H.; Tang, Y.; Sun, B.; Lai, Y.S.; Li, H.X. Transcriptome and Metabolome Analyses Reveal Molecular Responses of Two Pepper (Capsicum annuum L.) Cultivars to Cold Stress. Front. Plant Sci. 2022, 13, 8196390. [Google Scholar] [CrossRef]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation—What is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef]
- Ye, H.Y.; Du, H.; Tang, N.; Li, X.H.; Xiong, L.Z. Identification and expression profiling analysis of TIFY family genes involved in stress and phytohormone responses in rice. Plant Mol. Biol. 2009, 71, 291–305. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, L.Q.; Wang, F.; Yu, D. Jasmonate Regulates the Inducer Of Cbf Expression-C-Repeat Binding Factor/Dre Binding Factor1 Cascade and Freezing Tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef]
- Sun, M.Z.; Shen, Y.; Chen, Y.; Wang, Y.; Cai, X.; Yang, J.; Jia, B.; Dong, W.; Chen, X.; Sun, X. Osa-miR1320 targets the ERF transcription factor OsERF096 to regulate cold tolerance via JA-mediated signaling. Plant Physiol. 2022, 189, 2500–2516. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Xu, L.; Singh, A.; Wang, H.; Du, L.; Poovaiah, B.W. Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 2015, 6, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, F.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Rodriguez, M.C.S.; Petersen, M.; Mundy, J. Mitogen-Activated Protein Kinase Signaling in Plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Boro, P.; Sultana, A.; Mandal, K.; Chattopadhyay, S. Interplay between glutathione and mitogen-activated protein kinase 3 via transcription factor WRKY40 under combined osmotic and cold stress in Arabidopsis. J. Plant Physiol. 2022, 271, 153664. [Google Scholar] [CrossRef]
- Lian, W.; Sun, T.; Meng, X.; Sun, R.; Hui, F.; Jiang, Y.; Zhao, Y. Overexpression of the Panax ginseng MYB4 gene enhances stress tolerance in transgenic Arabidopsis thaliana. Biol. Plant. 2021, 65, 27–38. [Google Scholar] [CrossRef]
- Wang, M.; Dai, W.S.; Du, J.; Ming, R.H.; Dahro, B.; Liu, J. ERF109 of trifoliate orange (Poncirus trifoliata (L.) Raf.) contributes to cold tolerance by directly regulating expression of Prx1 involved in antioxidative process. Plant Biotechnol. J. 2019, 17, 1316–1332. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene ID | Primer Pairs | Tm (°C) | Product Size (bp) |
|---|---|---|---|---|
| Actin | DN31460_c0_g1_i2_1 | F:AACTGCTCCTGAAAATCG R:CAACATCAATGACAACACCT | 48.8 | 152 |
| TCH4 | DN35099_c0_g1_i9_1 | F:CGTCACCGCCTACTACCTA R:TTGCTGTTCCCTATTCCC | 53.8 | 139 |
| JAZ | DN22411_c0_g1_i2_1 | F:GAGTTGTCTCCAGGATAAA R:GATTAGAGGGCTGAAGAG | 52.6 | 126 |
| LOX | DN34633_c0_g1_i2_1 | F:GCTCAACTTCGGGCAGTA R:AGTGCGTGGACAGCGTAT | 56.8 | 197 |
| AOC2 | DN32010_c0_g1_i4_2 | F:GACAGGTGAGGGGAAGGTA R:GGAATGTAACTCCAAGGAAA | 54.1 | 191 |
| OPR1 | DN33575_c0_g1_i10_2 | F:GGAGAAACACGGAAGGAG R:CAACCATACACCCCAAAA | 52.4 | 159 |
| CAM | DN21102_c0_g1_i1_2 | F:ATTCAGGAGGATGGCAGA R:CGAAAAGGCAATTCAAGTC | 53.9 | 193 |
| CML | DN19310_c0_g1_i1_1 | F:CACAAGCATCCACATCAG R:GCACCACCACGACTACTA | 50.8 | 115 |
| CDPK | DN32056_c0_g2_i1_2 | F:CGATGTTTTCCCTGTCTA R:CTTGGATGTTATGGGTCTAT | 46.6 | 106 |
| MAP3K17/18 | DN28501_c0_g3_i1_1 | F:ATCAGACATCACCACCGA R:CTGGTATTCCGTTCATCG | 53.0 | 123 |
| OXI1 | DN29637_c0_g2_i1_1 | F:TCTGATGCTTGTTGATTTCG R:AGTCGGAGTTTCCTTGCC | 54.3 | 179 |
| HCT | DN34079_c0_g1_i1_2 | F:GTGATAAGAGCACCCGAGA R:GCAACGCCACAATAGAGC | 53.9 | 105 |
| CAD | DN33698_c3_g4_i1_2 | F:TGTCAGTGCCATTGTAGG R:GATGTCACCAGGTTCTCG | 53.9 | 136 |
| WRKY22A | DN30515_c2_g4_i2_2 | F:AGACTCCCAAGTCCAAAA R:GCTGCATCTGTAATAACCC | 52.0 | 155 |
| MYB15 | DN32158_c0_g4_i2_1 | F:GCTGGTCTGTTGAGGTGT R:GTTCTTGATTTCGTTGTCTG | 52.4 | 189 |
| Sample | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Valid Bases | Q30 | GC |
|---|---|---|---|---|---|---|---|
| Control_1 | 50.04 M | 7.51 G | 48.97 M | 6.88 G | 91.70% | 94.73% | 51.52% |
| Control_2 | 48.37 M | 7.26 G | 47.37 M | 6.64 G | 91.48% | 94.83% | 51.15% |
| Control_3 | 48.46 M | 7.27 G | 47.43 M | 6.69 G | 91.99% | 94.77% | 51.46% |
| Low-temperature_1 | 47.10 M | 7.06 G | 45.89 M | 6.55 G | 92.70% | 94.34% | 51.75% |
| Low-temperature_2 | 51.73 M | 7.76 G | 50.55 M | 7.25 G | 93.38% | 94.47% | 51.85% |
| Low-temperature_3 | 50.08 M | 7.51 G | 48.79 M | 6.97 G | 92.81% | 94.32% | 51.48% |
| Database | NR | Swissprot | KEGG | KOG | eggNOG | GO | Pfam |
|---|---|---|---|---|---|---|---|
| Annotated number | 28,413 | 20,059 | 5839 | 15,842 | 25,290 | 17,295 | 17,087 |
| Percentage (%) | 42.6 | 30.08 | 8.75 | 23.75 | 37.92 | 25.93 | 25.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Cai, X.; Tang, Y. Outcomes of Low-Temperature Stress on Biological Alterations within Pothos (Epipremnum aureum) Leaves. Life 2022, 12, 1432. https://doi.org/10.3390/life12091432
Wu Y, Cai X, Tang Y. Outcomes of Low-Temperature Stress on Biological Alterations within Pothos (Epipremnum aureum) Leaves. Life. 2022; 12(9):1432. https://doi.org/10.3390/life12091432
Chicago/Turabian StyleWu, Yanqing, Xiang Cai, and Yuhan Tang. 2022. "Outcomes of Low-Temperature Stress on Biological Alterations within Pothos (Epipremnum aureum) Leaves" Life 12, no. 9: 1432. https://doi.org/10.3390/life12091432
APA StyleWu, Y., Cai, X., & Tang, Y. (2022). Outcomes of Low-Temperature Stress on Biological Alterations within Pothos (Epipremnum aureum) Leaves. Life, 12(9), 1432. https://doi.org/10.3390/life12091432