
Cyclosporine A Modulates LSP1 Protein Levels in Human B Cells to Attenuate B Cell Migration at Low O2 Levels

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Cell Lines
2.2. O Controlled Experiments
2.3. Chemotaxis Assay
2.4. HIF-1 and LSP1 Western Blots
2.5. Statistical Analysis of Chemotaxis and Western Experiments
2.6. shRNA Lentiviral Transfection
2.7. Phosphoproteomics
2.7.1. RAMOS Cell Sample Preparation
2.7.2. Protein Sample Preparation
2.7.3. Phosphopeptide Enrichment
2.7.4. Phosphopeptide Labeling
2.7.5. Peptide Fractionation
2.8. Data Acquisition and Processing
2.9. Differential Abundance
2.10. Gene Set Enrichment Analysis and Gene Ontology Analysis
2.11. Discrete-State Modeling with BONITA
3. Results
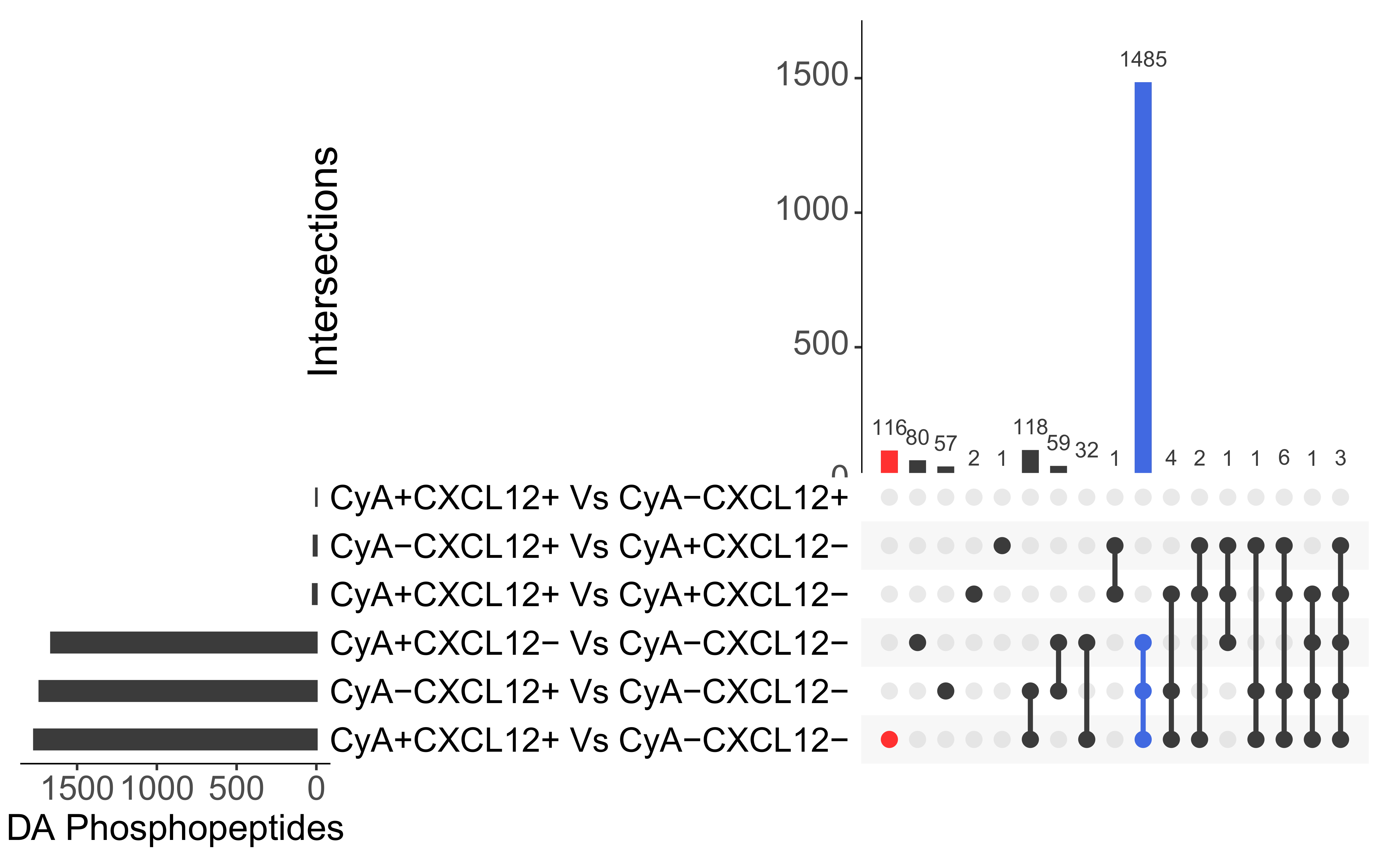
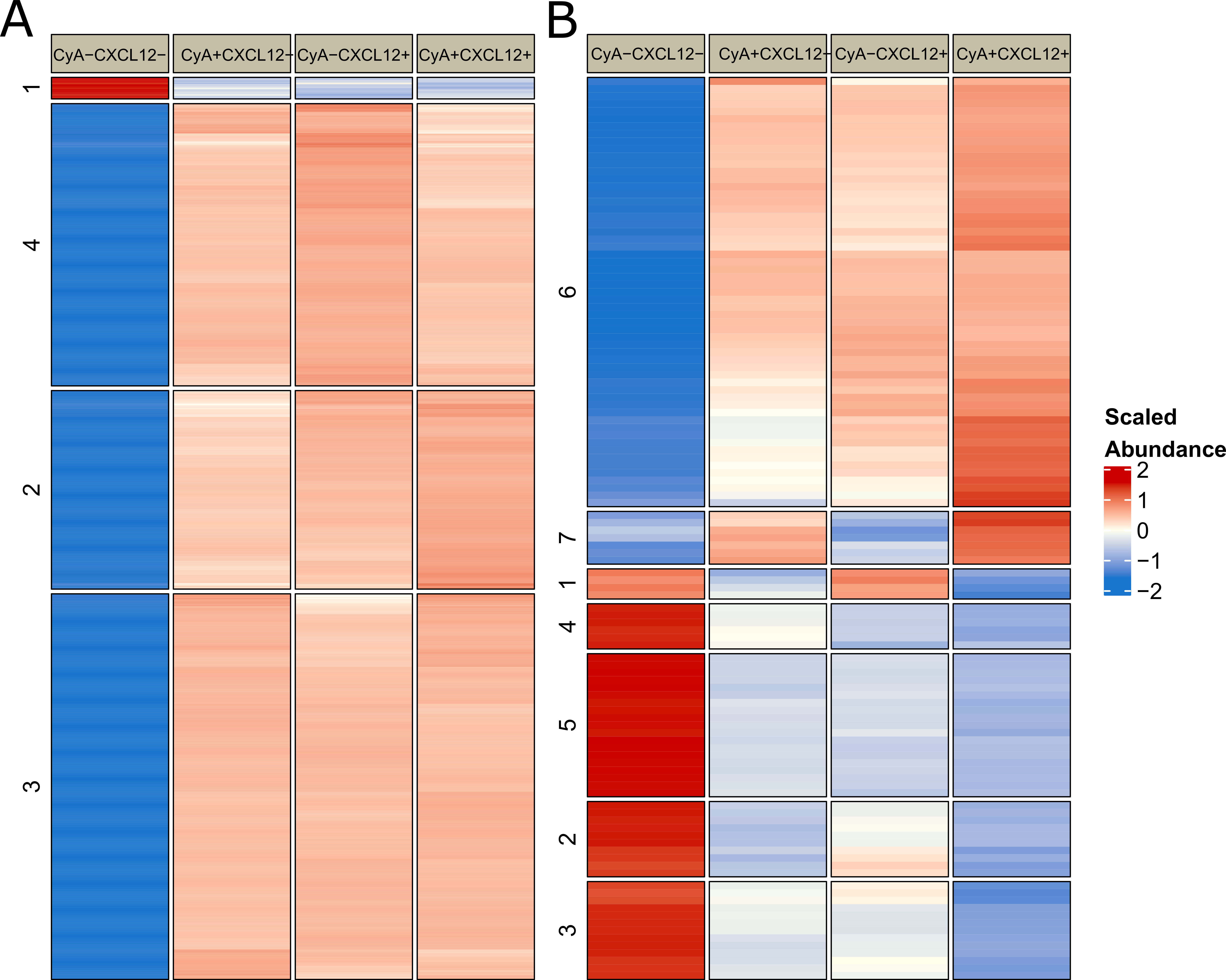
3.1. Initial Phosphoproteomic Analysis
3.2. CyclosporineA and CXCL12 Regulate Overlapping Signal Transduction Events
3.3. Kinase–Substrate Enrichment Analysis
3.4. Discrete-State Modeling with BONITA Links the HIF-1-Mediated Hypoxia Response to Remodeling of the Actin Cytoskeleton
3.5. LSP1 May Mediate Interaction between CyA and CXCL12
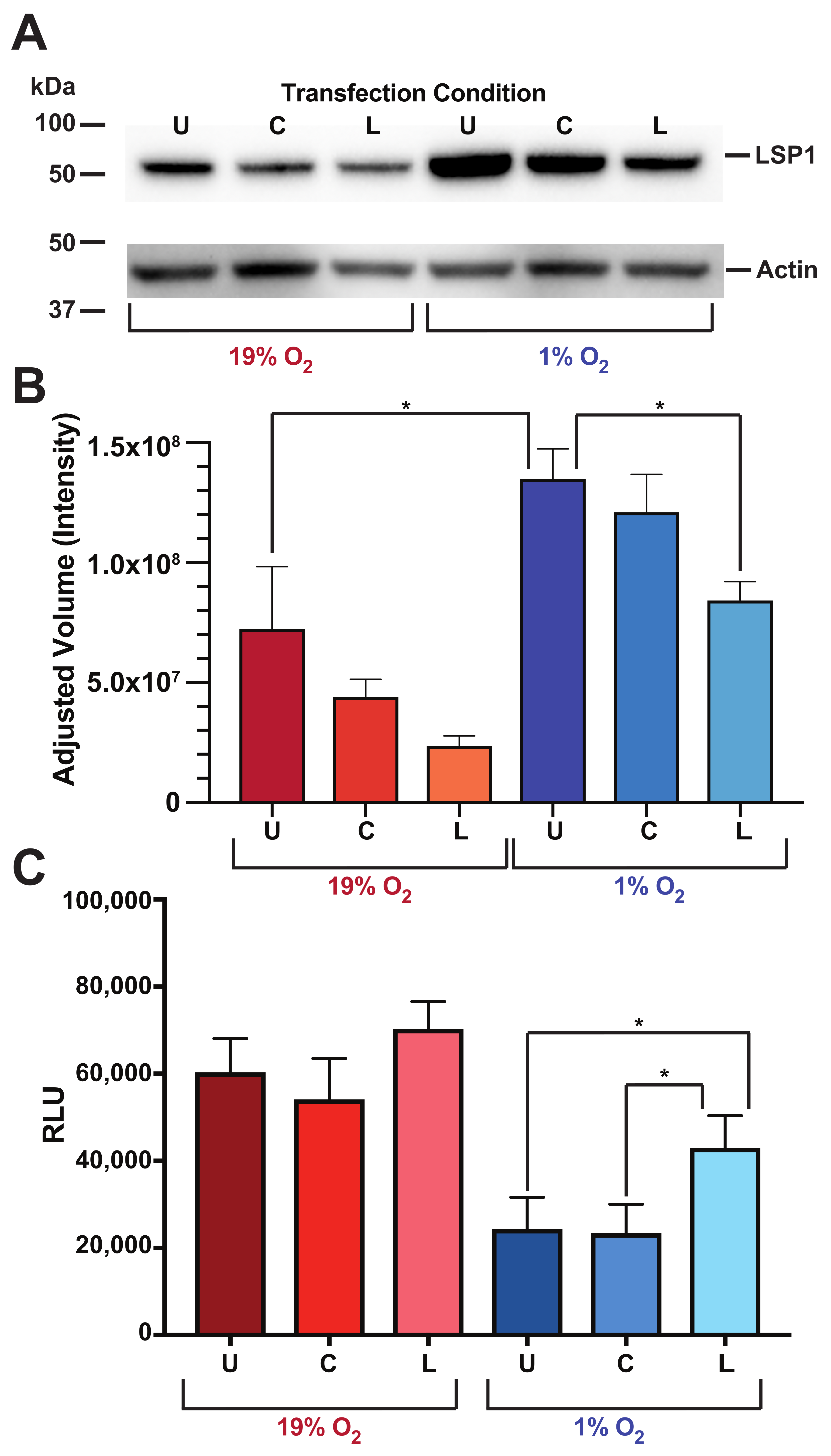
3.6. Human B Cell LSP1 Protein Amounts Are Increased at Low O Levels and Attenuated by CyA Treatment
3.7. LSP1 Protein Levels Directly Affect O-Dependent Chemokine Receptor Hypo-Responsiveness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carreau, A.; El Hafny-Rahbi, B.; Matejuk, A.; Grillon, C.; Kieda, C. Why is the partial oxygen pressure of human tissues a crucial parameter? Small molecules and hypoxia. J. Cell. Mol. Med. 2011, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shah, S.J.; Wilmes, L.J.; Feiner, J.; Kodibagkar, V.D.; Wendland, M.F.; Mason, R.P.; Hylton, N.; Hopf, H.W.; Rollins, M.D. Quantitative tissue oxygen measurement in multiple organs using 19F MRI in a rat model. Magn. Reson. Med. 2011, 66, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- Hilchey, S.; Palshikar, M.; Li, D.; Garigen, J.; Cipolla, V.; Thakar, J.; Zand, M. Cyclosporine A Directly Affects Human and Mouse B cell Migration in vitro by Disrupting a HIF-1α Dependent, O2 Sensing, Molecular Switch. BMC Immunol. 2019, 21, 1–18. [Google Scholar] [CrossRef]
- Cho, S.H.; Raybuck, A.L.; Stengel, K.; Wei, M.; Beck, T.C.; Volanakis, E.; Thomas, J.W.; Hiebert, S.; Haase, V.H.; Boothby, M.R. Germinal centre hypoxia and regulation of antibody qualities by a hypoxia response system. Nature 2016, 537, 234–238. [Google Scholar] [CrossRef]
- Jellusova, J.; Cato, M.H.; Apgar, J.R.; Ramezani-Rad, P.; Leung, C.R.; Chen, C.; Richardson, A.D.; Conner, E.M.; Benschop, R.J.; Woodgett, J.R.; et al. Gsk3 is a metabolic checkpoint regulator in B cells. Nat. Immunol. 2017, 18, 303–312. [Google Scholar] [CrossRef]
- Abbott, R.K.; Thayer, M.; Labuda, J.; Silva, M.; Philbrook, P.; Cain, D.W.; Kojima, H.; Hatfield, S.; Sethumadhavan, S.; Ohta, A.; et al. Germinal Center Hypoxia Potentiates Immunoglobulin Class Switch Recombination. J. Immunol. 2016, 197, 4014–4020. [Google Scholar] [CrossRef]
- Bannard, O.; Horton, R.M.; Allen, C.D.; An, J.; Nagasawa, T.; Cyster, J.G. Germinal center centroblasts transition to a centrocyte phenotype according to a timed program and depend on the dark zone for effective selection. Immunity 2013, 39, 912–924. [Google Scholar] [CrossRef]
- Malat, G.; Culkin, C. The ABCs of Immunosuppression: A Primer for Primary Care Physicians. Med. Clin. North Am. 2016, 100, 505–518. [Google Scholar] [CrossRef]
- Florescu, D.F.; Kalil, A.C.; Qiu, F.; Schmidt, C.M.; Sandkovsky, U. What is the impact of hypogammaglobulinemia on the rate of infections and survival in solid organ transplantation? A meta-analysis. Am. J. Transplant. 2013, 13, 2601–2610. [Google Scholar] [CrossRef]
- Felldin, M.; Andersson, B.; Studahl, M.; Svennerholm, B.; Friman, V. Antibody persistence 1 year after pandemic H1N1 2009 influenza vaccination and immunogenicity of subsequent seasonal influenza vaccine among adult organ transplant patients. Transpl. Int. 2014, 27, 197–203. [Google Scholar] [CrossRef]
- Felldin, M.; Studahl, M.; Svennerholm, B.; Friman, V. The antibody response to pandemic H1N1 2009 influenza vaccine in adult organ transplant patients. Transpl. Int. 2012, 25, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Heidt, S.; Roelen, D.L.; Eijsink, C.; Eikmans, M.; van Kooten, C.; Claas, F.H.; Mulder, A. Calcineurin inhibitors affect B cell antibody responses indirectly by interfering with T cell help. Clin. Exp. Immunol. 2010, 159, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Vargas, P.; Barbier, L.; Saez, P.J.; Piel, M. Mechanisms for fast cell migration in complex environments. Curr. Opin. Cell Biol. 2017, 48, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Maity, P.C.; Datta, M.; Nicolo, A.; Jumaa, H. Isotype Specific Assembly of B Cell Antigen Receptors and Synergism with Chemokine Receptor CXCR4. Front Immunol. 2018, 9, 2988. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J.; Wood, W. Unravelling the Actin Cytoskeleton: A New Competitive Edge? Trends Cell Biol. 2016, 26, 569–576. [Google Scholar] [CrossRef]
- Moulding, D.A.; Record, J.; Malinova, D.; Thrasher, A.J. Actin cytoskeletal defects in immunodeficiency. Immunol. Rev. 2013, 256, 282–299. [Google Scholar] [CrossRef]
- Howard, T.H.; Hartwig, J.; Cunningham, C. Lymphocyte-specific protein 1 expression in eukaryotic cells reproduces the morphologic and motile abnormality of NAD 47/89 neutrophils. Blood 1998, 91, 4786–4795. [Google Scholar] [CrossRef]
- Coates, T.D.; Torkildson, J.C.; Torres, M.; Church, J.A.; Howard, T.H. An inherited defect of neutrophil motility and microfilamentous cytoskeleton associated with abnormalities in 47-Kd and 89-Kd proteins. Blood 1991, 78, 1338–1346. [Google Scholar] [CrossRef]
- Jongstra, J.; Tidmarsh, G.F.; Jongstra-Bilen, J.; Davis, M.M. A new lymphocyte-specific gene which encodes a putative Ca2+-binding protein is not expressed in transformed T lymphocyte lines. J. Immunol. 1988, 141, 3999–4004. [Google Scholar]
- Jongstra-Bilen, J.; Janmey, P.A.; Hartwig, J.H.; Galea, S.; Jongstra, J. The lymphocyte-specific protein LSP1 binds to F-actin and to the cytoskeleton through its COOH-terminal basic domain. J. Cell Biol. 1992, 118, 1443–1453. [Google Scholar] [CrossRef]
- Zieseniss, A. Hypoxia and the modulation of the actin cytoskeleton-emerging interrelations. Hypoxia 2014, 2, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Palli, R.; Palshikar, M.G.; Thakar, J. Executable pathway analysis using ensemble discrete-state modeling for large-scale data. PLoS Comput. Biol. 2019, 15, e1007317. [Google Scholar] [CrossRef]
- Shen, S.; An, B.; Wang, X.; Hilchey, S.P.; Li, J.; Cao, J.; Tian, Y.; Hu, C.; Jin, L.; Ng, A.; et al. Surfactant Cocktail-Aided Extraction/Precipitation/On-Pellet Digestion Strategy Enables Efficient and Reproducible Sample Preparation for Large-Scale Quantitative Proteomics. Anal. Chem. 2018, 90, 10350–10359. [Google Scholar] [CrossRef] [PubMed]
- Paulo, J.A.; Gaun, A.; Gygi, S.P. Global Analysis of Protein Expression and Phosphorylation Levels in Nicotine-Treated Pancreatic Stellate Cells. J. Proteome. Res. 2015, 14, 4246–4256. [Google Scholar] [CrossRef] [PubMed]
- Erickson, B.K.; Jedrychowski, M.P.; McAlister, G.C.; Everley, R.A.; Kunz, R.; Gygi, S.P. Evaluating multiplexed quantitative phosphopeptide analysis on a hybrid quadrupole mass filter/linear ion trap/orbitrap mass spectrometer. Anal. Chem. 2015, 87, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [CrossRef]
- Barker, B.L.; Benovic, J.L. G protein-coupled receptor kinase 5 phosphorylation of hip regulates internalization of the chemokine receptor CXCR4. Biochemistry 2011, 50, 6933–6941. [Google Scholar] [CrossRef] [PubMed]
- Dayam, R.M.; Sun, C.X.; Choy, C.H.; Mancuso, G.; Glogauer, M.; Botelho, R.J. The Lipid Kinase PIKfyve Coordinates the Neutrophil Immune Response through the Activation of the Rac GTPase. J. Immunol. 2017, 199, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Forster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef]
- Dang, E.V.; Barbi, J.; Yang, H.Y.; Jinasena, D.; Yu, H.; Zheng, Y.; Bordman, Z.; Fu, J.; Kim, Y.; Yen, H.R.; et al. Control of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell 2011, 146, 772–784. [Google Scholar] [CrossRef]
- Doedens, A.L.; Phan, A.T.; Stradner, M.H.; Fujimoto, J.K.; Nguyen, J.V.; Yang, E.; Johnson, R.S.; Goldrath, A.W. Hypoxia-inducible factors enhance the effector responses of CD8(+) T cells to persistent antigen. Nat. Immunol. 2013, 14, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, S.R.; Print, C.; Farahi, N.; Peyssonnaux, C.; Johnson, R.S.; Cramer, T.; Sobolewski, A.; Condliffe, A.M.; Cowburn, A.S.; Johnson, N.; et al. Hypoxia-induced neutrophil survival is mediated by HIF-1alpha-dependent NF-kappaB activity. J. Exp. Med. 2005, 201, 105–115. [Google Scholar] [CrossRef]
- Burrows, N.; Maxwell, P.H. Hypoxia and B cells. Exp. Cell Res. 2017, 356, 197–203. [Google Scholar] [CrossRef]
- Bleul, C.C.; Schultze, J.L.; Springer, T.A. B Lymphocyte Chemotaxis Regulated in Association with Microanatomic Localization, Differentiation State, and B Cell Receptor Engagement. J. Exp. Med. 1998, 187, 753–762. [Google Scholar] [CrossRef]
- Bowman, E.P.; Campbell, J.J.; Soler, D.; Dong, Z.; Manlongat, N.; Picarella, D.; Hardy, R.R.; Butcher, E.C. Developmental Switches in Chemokine Response Profiles during B Cell Differentiation and Maturation. J. Exp. Med. 2000, 191, 1303–1318. [Google Scholar] [CrossRef]
- Melchers, F.; Rolink, A.G.; Schaniel, C. The Role of Chemokines in Regulating Cell Migration during Humoral Immune Responses. Cell 1999, 99, 351–354. [Google Scholar] [CrossRef]
- De Silva, N.S.; Klein, U. Dynamics of B cells in germinal centres. Nat. Rev. Immunol. 2015, 15, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Jung, S.H.; Lee, S.; Choi, S.; Yoo, S.A.; Park, J.H.; Hwang, D.; Shim, S.C.; Sabbagh, L.; Kim, K.J.; et al. Leukocyte-specific protein 1 regulates T-cell migration in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2015, 112, E6535–E6543. [Google Scholar] [CrossRef] [PubMed]
- Jongstra-Bilen, J.; Misener, V.L.; Wang, C.; Ginzberg, H.; Auerbach, A.; Joyner, A.L.; Downey, G.P.; Jongstra, J. LSP1 modulates leukocyte populations in resting and inflamed peritoneum. Blood 2000, 96, 1827–1835. [Google Scholar] [CrossRef]
- Harrison, R.E.; Sikorski, B.A.; Jongstra, J. Leukocyte-specific protein 1 targets the ERK/MAP kinase scaffold protein KSR and MEK1 and ERK2 to the actin cytoskeleton. J. Cell Sci. 2004, 117, 2151–2157. [Google Scholar] [CrossRef]
- Wu, J.L.; Wu, H.Y.; Tsai, D.Y.; Chiang, M.F.; Chen, Y.J.; Gao, S.; Lin, C.C.; Lin, C.H.; Khoo, K.H.; Chen, Y.J.; et al. Temporal regulation of Lsp1 O-GlcNAcylation and phosphorylation during apoptosis of activated B cells. Nat. Commun. 2016, 7, 12526. [Google Scholar] [CrossRef]
- Haerzschel, A.; Catusse, J.; Hutterer, E.; Paunovic, M.; Zirlik, K.; Eibel, H.; Krenn, P.W.; Hartmann, T.N.; Burger, M. BCR and chemokine responses upon anti-IgM and anti-IgD stimulation in chronic lymphocytic leukaemia. Ann. Hematol. 2016, 95, 1979–1988. [Google Scholar] [CrossRef]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Immunogenicity of a Single Dose of SARS-CoV-2 Messenger RNA Vaccine in Solid Organ Transplant Recipients. JAMA 2021, 325, 1784–1786. [Google Scholar] [CrossRef]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Antibody Response to 2-Dose SARS-CoV-2 mRNA Vaccine Series in Solid Organ Transplant Recipients. JAMA 2021, 325, 2204–2206. [Google Scholar] [CrossRef]
- Grupper, A.; Katchman, H. Reduced humoral response to mRNA SARS-CoV-2 BNT162b2 vaccine in kidney transplant recipients without prior exposure to the virus: Not alarming, but should be taken gravely. Am. J. Transplant. 2021, 21, 2909. [Google Scholar] [CrossRef]
- Shostak, Y.; Shafran, N.; Heching, M.; Rosengarten, D.; Shtraichman, O.; Shitenberg, D.; Amor, S.M.; Yahav, D.; Ben Zvi, H.; Pertzov, B.; et al. Early humoral response among lung transplant recipients vaccinated with BNT162b2 vaccine. Lancet Respir. Med. 2021, 9, e52–e53. [Google Scholar] [CrossRef]
- Heeger, P.S.; Larsen, C.P.; Segev, D.L. Implications of defective immune responses in SARS-CoV-2 vaccinated organ transplant recipients. Sci. Immunol. 2021, 6, eabj6513. [Google Scholar] [CrossRef] [PubMed]
- Embi, P.J.; Levy, M.E.; Naleway, A.L.; Patel, P.; Gaglani, M.; Natarajan, K.; Dascomb, K.; Ong, T.C.; Klein, N.P.; Liao, I.C.; et al. Effectiveness of 2-Dose Vaccination with mRNA COVID-19 Vaccines Against COVID-19-Associated Hospitalizations Among Immunocompromised Adults-Nine States, January-September 2021. MMWR Morb. Mortal. Wkly Rep. 2021, 70, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway | KEGG Code |
|---|---|
| MAPK signaling | hsa04010 |
| Chemokine signaling | hsa04062 |
| NF-kappa B signaling | hsa04064 |
| HIF-1 signaling | hsa04066 |
| mTOR signaling | hsa04150 |
| PI3K-Akt signaling | hsa04151 |
| VEGF signaling | hsa04370 |
| Cell adhesion | hsa04514 |
| C-type lectin receptor signaling | hsa04625 |
| JAK-STAT signaling | hsa04630 |
| TNF signaling | hsa04668 |
| Leukocyte transendothelial migration | hsa04670 |
| Regulation of actin cytoskeleton | hsa04810 |
| Node | Importance Score from BONITA |
|---|---|
| GRK5 | 1.00 |
| CXCR4 | 0.83 |
| SYK | 0.63 |
| STAT3 | 0.51 |
| PLCG2 | 0.38 |
| CD19 | 0.34 |
| PDPK1 | 0.34 |
| BLNK | 0.33 |
| MTOR | 0.30 |
| CSNK2B | 0.29 |
| PRKCG | 0.27 |
| PHLPP1 | 0.27 |
| MAP2K2 | 0.27 |
| LSP1 | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilchey, S.P.; Palshikar, M.G.; Mendelson, E.S.; Shen, S.; Rasam, S.; Emo, J.A.; Qu, J.; Thakar, J.; Zand, M.S. Cyclosporine A Modulates LSP1 Protein Levels in Human B Cells to Attenuate B Cell Migration at Low O2 Levels. Life 2022, 12, 1284. https://doi.org/10.3390/life12081284
Hilchey SP, Palshikar MG, Mendelson ES, Shen S, Rasam S, Emo JA, Qu J, Thakar J, Zand MS. Cyclosporine A Modulates LSP1 Protein Levels in Human B Cells to Attenuate B Cell Migration at Low O2 Levels. Life. 2022; 12(8):1284. https://doi.org/10.3390/life12081284
Chicago/Turabian StyleHilchey, Shannon P., Mukta G. Palshikar, Eric S. Mendelson, Shichen Shen, Sailee Rasam, Jason A. Emo, Jun Qu, Juilee Thakar, and Martin S. Zand. 2022. "Cyclosporine A Modulates LSP1 Protein Levels in Human B Cells to Attenuate B Cell Migration at Low O2 Levels" Life 12, no. 8: 1284. https://doi.org/10.3390/life12081284