
How Is CYP17A1 Activity Altered in Autism? A Pilot Study to Identify Potential Pharmacological Targets


Abstract
:1. Introduction
2. Material & Methods
2.1. Participants
2.2. Study Procedure
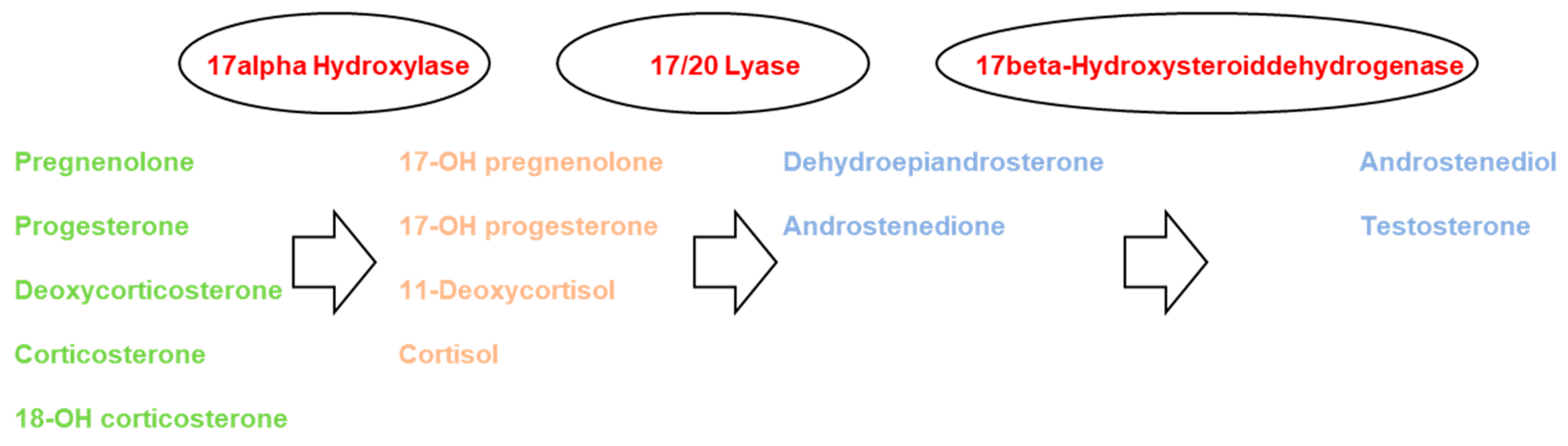
3. Methods
4. Statistical Analysis
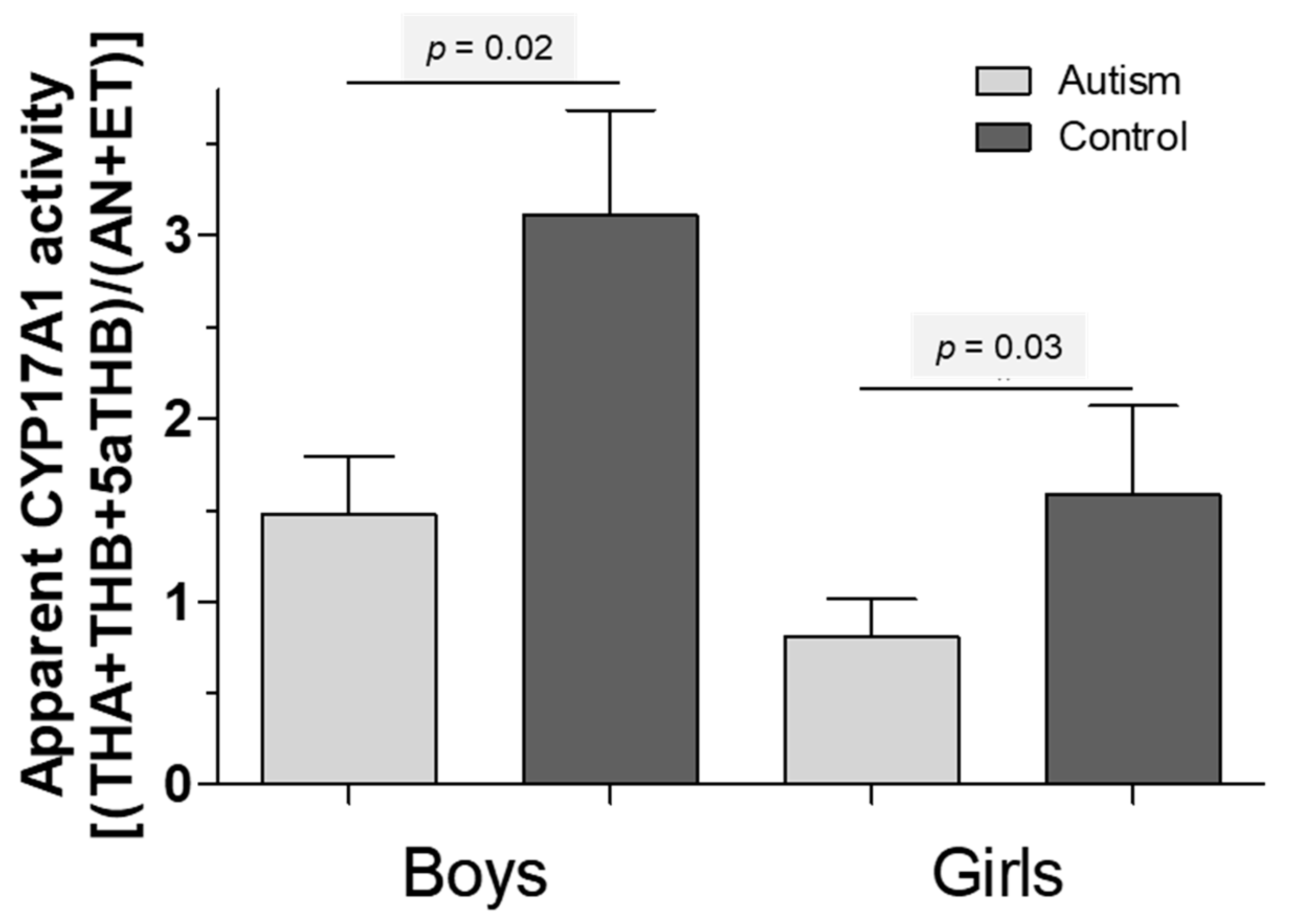
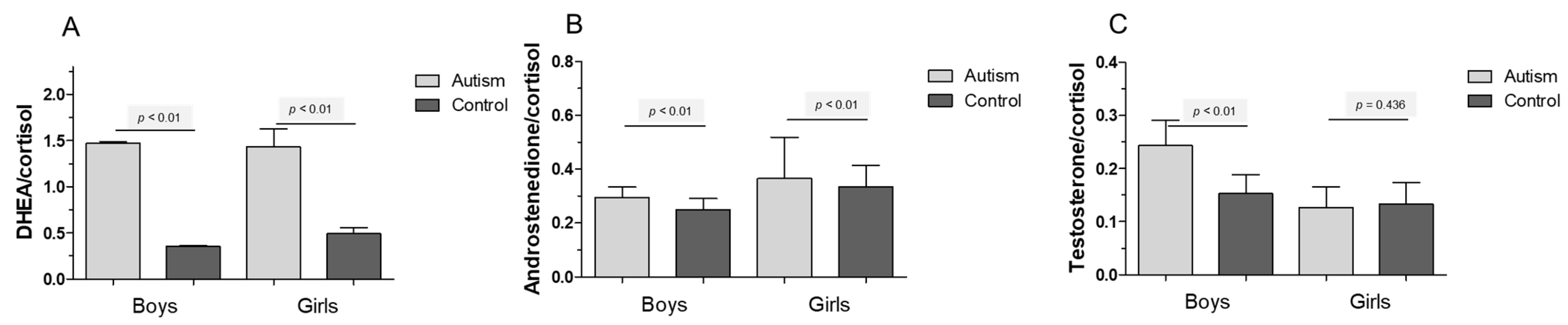
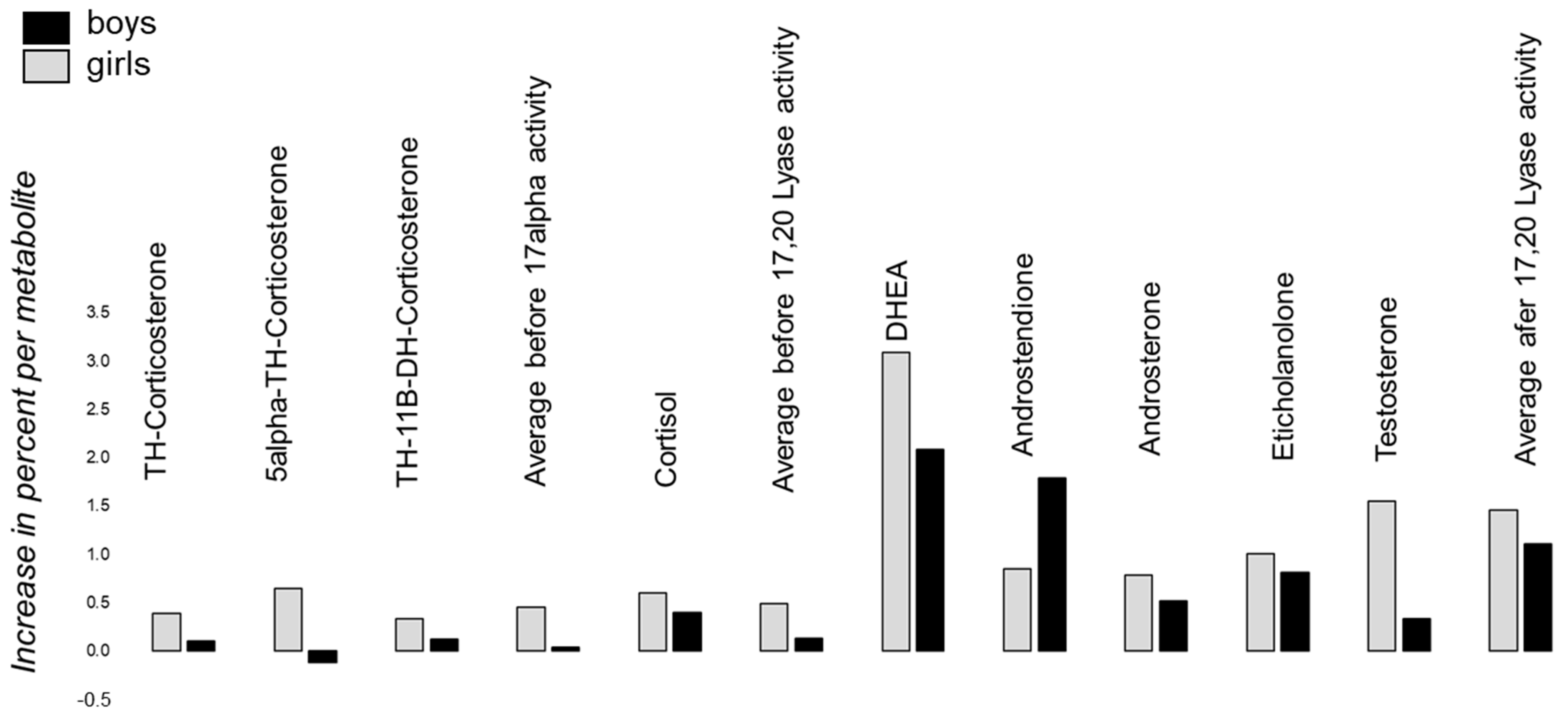
5. Results
6. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asperger, H. Die Autistischen Psychopathen im Kindesalter. Arch. Für Psychiatr. Und Nervenkrankh. 1944, 117, 76–136. [Google Scholar] [CrossRef]
- Gasser, B. The Case of Hellmuth in The Autistic Psychopathy—Suffering from Cushing Syndrome? Glob. J. Intellect. Dev. Disabil. 2018, 4, 555643. [Google Scholar] [CrossRef]
- Taylor, J.L.; Corbett, B.A. A review of rhythm and responsiveness of cortisol in individuals with autism spectrum disorders. Psychoneuroendocrinology 2014, 49, 207–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron Cohen, S.; Auyeung, B.; Norgaard Pedersen, B.; Hougaard, D.M.; Abdallah, M.W.; Melgaard, L.; Lombardo, M.V. Elevated fetal steroidogenic activity in autism. Mol. Psychiatry 2015, 20, 369–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillberg, C.; Fernell, E.; Kočovská, E.; Minnis, H.; Bourgeron, T.; Thompson, L.; Allely, C. The role of cholesterol metabolism and various steroid abnormalities in autism spectrum disorders: A hypothesis paper. Autism Res. 2017, 10, 1022–1044. [Google Scholar] [CrossRef]
- Niu, M.; Han, Y.; Dy, A.B.C.; Du, J.; Jin, H.; Qin, J.; Zhang, J.; Li, Q.; Hagerman, R.J. Autism Symptoms in Fragile X Syndrome. J. Child Neurol. 2017, 32, 903–909. [Google Scholar] [CrossRef]
- Sarachana, T.; Hu, V.W. Differential recruitment of coregulators to the RORA promoter adds another layer of complexity to gene (dys) regulation by sex hormones in autism. Mol. Autism 2013, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Sarachana, T.; Xu, M.; Wu, R.C.; Hu, V.W. Sex hormones in autism: Androgens and estrogens differentially and reciprocally regulate RORA, a novel candidate gene for autism. PLoS ONE 2011, 6, e17116. [Google Scholar] [CrossRef]
- Porubek, D. CYP17A1: A biochemistry, chemistry, and clinical review. Curr. Top. Med. Chem. 2013, 13, 1364–1384. [Google Scholar] [CrossRef]
- Hu, V.W.; Sarachana, T.; Sherrard, R.M.; Kocher, K.M. Investigation of sex differences in the expression of RORA and its transcriptional targets in the brain as a potential contributor to the sex bias in autism. Mol. Autism 2015, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Hu, V.W.; Nguyen, A.; Kim, K.S.; Steinberg, M.E.; Sarachana, T.; Scully, M.A.; Soldin, S.J.; Luu, T.; Lee, N.H. Gene expression profiling of lymphoblasts from autistic and nonaffected sib pairs: Altered pathways in neuronal development and steroid biosynthesis. PLoS ONE 2009, 4, e5775. [Google Scholar] [CrossRef] [PubMed]
- Gasser, B.A.; Kurz, J.; Dick, B.; Mohaupt, M.G. Are Steroid Hormones Dysregulated in Autistic Girls? Diseases 2020, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, B.A.; Kurz, J.; Dick, B.; Mohaupt, M.G. Steroid Metabolites Support Evidence of Autism as a Spectrum. Behav. Sci. 2019, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janšáková, K.; Hill, M.; Čelárová, D.; Celušáková, H.; Repiská, G.; Bičíková, M.; Máčová, L.; Ostatníková, D. Alteration of the steroidogenesis in boys with autism spectrum disorders. Transl. Psychiatry 2020, 10, 340. [Google Scholar] [CrossRef]
- Gasser, B.A.; Buerki, S.F.; Kurz, J.; Mohaupt, M.G. Hyperandrogenism? Increased 17, 20-Lyase Activity? A Metanalysis and Systematic Review of Altered Androgens in Boys and Girls with Autism. Int. J. Mol. Sci. 2021, 22, 12324. [Google Scholar] [CrossRef]
- Miller, W.L.; Tee, M.K. The post-translational regulation of 17,20 lyase activity. Mol. Cell Endocrinol. 2015, 408, 99–106. [Google Scholar] [CrossRef]
- Sun, M.; Mueller, J.W.; Gilligan, L.C.; Taylor, A.E.; Shaheen, F.; Noczyńska, A.; T’Sjoen, G.; Denvir, L.; Shenoy, S.; Fulton, P.; et al. The broad phenotypic spectrum of 17α-hydroxylase/17,20-lyase (CYP17A1) deficiency: A case series. Eur. J. Endocrinol. 2021, 185, 729–741. [Google Scholar] [CrossRef]
- Rubtsov, P.; Nizhnik, A.; Dedov, I.; Kalinchenko, N.; Petrov, V.; Orekhova, A.; Spirin, P.; Prassolov, V.; Tiulpakov, A. Partial deficiency of 17α-hydroxylase/17,20-lyase caused by a novel missense mutation in the canonical cytochrome heme-interacting motif. Eur. J. Endocrinol. 2015, 172, K19–K25. [Google Scholar] [CrossRef] [Green Version]
- Yanase, T.; Simpson, E.R.; Waterman, M.R. 17Alpha-hydroxylase/17,20-lyase deficiency: From clinical investigation to molecular definition. Endocr. Rev. 1991, 12, 91–108. [Google Scholar] [CrossRef]
- Taniyama, M.; Tanabe, M.; Saito, H.; Ban, Y.; Nawata, H.; Yanase, T. Subtle 17α-hydroxylase/17,20-lyase deficiency with homozygous Y201N mutation in an infertile woman. J. Clin. Endocrinol. Metab. 2005, 90, 2508–2511. [Google Scholar] [CrossRef] [Green Version]
- Yanase, T.; Kagimoto, M.; Suzuki, S.; Hashiba, K.; Simpson, E.R.; Waterman, M.R. Deletion of a phenylalanine in the N-terminal region of human cytochrome P-450(17 alpha) results in partial combined 17 alpha-hydroxylase/17,20-lyase deficiency. J. Biol. Chem. 1989, 264, 18076–18082. [Google Scholar] [CrossRef]
- Yao, F.; Huang, S.; Kang, X.; Zhang, W.; Wang, P.; Tian, Q. CYP17A1 mutations identified in 17 Chinese patients with 17α-hydroxylase/17,20-lyase deficiency. Gynecol. Endocrinol. 2013, 29, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.D.; Hill, M.; Urbanowicz, E.; Rok-Bujko, P.; Bieńkowski, P.; Namysłowska, I.; Mierzejewski, P. Marked elevation of adrenal steroids, especially androgens, in saliva of prepubertal autistic children. Eur. Child Adolesc. Psychiatry 2013, 23, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Baz, F.; Hamza, R.T.; Ayad, M.S.; Mahmoud, N.H. Hyperandrogenemia in male autistic children and adolescents: Relation to disease severity. Int. J. Adolesc. Med. Health 2014, 26, 79–84. [Google Scholar] [CrossRef]
- Katagiri, M.; Kagawa, N. The regulation of steroidogenesis by 17 alpha-hydroxylase/17,20-lyase (P450c17). Folia Pharmacol. Jpn. 1998, 112, 43–50. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Hyun, D.H.; Lee, G.H. Cytochrome b5 reductase, a plasma membrane redox enzyme, protects neuronal cells against metabolic and oxidative stress through maintaining redox state and bioenergetics. Age 2015, 37, 122. [Google Scholar] [CrossRef] [Green Version]
- Popper, K.R. Logik der Forschung; Mohr Siebeck: Tübingen, Germany, 1969. [Google Scholar]
- Xu, S.; Hu, S.; Yu, X.; Zhang, M.; Yang, Y. 17α-hydroxylase/17,20-lyase deficiency in congenital adrenal hyperplasia: A case report. Mol. Med. Rep. 2016, 15, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, C.H.L. Profiling steroid hormones and urinary steroids. J. Chromatogr. 1986, 379, 91–156. [Google Scholar] [CrossRef]
- Shackleton, C.H.L. Role of a Disordered Steroid Metabolome in the Elucidation of Sterol and Steroid Biosynthesis. Lipids 2012, 47, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vogt, B.; Dick, B.; N’Gankam, V.; Frey, F.J.; Frey, B.M. Reduced 11B-hydroxysteroid dehydrogenase activity in patients with the nephrotic syndrome. J. Clin. Endocrinol. Metab. 1999, 84, 811–814. [Google Scholar]
- Jarque, C.M.; Bera, A.K. Efficent tests for normality, homoscedasticity and serial independence of regression residuals. Econ. Lett. 1980, 6, 255–259. [Google Scholar] [CrossRef]
- Gevi, F.; Zolla, L.; Gabriele, S.; Persico, A.M. Urinary metabolomics of young Italian autistic children supports abnormal tryptophan and purine metabolism. Mol. Autism. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shifren, J.L.; Mesiano, S.; Taylor, R.N.; Ferrara, N.; Jaffe, R.B. Corticotropin regulates vascular endothelial growth factor expression in human fetal adrenal cortical cells. J. Clin. Endocrinol. Metab. 1998, 83, 1342–1347. [Google Scholar] [CrossRef]
- Ishimoto, H.; Ginzinger, D.G.; Jaffe, R.B. Adrenocorticotropin preferentially up-regulates angiopoietin 2 in the human fetal adrenal gland: Implications for coordinated adrenal organ growth and angiogenesis. J. Clin. Endocrinol. Metab. 2006, 91, 1909–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, A.; Hahn, D.; Kempná, P.; Hofer, G.; Nuoffer, J.M.; Mullis, P.E.; Flück, C.E. Metformin inhibits human androgen production by regulating steroidogenic enzymes HSD3B2 and CYP17A1 and complex I activity of the respiratory chain. Endocrinology 2012, 153, 4354–4366. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, L. Hypothalamic-pituitary-adrenocortical axis: Neuropsychiatric aspects. Compr. Physiol. 2014, 4, 715–738. [Google Scholar]
- Bitsika, V.; Sharpley, C.F.; Sweeney, J.A.; McFarlane, J.R. HPA and SAM axis responses as correlates of self- vs parental ratings of anxiety in boys with an Autistic Disorder. Physiol. Behav. 2014, 127, 1–7. [Google Scholar] [CrossRef]
- Markou, A.; Sertedaki, A.; Kaltsas, G.; Androulakis, I.; Marakaki, C.; Pappa, T.; Gouli, A.; Panastasiou, L.; Fountoulakis, S.; Zacharoulis, A.; et al. Piaditis, Stress-induced aldosterone hyper-secretion in a substantial subset of patients with essential hypertension. J. Clin. Endocrinol. Metab. 2015, 100, 2857–2864. [Google Scholar] [CrossRef] [Green Version]
- Cheong, E.J.Y.; Nair, P.; Neo, R.W.Y.; Tu, H.T.; Lin, F.; Chiong, E.; Esuvaranathan, K.; Fan, H.; Szmulewitz, R.Z.; Peer, C.J.; et al. Slow-, Tight-Binding Inhibition of CYP17A1 by Abiraterone Redefines Its Kinetic Selectivity and Dosing Regimen. J. Pharmacol. Exp. Ther. 2020, 374, 438–451. [Google Scholar] [CrossRef]
- Wróbel, T.M.; Rogova, O.; Sharma, K.; Rojas Velazquez, M.N.; Pandey, A.V.; Jørgensen, F.S.; Arendrup, F.S.; Andersen, K.L.; Björkling, F. Synthesis and Structure-Activity Relationships of Novel Non-Steroidal CYP17A1 Inhibitors as Potential Prostate Cancer Agents. Biomolecules 2022, 12, 165. [Google Scholar] [CrossRef]
- Auchus, R.J. Steroid 17-hydroxylase and 17,20-lyase deficiencies, genetic and pharmacologic. J. Steroid Biochem. Mol. Biol. 2016, 165, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boris, M.; Kaiser, C.C.; Goldblatt, A.; Elice, M.W.; Edelson, S.M.; Adams, J.B.; Feinstein, D.L. Effect of pioglitazone treatment on behavioral symptoms in autistic children. J. Neuroinflamm. 2007, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, B.A.; Kurz, J.; Senn, W.; Escher, G.; Mohaupt, M.G. Stress-induced alterations of social behavior are reversible by antagonism of steroid hormones in C57/BL6 mice. Naunyn. Schmiedebergs Arch. Pharmacol. 2021, 394, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Wink, L.K.; Adams, R.; Pedapati, E.V.; Dominick, K.C.; Fox, E.; Buck, C.; Erickson, C.A. Brief report: Metformin for antipsychotic_induced weight gain in youth with autism spectrum disorder. J. Autism Dev. Disord. 2017, 47, 2290–2294. [Google Scholar] [CrossRef]
- DeVore, N.M.; Scott, E.E. Structures of cytochrome P450 17A1 with prostate cancer drugs abiraterone and TOK-001. Nature 2012, 482, 116–119. [Google Scholar] [CrossRef] [Green Version]
- Viollet, B.; Guigas, B.; Garcia, N.S.; Leclerc, J.; Foretz, M.; Andreelli, F. Cellular and Molecular Mechanisms of Metformin: An Overview. Clin. Sci. 2012, 122, 253–270. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.; Gu, F.; Essa, M.M.; Wegiel, J.; Kaur, K.; Brown, W.T.; Chauhan, V. Brain region-specific deficit in mitochondrial electron transport chain complexes in children with autism. J. Neurochem. 2011, 117, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, D.A.; Frye, R.E. Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism. Front. Physiol. 2014, 5, 150. [Google Scholar] [CrossRef] [Green Version]
- Tang, G.; Gutierrez Rios, P.; Kuo, S.-H.; Akman, H.O.; Rosoklija, G.; Tanji, K.; Dwork, A.; Schon, E.A.; DiMauro, S.; Goldman, J.; et al. Mitochondrial abnormalities in temporal lobe of autistic brain. Neurobiol. Dis. 2013, 54, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Rose, S.; Melnyk, S.; Pavliv, O.; Bai, S.; Nick, T.G.; Frye, R.E.; James, S.J. Evidence of oxidative damage and inflammation associated with low glutathione redox status in the autism brain. Transl. Psychiatry 2012, 2, e134. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.; Chauhan, V.; Chauhan, A. Impaired synthesis and antioxidant defense of glutathione in the cerebellum of autistic subjects: Alterations in the activities and protein expression of glutathione-related enzymes. Free Radic. Biol. Med. 2013, 65, 488–496. [Google Scholar] [CrossRef]
- Hartwig, J.; Loebel, M.; Steiner, S.; Bauer, S.; Karadeniz, Z.; Roeger, C.; Skurk, C.; Scheibenbogen, C.; Sotzny, F. Metformin Attenuates ROS via FOXO3 Activation in Immune Cells. Front. Immunol. 2021, 12, 581799. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Han, B.; Fan, M.; Wang, N.; Wang, H.; Zhu, H.; Cheng, T.; Zhao, S.; Song, H.; Qiao, J. Oxidative stress increases the 17,20-lyase-catalyzing activity of adrenal P450c17 through p38α in the development of hyperandrogenism. Mol. Cell. Endocrinol. 2019, 484, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Yorbik, O.; Sayal, A.; Akay, C.; Akbiyik, D.I.; Sohmen, T. Investigation of antioxidant enzymes in children with autistic disorder. Prostaglandins Leukot. Essent. Fat. Acids 2002, 67, 341–343. [Google Scholar] [CrossRef] [PubMed]
- James, S.J.; Cutler, P.; Melnyk, S.; Jernigan, S.; Janak, L.; Gaylor, D.W.; Neubrander, J.A. Metabolic biomarkers of increased oxidative stress and impaired methylation capacity in children with autism. Am. J. Clin. Nutr. 2004, 80, 1611–1617. [Google Scholar] [PubMed] [Green Version]
- James, S.J.; Melnyk, S.; Fuchs, G.; Reid, T.; Jernigan, S.; Pavliv, O.; Hubanks, A.; Gaylor, D.W. Efficacy of methylcobalamin and folinic acid treatment on glutathione redox status in children with autism. Am. J. Clin. Nutr. 2008, 89, 425–430. [Google Scholar] [CrossRef]
- James, S.J.; Melnyk, S.; Jernigan, S.; Cleves, M.A.; Halsted, C.H.; Wong, D.H.; Cutler, P.; Bock, K.; Boris, M.; Bradstreet, J.J.; et al. Metabolic endophenotype and related genotypes are associated with oxidative stress in children with autism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Ming, X.; Brimacombe, M.; Chaaban, J.; Zimmerman-Bier, B.; Wagner, G.C. Autism spectrum disorders: Concurrent clinical disorders. J. Child Neurol. 2007, 23, 6–13. [Google Scholar] [CrossRef]
- Chauhan, A.; Chauhan, V. Oxidative stress in autism. Pathophysiology 2006, 13, 171–181. [Google Scholar] [CrossRef]
- Yao, Y.; Walsh, W.J.; McGinnis, W.R.; Praticò, D. Altered vascular phenotype in autism: Correlation with oxidative stress. Arch. Neurol. 2006, 63, 1161–1164. [Google Scholar] [CrossRef] [Green Version]
- Al-Gadani, Y.; El-Ansary, A.; Attas, O.; Al-Ayadhi, L. Metabolic biomarkers related to oxidative stress and antioxidant status in Saudi autistic children. Clin. Biochem. 2009, 42, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, S.; Fuchs, G.J.; Schulz, E.; Lopez, M.; Kahler, S.G.; Fussell, J.J.; Bellando, J.; Pavliv, O.; Rose, S.; Seidel, L.; et al. Metabolic imbalance associated with methylation dysregulation and oxidative damage in children with autism. J. Autism Dev. Disord. 2011, 42, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boris, M.; Goldblatt, A.; Galanko, J.; James, S.J. Association of MTHFR gene variants with autism. J. Am. Phys. Surg. 2004, 9, 106–108. Available online: http://www.jpands.org/vol9no4/boris.pdf (accessed on 15 March 2022).
- Bowers, K.; Li, Q.; Bressler, J.; Avramopoulos, D.; Newschaffer, C.; Fallin, M.D. Glutathione pathway gene variation and risk of autism spectrum disorders. J. Neurodev. Disord. 2011, 3, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Frustaci, A.; Neri, M.; Cesario, A.; Adams, J.B.; Domenici, E.; Bernardina, B.D.; Bonassi, S. Oxidative stress-related biomarkers in autism: Systematic review and meta-analyses. Free Radic. Biol. Med. 2012, 52, 2128–2141. [Google Scholar] [CrossRef]
- Goin-Kochel, R.P.; Porter, A.E.; Peters, S.U.; Shinawi, M.; Sahoo, T.; Beaudet, A.L. The MTHFR 677C–>T polymorphism and behaviors in children with autism: Exploratory genotype-phenotype correlations. Autism Res. 2009, 2, 98–108. [Google Scholar] [CrossRef]
- Guo, T.; Chen, H.; Liu, B.; Ji, W.; Yang, C. Methylenetetrahydrofolate reductase polymorphisms C677T and risk of autism in the Chinese Han population. Genet. Test. Mol. Biomark. 2012, 16, 968–973. [Google Scholar] [CrossRef]
- James, S.J.; Rose, S.; Melnyk, S.; Jernigan, S.; Blossom, S.; Pavliv, O.; Gaylor, D.W. Cellular and mitochondrial glutathione redox imbalance in lymphoblastoid cells derived from children with autism. FASEB J. 2009, 23, 2374–2383. [Google Scholar] [CrossRef]
- Adams, J.B.; Baral, M.; Geis, E.; Mitchell, J.; Ingram, J.; Hensley, A.; Zappia, I.; Newmark, S.; Gehn, E.; Rubin, R.A.; et al. The severity of autism is associated with toxic metal body burden and red blood cell glutathione levels. J. Toxicol. 2009, 2009, 532640. [Google Scholar] [CrossRef] [Green Version]
- Ghezzo, A.; Visconti, P.; Abruzzo, P.M.; Bolotta, A.; Ferreri, C.; Gobbi, G.; Malisardi, G.; Manfredini, S.; Marini, M.; Nanetti, L.; et al. Oxidative stress and erythrocyte membrane alterations in children with autism: Correlation with clinical features. PLoS ONE 2013, 8, e66418. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.A.; Siedlak, S.L.; Lu, L.; Fu, X.; Wang, Z.; McGinnis, W.R.; Fakhoury, E.; Castellani, R.J.; Hazen, S.L.; Walsh, W.J.; et al. The autistic phenotype exhibits a remarkably localized modification of brain protein by products of free radical-induced lipid oxidation. Am. J. Biochem. Biotechnol. 2008, 4, 61–72. [Google Scholar] [CrossRef] [Green Version]
- López-Hurtado, E.; Prieto, J.J. A microscopic study of language-related cortex in autism. Am. J. Biochem. Biotechnol. 2008, 4, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Sajdel-Sul, E.M.; Windom, H.; McGinnis, W.; Lipinski, B.; Audhya, T. Oxidative stress in autism: Elevated cerebellar 3-nitrotyrosine levels. Am. J. Biochem. Biotechnol. 2008, 4, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Sajdel-Sulkowska, E.M.; Xu, M.; Koibuchi, N. Increase in cerebellar neurotrophin-3 and oxidative stress markers in autism. Cerebellum 2009, 8, 366–372. [Google Scholar] [CrossRef]
- Sajdel-Sulkowska, E.M.; Xu, M.; McGinnis, W.; Koibuchi, N. Brain region-specific changes in oxidative stress and neurotrophin levels in autism spectrum disorders (ASD). Cerebellum 2010, 10, 43–48. [Google Scholar] [CrossRef]
- Palmieri, L.; Papaleo, V.; Porcelli, V.; Scarcia, P.; Gaita, L.; Sacco, R.; Hager, J.; Rousseau, F.; Curatolo, P.; Manzi, B.; et al. Altered calcium homeostasis in autism-spectrum disorders: Evidence from biochemical and genetic studies of the mitochondrial aspartate/glutamate carrier AGC1. Mol. Psychiatry 2008, 15, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Galano, A.; Tan, D.-X.; Reiter, R.J. Melatonin: A Versatile Protector against Oxidative DNA Damage. Molecules 2018, 23, 530. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Urinary Steroid Hormone Metabolites | Boys | Girls | ||||
|---|---|---|---|---|---|---|
| [μg/mmol creatinine] | Autistic | p< | Control | Autistic | p< | Control |
| Androsterone | 88.2 ± 10.9 | <0.01 | 49.5 ± 7.1 | 80.4 ± 21.9 | 0.06 | 52.9 ± 11.6 |
| Etiocholanolone | 57.7 ± 8.0 | <0.01 | 28.8 ± 4.1 | 65.0 ± 17.1 | 0.07 | 35.8 ± 10.1 |
| DHEA | 18.8 ± 8.8 | 0.11 | 4.6 ± 1.0 | 14.5 ± 7.9 | 0.18 | 4.7 ± 2.5 |
| Androstendione | 5.0 ± 1.2 | 0.08 | 2.7 ± 0.4 | 6.7 ± 2.4 | 0.06 | 2.4 ± 0.6 |
| Testosterone | 2.8 ± 0.5 | <0.01 | 1.1 ± 0.2 | 0.8 ± 0.1 | 0.01 | 0.6 ± 0.1 |
| TH-11β-DH-Corticosterone | 14.5 ± 1.3 | <0.01 | 10.9 ± 1.0 | 10.7 ± 1.6 | 0.88 | 9.5 ± 1.2 |
| TH-Corticosterone | 14.6 ± 1.1 | <0.01 | 10.5 ± 0.8 | 9.7 ± 1.1 | 0.93 | 8.8 ± 1.5 |
| 5α-TH-Corticosterone | 43.7 ± 7.8 | <0.01 | 26.6 ± 2.6 | 18.9 ± 2.6 | 0.63 | 21.3 ± 4.5 |
| Cortisol | 11.5 ± 1.6 | <0.01 | 7.2 ± 0.7 | 6.3 ± 0.7 | 0.04 | 4.5 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasser, B.A.; Kurz, J.; Dick, B.; Mohaupt, M.G. How Is CYP17A1 Activity Altered in Autism? A Pilot Study to Identify Potential Pharmacological Targets. Life 2022, 12, 867. https://doi.org/10.3390/life12060867
Gasser BA, Kurz J, Dick B, Mohaupt MG. How Is CYP17A1 Activity Altered in Autism? A Pilot Study to Identify Potential Pharmacological Targets. Life. 2022; 12(6):867. https://doi.org/10.3390/life12060867
Chicago/Turabian StyleGasser, Benedikt Andreas, Johann Kurz, Bernhard Dick, and Markus Georg Mohaupt. 2022. "How Is CYP17A1 Activity Altered in Autism? A Pilot Study to Identify Potential Pharmacological Targets" Life 12, no. 6: 867. https://doi.org/10.3390/life12060867
APA StyleGasser, B. A., Kurz, J., Dick, B., & Mohaupt, M. G. (2022). How Is CYP17A1 Activity Altered in Autism? A Pilot Study to Identify Potential Pharmacological Targets. Life, 12(6), 867. https://doi.org/10.3390/life12060867