
Data Mining Identifies CCN2 and THBS1 as Biomarker Candidates for Cardiac Hypertrophy

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
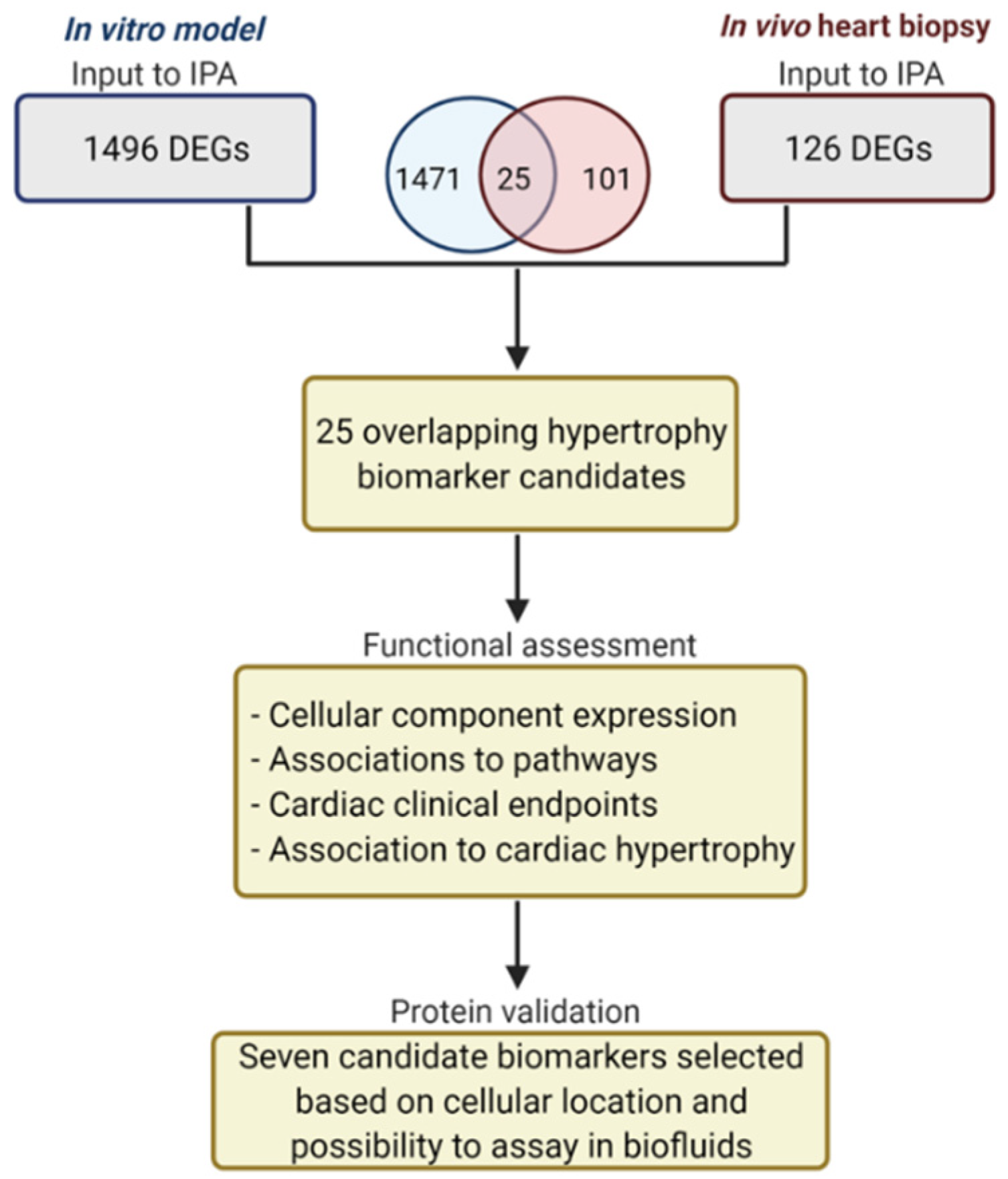
2.1. Transcriptomics Datasets from In Vitro and In Vivo Experiments
2.2. Differentially Expressed Genes in Biopsies from Cardiac Hypertrophic Biopsies
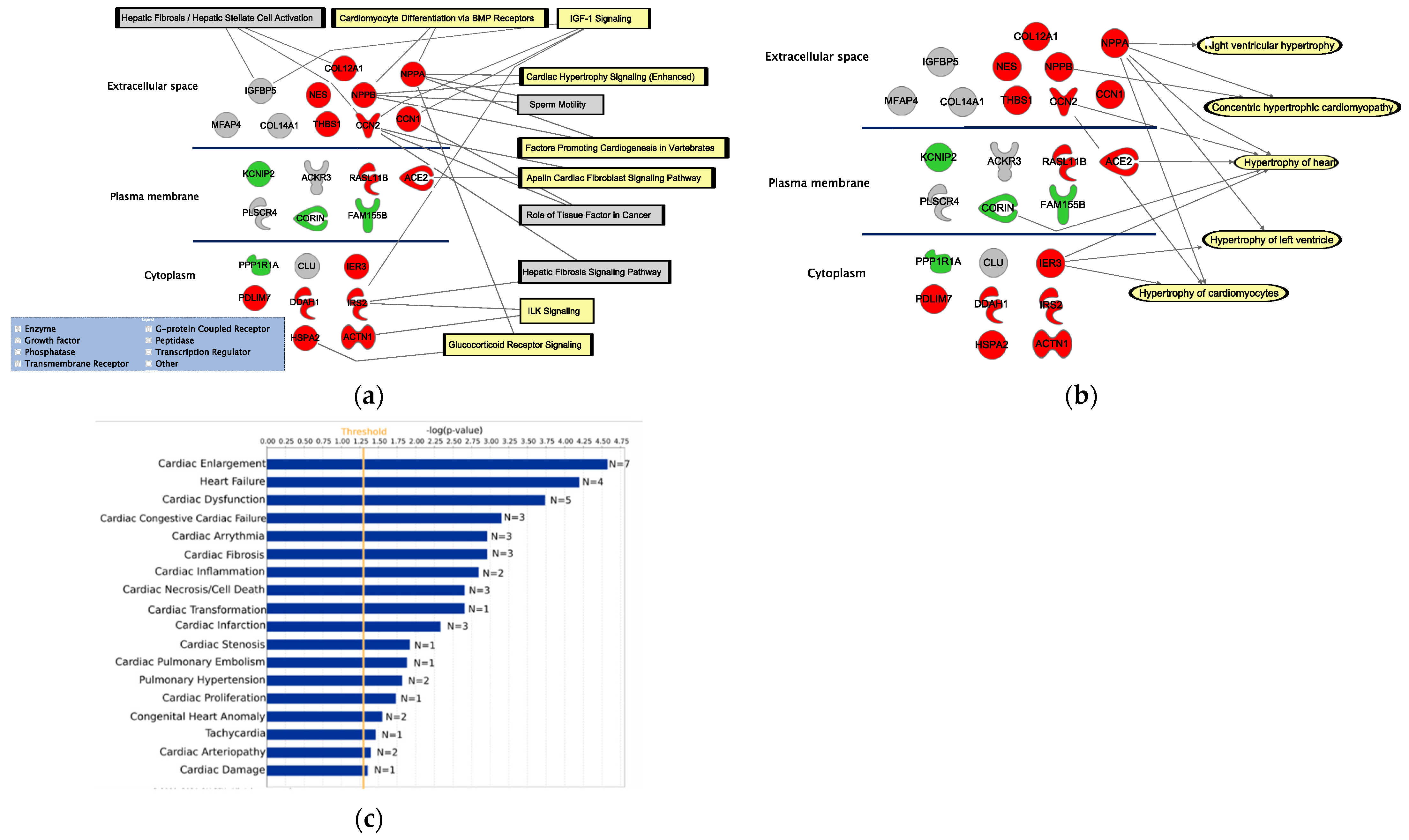
2.3. Functional Annotation Analysis of Differentially Expressed Genes Using IPA
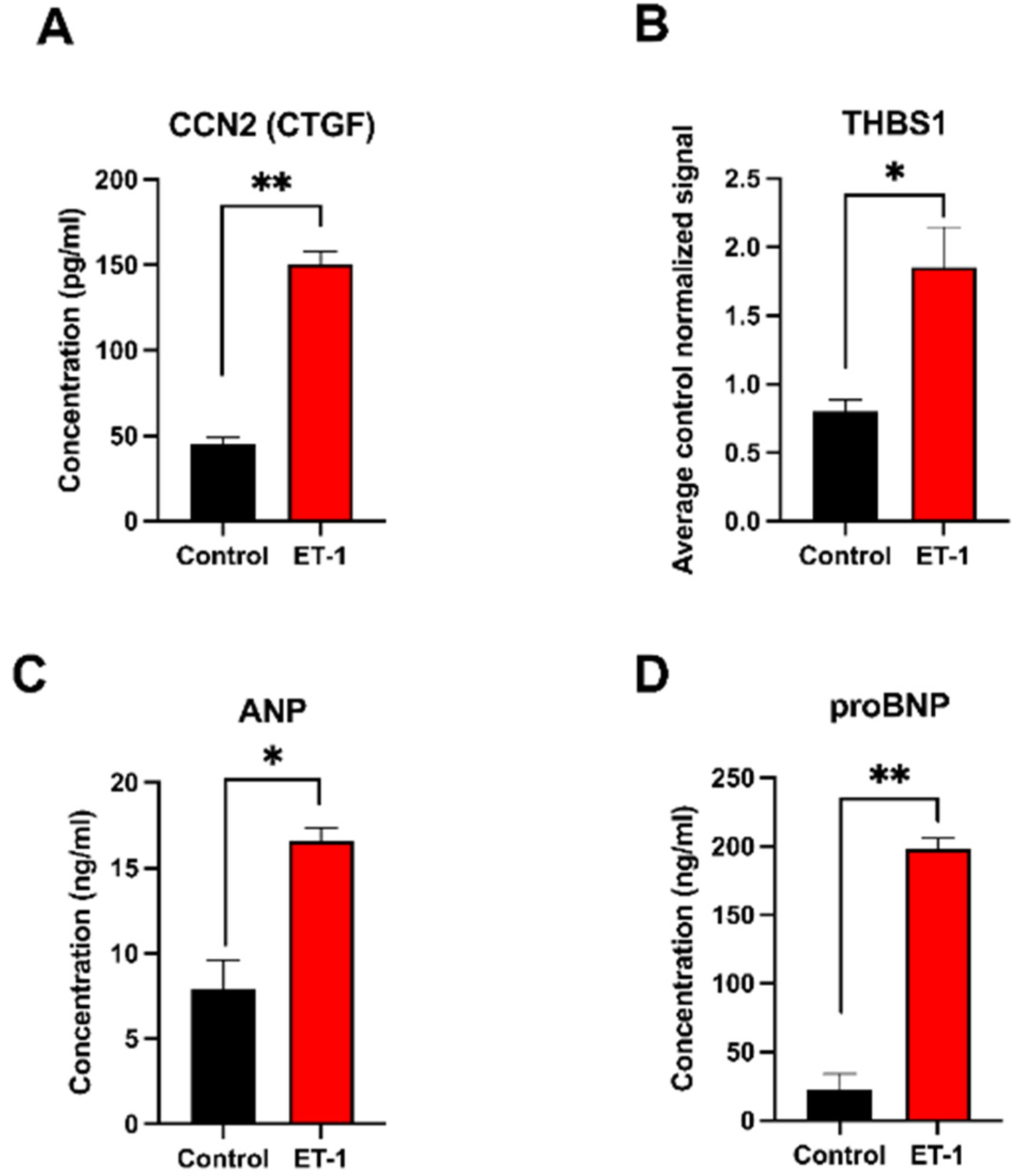
2.4. Protein Validation of Selected Cardiac Hypertrophy Candidates
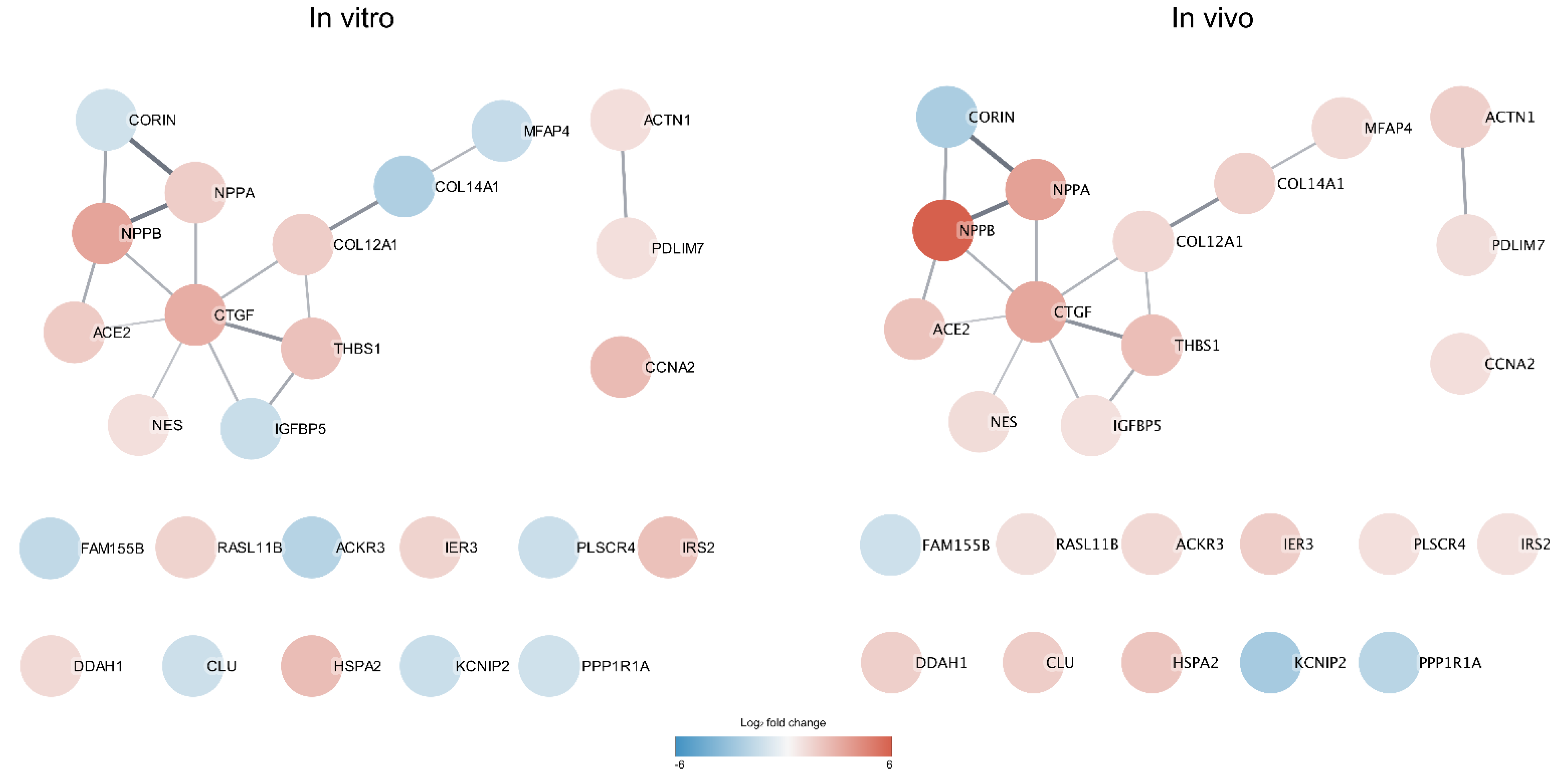
2.5. Protein-Network Analysis
2.6. Gene Ontology Enrichment Analysis
3. Results
3.1. Transcriptomic Analysis of In Vitro and In Vivo Data
3.2. Functional Assessment of the 25 DEGs
3.3. Protein Validation of the Selected DEGs
3.4. Protein Interaction Network Analysis
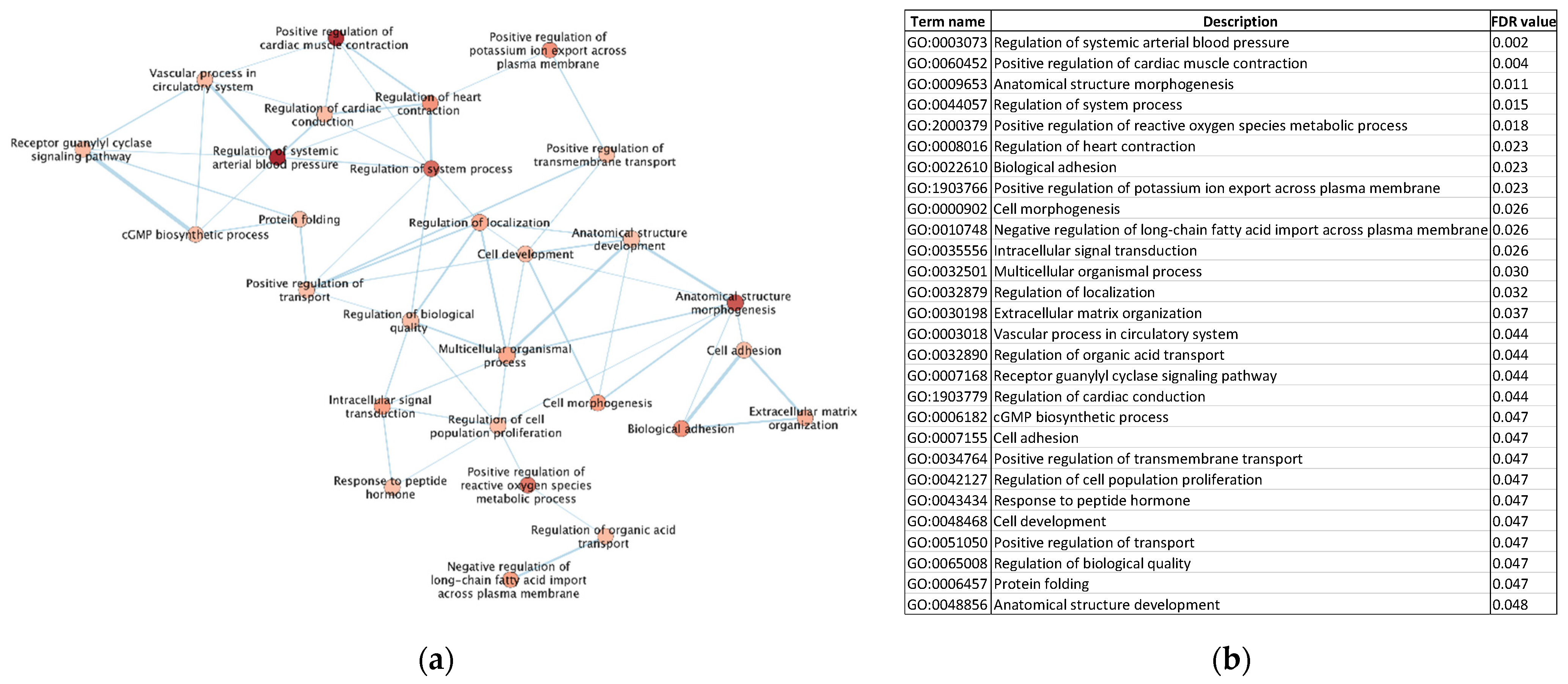
3.5. Gene Ontology (GO) Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | 8 h | 24 h | 48 h | 72 h | 96 h |
|---|---|---|---|---|---|
| ACE2 | 1.831 | 1.358 | 1.432 | ||
| ACKR3 | −1.116 | −1.738 | −1.119 | ||
| ACTN1 | 1.066 | ||||
| CCN1 | 2.431 | 1.053 | |||
| CCN2 | 2.999 | 1.568 | |||
| CLU | −1.066 | −1.125 | |||
| COL12A1 | 1.335 | 1.745 | 1.299 | 1.255 | 1.259 |
| COL14A1 | −1.938 | −1.661 | −1.557 | ||
| CORIN | −1.033 | ||||
| DDAH1 | 1.283 | ||||
| FAM155B | −1.419 | ||||
| HSPA2 | 1.997 | 2.369 | 1.101 | ||
| IER3 | 1.517 | 1.182 | |||
| IGFBP5 | −1.235 | ||||
| IRS2 | 2.218 | ||||
| KCNIP2 | −1.186 | ||||
| MFAP4 | −1.098 | −1.228 | −1.305 | −1.166 | |
| NES | 1.053 | ||||
| NPPA | 1.048 | 1.754 | 1.517 | 1.025 | 1.208 |
| NPPB | 3.334 | 2.903 | 1.082 | ||
| PDLIM7 | 1.031 | ||||
| PLSCR4 | −1.162 | ||||
| PPP1R1A | −1.039 | ||||
| RASL11B | 1.524 | 1.174 | |||
| THBS1 | 2.204 | 1.118 |
Appendix B
| Gene Symbol | Normal EF | Low EF |
|---|---|---|
| ACE2 | 2.145 | |
| ACKR3 | 1.407 | 1.203 |
| ACTN1 | 1.662 | |
| CCN1 | 1.09 | |
| CCN2 | 3.25 | 3.225 |
| CLU | 1.74 | |
| COL12A1 | 1.36 | |
| COL14A1 | 1.86 | 1.381 |
| CORIN | −2.035 | |
| DDAH1 | 1.511 | 1.843 |
| FAM155B | −1.073 | |
| HSPA2 | 1.754 | 2.355 |
| IER3 | 1.747 | |
| IGFBP5 | 1.002 | |
| IRS2 | 1.001 | |
| KCNIP2 | −2.177 | |
| MFAP4 | 1.284 | |
| NES | 1.173 | |
| NPPA | 3.459 | |
| NPPB | 5.987 | |
| PDLIM7 | 1.108 | |
| PLSCR4 | 1.059 | |
| PPP1R1A | −1.637 | −1.659 |
| RASL11B | 1.094 | |
| THBS1 | 2.329 |
References
- Frey, N.; Katus, H.A.; Olson, E.N.; Hill, J.A. Hypertrophy of the heart: A new therapeutic target? Circulation 2004, 109, 1580–1589. [Google Scholar] [CrossRef] [Green Version]
- Pluim, B.M.; Zwinderman, A.H.; van der Laarse, A.; van der Wall, E.E. The athlete's heart. A meta-analysis of cardiac structure and function. Circulation 2000, 101, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Eghbali, M.; Deva, R.; Alioua, A.; Minosyan, T.Y.; Ruan, H.; Wang, Y.; Toro, L.; Stefani, E. Molecular and functional signature of heart hypertrophy during pregnancy. Circ. Res. 2005, 96, 1208–1216. [Google Scholar] [CrossRef] [Green Version]
- Janz, K.F.; Dawson, J.D.; Mahoney, L.T. Predicting heart growth during puberty: The Muscatine Study. Pediatrics 2000, 105, E63. [Google Scholar] [CrossRef] [Green Version]
- Badiani, S.; van Zalen, J.; Treibel, T.A.; Bhattacharyya, S.; Moon, J.C.; Lloyd, G. Aortic Stenosis, a Left Ventricular Disease: Insights from Advanced Imaging. Curr. Cardiol. Rep. 2016, 18, 80. [Google Scholar] [CrossRef] [Green Version]
- Marian, A.J. Genetic determinants of cardiac hypertrophy. Curr. Opin. Cardiol. 2008, 23, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Frey, N.; Olson, E.N. Cardiac hypertrophy: The good, the bad, and the ugly. Annu. Rev. Physiol. 2003, 65, 45–79. [Google Scholar] [CrossRef]
- Almeida, P.; Azevedo, A.; Rodrigues, R.; Dias, P.; Frioes, F.; Vazquez, B.; Abreu-Lima, C.; Bettencourt, P.; Barros, H. B-type natriuretic peptide and left ventricular hypertrophy in hypertensive patients. Rev. Port. Cardiol. 2003, 22, 327–336. [Google Scholar]
- Kolwicz, S.C., Jr.; Tian, R. Glucose metabolism and cardiac hypertrophy. Cardiovasc. Res. 2011, 90, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Ulfenborg, B.; Andersson, C.X.; Heydarkhan-Hagvall, S.; Jeppsson, A.; Sartipy, P.; Synnergren, J. Cardiac hypertrophy in a dish: A human stem cell based model. Biol. Open 2020, 9, bio052381. [Google Scholar] [CrossRef]
- Carlson, C.; Koonce, C.; Aoyama, N.; Einhorn, S.; Fiene, S.; Thompson, A.; Swanson, B.; Anson, B.; Kattman, S. Phenotypic screening with human iPS cell-derived cardiomyocytes: HTS-compatible assays for interrogating cardiac hypertrophy. J. Biomol. Screen 2013, 18, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, P.; Turner, A.; Matter, A.; Kattman, S.J.; Stoddard, A.; Lorier, R.; Swanson, B.J.; Arnett, D.K.; Broeckel, U. RNA expression profiling of human iPSC-derived cardiomyocytes in a cardiac hypertrophy model. PLoS ONE 2014, 9, e108051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaBarge, W.; Mattappally, S.; Kannappan, R.; Fast, V.G.; Pretorius, D.; Berry, J.L.; Zhang, J. Maturation of three-dimensional, hiPSC-derived cardiomyocyte spheroids utilizing cyclic, uniaxial stretch and electrical stimulation. PLoS ONE 2019, 14, e0219442. [Google Scholar] [CrossRef]
- Johansson, M.; Ulfenborg, B.; Andersson, C.X.; Heydarkhan-Hagvall, S.; Jeppsson, A.; Sartipy, P.; Synnergren, J. Multi-Omics Characterization of a Human Stem Cell-Based Model of Cardiac Hypertrophy. Life 2022, 12, 293. [Google Scholar] [CrossRef]
- Bupha-Intr, T.; Haizlip, K.M.; Janssen, P.M. Role of endothelin in the induction of cardiac hypertrophy in vitro. PLoS ONE 2012, 7, e43179. [Google Scholar] [CrossRef] [Green Version]
- Jen, H.L.; Yin, W.H.; Chen, J.W.; Lin, S.J. Endothelin-1-Induced Cell Hypertrophy in Cardiomyocytes is Improved by Fenofibrate: Possible Roles of Adiponectin. J. Atheroscler. Thromb. 2017, 24, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Li, Q.; May, H.I.; Li, C.; Zhang, G.; Sharma, G.; Sherry, A.D.; Malloy, C.R.; Khemtong, C.; Zhang, Y.; et al. Lactate Dehydrogenase A Governs Cardiac Hypertrophic Growth in Response to Hemodynamic Stress. Cell Rep. 2020, 32, 108087. [Google Scholar] [CrossRef]
- Ulfenborg, B. Vertical and horizontal integration of multi-omics data with miodin. BMC Bioinform. 2019, 20, 649. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, F.; Miyagawa, S.; Yasuda, S.; Miura, T.; Kuroda, T.; Itoh, M.; Kawaji, H.; Ito, E.; Yoshida, S.; Saito, A.; et al. CXCL4/PF4 is a predictive biomarker of cardiac differentiation potential of human induced pluripotent stem cells. Sci. Rep. 2019, 9, 4638. [Google Scholar] [CrossRef]
- Bharathan, S.P.; Manian, K.V.; Aalam, S.M.; Palani, D.; Deshpande, P.A.; Pratheesh, M.D.; Srivastava, A.; Velayudhan, S.R. Systematic evaluation of markers used for the identification of human induced pluripotent stem cells. Biol. Open 2017, 6, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.S.; Øie, E.; Vinge, L.E.; Yndestad, A.; Øystein Andersen, G.; Andersson, Y.; Attramadal, T.; Attramadal, H. Connective tissue growth factor--a novel mediator of angiotensin II-stimulated cardiac fibroblast activation in heart failure in rats. J. Mol. Cell. Cardiol. 2004, 36, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Lam, A.; Abraham, J.A.; Schreiner, G.F.; Joly, A.H. CTGF expression is induced by TGF- beta in cardiac fibroblasts and cardiac myocytes: A potential role in heart fibrosis. J. Mol. Cell. Cardiol. 2000, 32, 1805–1819. [Google Scholar] [CrossRef]
- Way, K.J.; Isshiki, K.; Suzuma, K.; Yokota, T.; Zvagelsky, D.; Schoen, F.J.; Sandusky, G.E.; Pechous, P.A.; Vlahos, C.J.; Wakasaki, H.; et al. Expression of connective tissue growth factor is increased in injured myocardium associated with protein kinase C beta2 activation and diabetes. Diabetes 2002, 51, 2709–2718. [Google Scholar] [CrossRef] [Green Version]
- Hayata, N.; Fujio, Y.; Yamamoto, Y.; Iwakura, T.; Obana, M.; Takai, M.; Mohri, T.; Nonen, S.; Maeda, M.; Azuma, J. Connective tissue growth factor induces cardiac hypertrophy through Akt signaling. Biochem. Biophys. Res. Commun. 2008, 370, 274–278. [Google Scholar] [CrossRef]
- Koitabashi, N.; Arai, M.; Niwano, K.; Watanabe, A.; Endoh, M.; Suguta, M.; Yokoyama, T.; Tada, H.; Toyama, T.; Adachi, H.; et al. Plasma connective tissue growth factor is a novel potential biomarker of cardiac dysfunction in patients with chronic heart failure. Eur. J. Heart Fail. 2008, 10, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Koitabashi, N.; Arai, M.; Kogure, S.; Niwano, K.; Watanabe, A.; Aoki, Y.; Maeno, T.; Nishida, T.; Kubota, S.; Takigawa, M.; et al. Increased Connective Tissue Growth Factor Relative to Brain Natriuretic Peptide as a Determinant of Myocardial Fibrosis. Hypertension 2007, 49, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Gerritsen, K.G.; Falke, L.L.; van Vuuren, S.H.; Leeuwis, J.W.; Broekhuizen, R.; Nguyen, T.Q.; de Borst, G.J.; Nathoe, H.M.; Verhaar, M.C.; Kok, R.J.; et al. Plasma CTGF is independently related to an increased risk of cardiovascular events and mortality in patients with atherosclerotic disease: The SMART study. Growth Factors 2016, 34, 149–158. [Google Scholar] [CrossRef]
- Patel, V.; Syrris, P.; Coats, C.; Lucena, J.; Lara-Pezzi, E.; Garcia-Pavia, P.; Elliott, P.M. Genetic regulation of myocardial fibrosis in hypertrophic cardiomyopathy. Eur. Heart J. 2021, 42, ehab724.1778. [Google Scholar] [CrossRef]
- Staniszewska, I.; Zaveri, S.; Del Valle, L.; Oliva, I.; Rothman, V.L.; Croul, S.E.; Roberts, D.D.; Mosher, D.F.; Tuszynski, G.P.; Marcinkiewicz, C. Interaction of alpha9beta1 integrin with thrombospondin-1 promotes angiogenesis. Circ. Res. 2007, 100, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Dawson, D.W.; Pearce, S.F.; Zhong, R.; Silverstein, R.L.; Frazier, W.A.; Bouck, N.P. CD36 mediates the In vitro inhibitory effects of thrombospondin-1 on endothelial cells. J. Cell Biol. 1997, 138, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Martin-Manso, G.; Pendrak, M.L.; Garfield, S.H.; Isenberg, J.S.; Roberts, D.D. Thrombospondin-1 inhibits VEGF receptor-2 signaling by disrupting its association with CD47. J. Biol. Chem. 2010, 285, 38923–38932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, H.R.; Campbell, G.E.; Aytoda, P.; Mathiesen, A.H.; Duffy, D.M. Thrombospondin 1 (THBS1) Promotes Follicular Angiogenesis, Luteinization, and Ovulation in Primates. Front. Endocrinol. 2019, 10, 727. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Guo, B.; Lin, F.; Hui, Q.; Tao, K. Effect of THBS1 on the Biological Function of Hypertrophic Scar Fibroblasts. Biomed Res. Int. 2020, 2020, 8605407. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, D.; Schips, T.G.; Vo, A.; Grimes, K.M.; Baldwin, T.A.; Brody, M.J.; Accornero, F.; Sargent, M.A.; Molkentin, J.D. Thbs1 induces lethal cardiac atrophy through PERK-ATF4 regulated autophagy. Nat. Commun. 2021, 12, 3928. [Google Scholar] [CrossRef]
- Schellings, M.W.; van Almen, G.C.; Sage, E.H.; Heymans, S. Thrombospondins in the heart: Potential functions in cardiac remodeling. J. Cell Commun. Signal 2009, 3, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G.; Ren, G.; Dewald, O.; Zymek, P.; Haudek, S.; Koerting, A.; Winkelmann, K.; Michael, L.H.; Lawler, J.; Entman, M.L. Critical role of endogenous thrombospondin-1 in preventing expansion of healing myocardial infarcts. Circulation 2005, 111, 2935–2942. [Google Scholar] [CrossRef] [Green Version]
- Schroen, B.; Heymans, S.; Sharma, U.; Blankesteijn, W.M.; Pokharel, S.; Cleutjens, J.P.; Porter, J.G.; Evelo, C.T.; Duisters, R.; van Leeuwen, R.E.; et al. Thrombospondin-2 is essential for myocardial matrix integrity: Increased expression identifies failure-prone cardiac hypertrophy. Circ. Res. 2004, 95, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Tsoutsman, T.; Wang, X.; Garchow, K.; Riser, B.; Twigg, S.; Semsarian, C. CCN2 plays a key role in extracellular matrix gene expression in severe hypertrophic cardiomyopathy and heart failure. J. Mol. Cell. Cardiol. 2013, 62, 164–178. [Google Scholar] [CrossRef]
- Sharifi-Sanjani, M.; Shoushtari, A.H.; Quiroz, M.; Baust, J.; Sestito, S.F.; Mosher, M.; Ross, M.; McTiernan, C.F.; St. Croix, C.M.; Bilonick, R.A.; et al. Cardiac CD47 Drives Left Ventricular Heart Failure Through Ca2+-CaMKII-Regulated Induction of HDAC3. J. Am. Heart Assoc. 2014, 3, e000670. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.N. Biology of natriuretic peptides and their receptors. Peptides 2005, 26, 901–932. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Bigatti, G.; Evangelista, A.; Lanzani, C.; Stanzione, R.; Zagato, L.; Manunta, P.; Marchitti, S.; Venturelli, V.; Bianchi, G.; et al. Association of atrial natriuretic peptide and type a natriuretic peptide receptor gene polymorphisms with left ventricular mass in human essential hypertension. J. Am. Coll. Cardiol. 2006, 48, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, I.; Tokudome, T.; Nakao, K.; Kangawa, K. Natriuretic peptide system: An overview of studies using genetically engineered animal models. FEBS J. 2011, 278, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- von Lueder, T.G.; Sangaralingham, S.J.; Wang, B.H.; Kompa, A.R.; Atar, D.; Burnett, J.C., Jr.; Krum, H. Renin-angiotensin blockade combined with natriuretic peptide system augmentation: Novel therapeutic concepts to combat heart failure. Circ. Heart Fail. 2013, 6, 594–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, K.R.; Knowlton, K.U.; Zhu, H.; Chien, S. Regulation of cardiac gene expression during myocardial growth and hypertrophy: Molecular studies of an adaptive physiologic response. FASEB J. 1991, 5, 3037–3046. [Google Scholar] [CrossRef] [PubMed]
- Cameron, V.A.; Aitken, G.D.; Ellmers, L.J.; Kennedy, M.A.; Espiner, E.A. The sites of gene expression of atrial, brain, and C-type natriuretic peptides in mouse fetal development: Temporal changes in embryos and placenta. Endocrinology 1996, 137, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Houweling, A.C.; Somi, S.; Massink, M.P.; Groenen, M.A.; Moorman, A.F.; Christoffels, V.M. Comparative analysis of the natriuretic peptide precursor gene cluster in vertebrates reveals loss of ANF and retention of CNP-3 in chicken. Dev. Dyn. 2005, 233, 1076–1082. [Google Scholar] [CrossRef]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020, 17, 698–717. [Google Scholar] [CrossRef]
- Sergeeva, I.A.; Hooijkaas, I.B.; Ruijter, J.M.; van der Made, I.; de Groot, N.E.; van de Werken, H.J.; Creemers, E.E.; Christoffels, V.M. Identification of a regulatory domain controlling the Nppa-Nppb gene cluster during heart development and stress. Development 2016, 143, 2135–2146. [Google Scholar] [CrossRef] [Green Version]
- Man, J.C.K.; van Duijvenboden, K.; Krijger, P.H.L.; Hooijkaas, I.B.; van der Made, I.; de Gier-de Vries, C.; Wakker, V.; Creemers, E.E.; de Laat, W.; Boukens, B.J.; et al. Genetic Dissection of a Super Enhancer Controlling the Nppa-Nppb Cluster in the Heart. Circ. Res. 2021, 128, 115–129. [Google Scholar] [CrossRef]





 ), downregulated (
), downregulated (  ), or non-differentially expressed gene (
), or non-differentially expressed gene (  ) at the different time points (8, 24, 48, 72, and 96 h; n = 3 each) in the in vitro samples and in the cardiac samples with a normal EF (n = 3) or low EF (n = 3).
), downregulated ( ), or non-differentially expressed gene ( ) at the different time points (8, 24, 48, 72, and 96 h; n = 3 each) in the in vitro samples and in the cardiac samples with a normal EF (n = 3) or low EF (n = 3).
) at the different time points (8, 24, 48, 72, and 96 h; n = 3 each) in the in vitro samples and in the cardiac samples with a normal EF (n = 3) or low EF (n = 3).
), downregulated ( ), or non-differentially expressed gene ( ) at the different time points (8, 24, 48, 72, and 96 h; n = 3 each) in the in vitro samples and in the cardiac samples with a normal EF (n = 3) or low EF (n = 3).| Gene Symbol | Blood | Plasma/Serum | Urine | Not Detected in Biofluids | In Vitro Expression (8–96 h) | In Vivo Expression (Normal and Low EF) |
|---|---|---|---|---|---|---|
| ACE2 | x | | | |||
| ACKR3 | x | x | | | ||
| ACTN1 | x | x | x | | | |
| CCN1 | x | x | | | ||
| CCN2 | x | x | | | ||
| CLU | x | x | x | | | |
| COL12A1 | x | | | |||
| COL14A1 | x | | | |||
| CORIN | x | | | |||
| DDAH1 | x | | | |||
| FAM155B | x | x | | | ||
| HSPA2 | x | | | |||
| IER3 | x | | | |||
| IGFBP5 | x | x | x | | | |
| IRS2 | x | x | | | ||
| KCNIP2 | x | | | |||
| MFAP4 | x | | | |||
| NES | x | x | | | ||
| NPPA | x | x | | | ||
| NPPB | x | x | | | ||
| PDLIM7 | x | | | |||
| PLSCR4 | x | | | |||
| PPP1R1A | x | | | |||
| RASL11B | x | | | |||
| THBS1 | x | x | x | | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansson, M.; Tangruksa, B.; Heydarkhan-Hagvall, S.; Jeppsson, A.; Sartipy, P.; Synnergren, J. Data Mining Identifies CCN2 and THBS1 as Biomarker Candidates for Cardiac Hypertrophy. Life 2022, 12, 726. https://doi.org/10.3390/life12050726
Johansson M, Tangruksa B, Heydarkhan-Hagvall S, Jeppsson A, Sartipy P, Synnergren J. Data Mining Identifies CCN2 and THBS1 as Biomarker Candidates for Cardiac Hypertrophy. Life. 2022; 12(5):726. https://doi.org/10.3390/life12050726
Chicago/Turabian StyleJohansson, Markus, Benyapa Tangruksa, Sepideh Heydarkhan-Hagvall, Anders Jeppsson, Peter Sartipy, and Jane Synnergren. 2022. "Data Mining Identifies CCN2 and THBS1 as Biomarker Candidates for Cardiac Hypertrophy" Life 12, no. 5: 726. https://doi.org/10.3390/life12050726
APA StyleJohansson, M., Tangruksa, B., Heydarkhan-Hagvall, S., Jeppsson, A., Sartipy, P., & Synnergren, J. (2022). Data Mining Identifies CCN2 and THBS1 as Biomarker Candidates for Cardiac Hypertrophy. Life, 12(5), 726. https://doi.org/10.3390/life12050726