
A Taxonomic Appraisal of Bambusicolous Fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with New Collections from Yunnan Province, China

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection, Examination, Isolation and Preservation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Alignment and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa Names | Strain Numbers | Origin | Substrate/Host | GenBank Accession Numbers | Refs. | ||||
|---|---|---|---|---|---|---|---|---|---|
| LSU | SSU | TEF1-α | RPB2 | ITS | |||||
| Brunneofusispora clematidis | MFLUCC 17-2070 | Chiang Rai, Thailand | Dead stems of Clematis subumbellata | MT214570 | NG_070658 | MT394629 | MT394692 | MT310615 | [7] |
| Brunneofusispora hyalina | MFLUCC 21-0008 | Chiang Mai, Thailand | Decaying wood | MW287234 | MW485613 | MW512606 | MW512609 | MW260330 | [16] |
| Brunneofusispora sinensis | KUMCC 17-0030 | Yunnan, China | Dead wood | MH393557 | MH393556 | MH395329 | / | MH393558 | [5] |
| Brunneofusispora sinensis | MFLUCC 20-0016 | Yunnan, China | Dead branches of Magnolia denudata | MT159624 | MT159636 | MT159607 | MT159613 | MT159630 | [15] |
| Brunneofusispora sp. | X135 | China | Ageratina adenophora | / | / | / | / | MK304223 | [16] |
| Massarina rubi | CBS 691.95 | Austria | Ulmus glabra | FJ795453 | GU456301 | / | FJ795470 | / | Unknown |
| Massarina rubi | MUT 4323 | Italy | Rhizomes of Posidonia oceanica | KF636772 | / | / | / | KF636766 | Unpublished |
| Massarina rubi | MUT 4887 | Italy | Flabellia petiolata | KP671721 | KT587318 | / | / | KR014359 | Unpublished |
| Massarina sp. | MUT 4860 | Italy | Flabellia petiolata | KP671730 | KT587325 | / | / | KR014362 | Unpublished |
| Neooccultibambusa chiangraiensis | MFLUCC 12-0559 | Chiang Rai, Thailand | Dead twigs of Tectona grandis | KU764699 | NG_061230 | KU872761 | / | NR_154238 | [6] |
| Neooccultibambusa jonesii | MFLUCC 16-0643 | Italy | Dead and stems of Ammophila arenaria | NG_059741 | NG_062422 | / | / | / | [9] |
| Neooccultibambusa pandanicola | KUMCC 17-0179 | Yunnan, China | Dead leaves of Pandanus utilis | MG298940 | MG298942 | MG298943 | MG298944 | MG298941 | [17] |
| Neooccultibambusa thailandensis | MFLUCC 16-0274 | Prachuap Khiri Khan, Thailand | Dead leaf of Pandanus sp. | MH260308 | MH260348 | MH412780 | MH412758 | MH275074 | [11] |
| Nigrograna mackinnonii | E5202H | Ecuador | Dead stems of Guazuma ulmifolia | KJ605422 | JX264155 | JX264154 | JX264156 | JX264157 | Unpublished |
| Nigrograna obliqua | MRP | Austria | Ribes uva-crispa | KX650561 | / | KX650532 | KX650581 | KX650561 | [37] |
| Nigrograna obliqua | BW4 | Austria | A twig of Sambucus racemosa | KX650557 | / | KX650529 | / | KX650557 | [37] |
| Occultibambusa aquatica | MFLUCC 11-0006 | Chiang Rai, Thailand | Bamboo | KX698110 | KX698112 | / | / | / | [8] |
| Occultibambusa bambusae | MFLUCC 11-0394 | Chiang Mai, Thailand | Dead culms of bamboo | KU863113 | KU872117 | KU940194 | KU940171 | KU940124 | [4] |
| Occultibambusa bambusae | MFLUCC 13-0855 | Chiang Rai, Thailand | Dead culms of bamboo | KU863112 | KU872116 | KU940193 | KU940170 | KU940123 | [4] |
| Occultibambusa chiangraiensis | MFLUCC 16-0380 | Chiang Rai, Thailand | Dead stems of Bambusoideae sp. | KX655546 | NG_062421 | KX655561 | KX655566 | / | [8] |
| Occultibambusa fusispora | MFLUCC 11-0127 | Chiang Rai, Thailand | Dead branches of bamboo | NG_059669 | / | KU940195 | KU940172 | NR_154340 | [4] |
| Occultibambusa fusispora | MFLUCC 11-0127II | Chiang Rai, Thailand | Dead branches of bamboo | MZ329032 | MZ329028 | MZ325466 | MZ325469 | MZ329036 | This study |
| Occultibambusa hongheensis | KUMCC 21-0020 | Yunnan, China | Dead branches of bamboo | MZ329033 | MZ329029 | MZ325467 | / | MZ329037 | This study |
| Occultibambusa jonesii | GZCC 16-0117 | Guizhou, China | Dead culms of bamboo | NG_066381 | NG_065104 | KY814756 | KY814758 | / | [10] |
| Occultibambusa kunmingensis | KUN-HKAS 102151 | Yunnan, China | Decaying bam | MN913733 | MT864342 | MT954407 | MT878453 | MT627716 | [13] |
| Occultibambusa kunmingensis | KUMCC 21-0019 | Yunnan, China | Submerged bamboo | MZ329034 | MZ329030 | / | / | MZ329038 | This study |
| Occultibambusa maolanensis | GZCC 16-0116 | Guizhou, China | Dead culms of bamboo | KY628323 | KY628325 | KY814757 | KY814759 | / | [10] |
| Occultibambusa pustula | MFLUCC 11-0502 | Chiang Rai, Thailand | Dead culm of bamboo | KU863115 | NG_062419 | / | / | NR_154341 | [4] |
| Ohleria modesta | MGC | Spain | Branches of Chamaecytisus proliferus | KX650562 | / | KX650533 | KX650582 | KX650562 | [37] |
| Ohleria modesta | OM | Spain | Branches of Chamaecytisus proliferus | KX650563 | KX650513 | KX650534 | KX650583 | KX650563 | [37] |
| Seriascoma bambusae | KUMCC 21-0021 | Yunnan, China | Dead culms of bamboo | MZ329035 | MZ329031 | MZ325468 | MZ325470 | MZ329039 | This study |
| Seriascoma didymosporum | MFLUCC 11-0179 | Chiang Rai, Thailand | Dead culms of bamboo | NG_059670 | KU872119 | KU940196 | KU940173 | NR_154433 | [4] |
| Seriascoma didymosporum | MFLUCC 11-0194 | Chiang Rai, Thailand | Dead culms of bamboo | KU863117 | KU872120 | KU940197 | KU940174 | KU940128 | [4] |
| Seriascoma sp. | KUMCC 21-0007 | Yunnan, China | Dead branches of bamboo | MW981347 | MZ325471 | MZ325472 | MZ325473 | MW981351 | [38] |
| Seriascoma yunnanense | MFLU 19-0690 | Yunnan, China | Dead branches of bamboo | NG_068303 | MN174694 | MN381858 | MN210324 | / | [12] |
| Versicolorisporium triseptatum | HHUF 28815 | Honshu, Japan | Dead culms of Pleioblastus chino | NG_042318 | NG_060995 | / | / | NR_119392 | [14] |
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
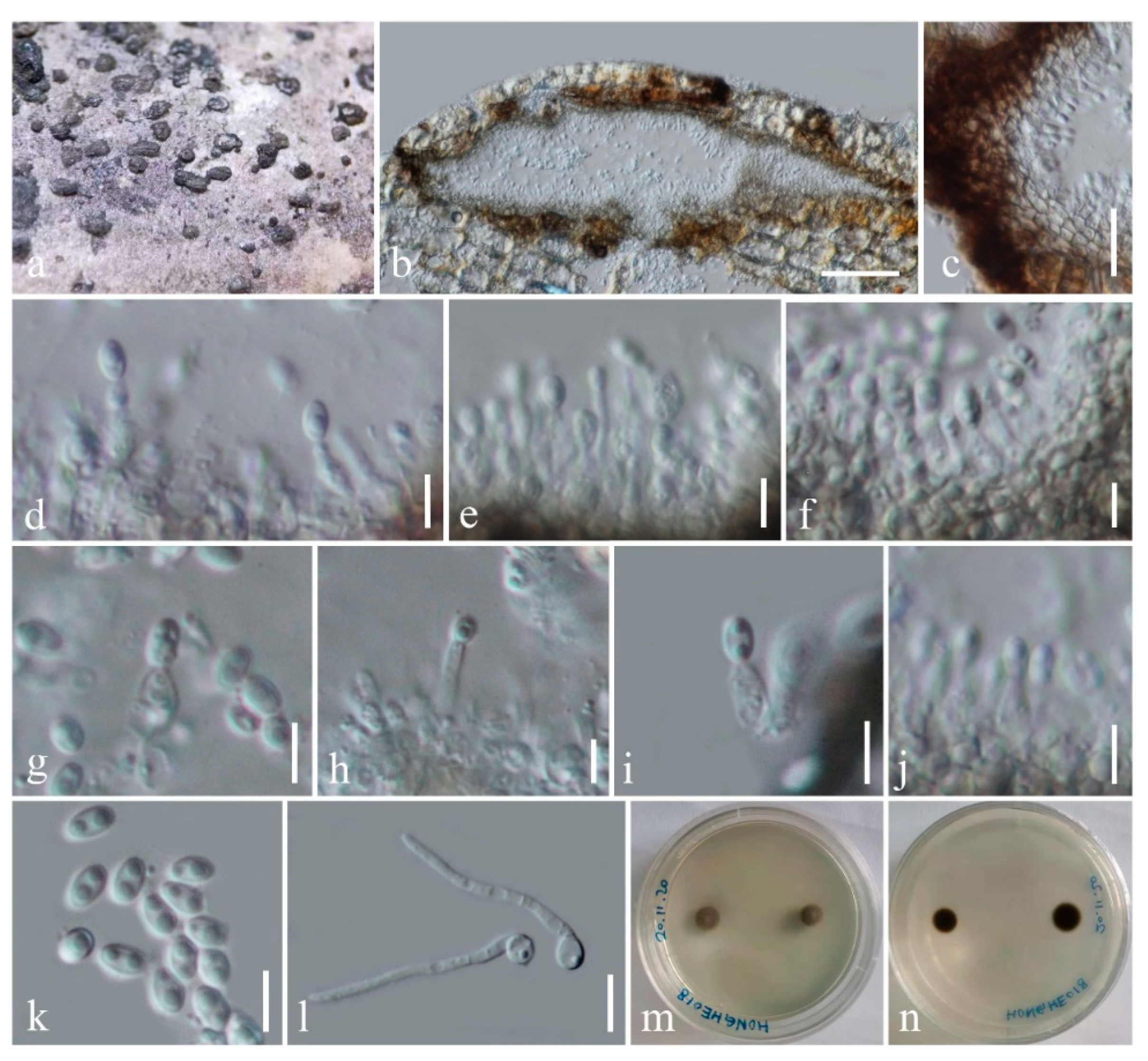
3.2.1. Occultibambusa hongheensis H.B. Jiang, K.D. Hyde & Phookamsak, sp. nov.
3.2.2. Occultibambusa kunmingensis C.X. Liu, H. Zhang & K.D. Hyde in Dong et al., Fungal Diversity 105: 471
3.2.3. Seriascoma bambusae H.B. Jiang, K.D. Hyde & Phookamsak, sp. nov.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, J.D.; Liu, N.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Pem, D.; Hongsanan, S.; Doilom, M.; Tibpromma, S.; Wanasinghe, D.N.; Dong, W.; Liu, N.G.; Phookamsak, R.; Phillips, A.J.L.; Jeewon, R.; et al. https://www.dothideomycetes.org: An online taxonomic resource for the classification, identification, and nomenclature of Dothideomycetes. Asian J. Mycol. 2019, 2, 287–297. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspe’, O.; Karunarathna, S.K.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Camporesi, E.; Kang, J.C. Neooccultibambusajonesii, a novel taxon within Occultibambusaceae. Mycosphere 2016, 7, 1458–1472. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Yang, W.; Liu, Z.Y. Fungi from Asian Karst formations II. Two new species of Occultibambusa (Occultibambusaceae, Dothideomycetes) from karst landforms of China. Mycosphere 2017, 8, 550–559. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Rathnayaka, A.R.; Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Liu, J.K.; Tennakoon, D.S.; Hyde, K.D. Introducing Seriascomayunnanense sp. nov. (Occultibambusaceae, Pleosporales) based on evidence from morphology and phylogeny. Asian J. Mycol. 2019, 2, 245–253. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Tanaka, K.; Harada, Y. Bambusicolous fungi in Japan (7): A new coelomycetous genus, Versicolorisporium. Mycoscience 2008, 49, 211–214. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.C.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef] [PubMed]
- Calabon, M.S.; Jones, E.B.G.; Boonmee, S.; Doilom, M.; Lumyong, S.; Hyde, K.D. Five novel freshwater ascomycetes indicate high undiscovered diversity in lotic habitats in Thailand. J. Fungi 2021, 7, 117. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; de Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Species Fungorum. Available online: http://www.speciesfungorum.org/Names/Names.asp (accessed on 30 January 2021).
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 30 January 2021).
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S. Primers for Elongation Factor 1-alpha (EF1-alpha). 2001. Available online: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf (accessed on 30 December 2020).
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Jiang, H.B.; Hyde, K.D.; Jayawardena, R.S.; Doilom, M.; Xu, J.C.; Phookamsak, R. Taxonomic and phylogenetic characterizations reveal two new species and two new records of Roussoella (Roussoellaceae, Pleosporales) from Yunnan, China. Mycol. Prog. 2019, 18, 577–591. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. Mafft online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. Bioedit Version 6.0.7. 2004. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 1 March 2021).
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the cipres science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest2 v. 2.3 (Program for Selecting DNA Substitution Models Using PAUP*); Evolutionary Biology Centre: Uppsala, Sweden, 2008. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. Genomics 2002, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Jaklitsch, W.M.; Voglmayr, H. Hidden diversity in Thyridaria and a new circumscription of the Thyridariaceae. Stud. Mycol. 2016, 85, 35–64. [Google Scholar] [CrossRef] [Green Version]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, E.B.G.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021. in preparation. [Google Scholar]
- Dai, D.Q.; Tang, L.Z.; Wang, H.B. A review of bambusicolous ascomycetes. In Bamboo: Current and Future Prospects; Abdul Khalil, H.P.S., Ed.; IntechOpen: London, UK, 2018; pp. 165–183. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Zhou, D.Q.; Dalisay, T. Bambusicolous fungi: A review. Fungal Divers. 2002, 9, 1–14. [Google Scholar]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Phookamsak, R.; Norphanphoun, C.; Tanaka, K.; Dai, D.Q.; Luo, Z.L.; Liu, J.K.; Su, H.Y.; Bhat, D.J.; Bahkali, A.H.; Mortimer, P.E.; et al. Towards a natural classification of Astrosphaeriella-like species; introducing Astrosphaeriellaceae and Pseudoastrosphaeriellaceae fam. nov. and Astrosphaeriellopsis, gen. nov. Fungal Divers. 2015, 74, 143–197. [Google Scholar] [CrossRef]




| Species Name | Sexual Morph | Origin | Host | Habitat | References | ||
|---|---|---|---|---|---|---|---|
| Ascostromata | Asci | Ascospores | |||||
| Occultibambusa aquatica | 180–280 × 100–250 μm, subglobose, brown to dark brown, papillate ostiole | 73–86 × 9–13 μm, clavate, with a short furcate pedicel | 19–25 × 3.5–6.5 μm, 2-seriate, narrow fusiform with acute ends, 1-septate, not constricted at the septum, brownish, with sheath | Chiang Rai, Thailand | Submerged bamboo | Freshwater | [8] |
| O. bambusae | 400–550 × 150–200 μm, subglobose, dark brown to black, papillate ostiole | (50–)60–80(−90) × (9.5–)11.5–14.5(−15) μm, broadly cylindrical, with a short furcate pedicel | (22–)23.5–27.5 × 4.5–7 μm, 2–3-seriate, slightly broad fusiform, 1-septate, not constricted at the septum, dark brown, with sheath | Chiang Rai, Thailand | Dead bamboo | Terrestrial | [4] |
| O. chiangraiensis | 352–520 × 195–295 μm, depressed globose to subglobose, brown to light brown, ostiole with a slit-like opening | 47–92 × 12–16 μm, clavate-oblong, with a short pedicel | 16–24 × 5–7 μm, 2-seriate, pale brown to reddish brown, fusiform, tapering towards the ends, (1–)3-septate, not constricted at the septa, without any mucilaginous sheaths and appendages | Chiang Rai, Thailand | Dead stem of Bambusoideae sp. | Terrestrial | [8] |
| O. fusispora | 240–275 × 135–185 μm, conical with wedged sides, brown to dark brown, papillate ostiole | (60–)65–90(−110) × (11–)12–14(−15)(−16) μm, clavate to cylindric-clavate, with a short furcate pedicel | (20–)22–25(−26) × 5–6(−6.5) μm, 2-seriate, fusiform with acute ends, light brown, 1–(2–3)-septate, not constricted at the septa, without any mucilaginous sheaths and appendages | Chiang Rai, Thailand | Dead bamboo | Terrestrial | [4] |
| O. hongheensis | 400–550 × 180–340 µm, ampulliform, conical to subglobose, black, ostiolate | (78–)80–130(–137) × (18–)19–23(–25) μm, cylindrical to clavate, with a short pedicel | (25–)27–30 × (5.5–)8–9(–10) μm, 2-seriate, inequilateral-fusiform, pale brown, 1-septate, slightly constricted at the septum, with a broad mucilaginous sheath | Yunnan, China | Dead bamboo | Terrestrial | This study |
| O. jonesii | 200–260 × 196–236 µm, subglobose, dark brown, papillate ostiole | (65–)75–89(–105) × 13.5–19 µm, broadly cylindrical to clavate, with a short pedicel | 27–33.5 × 5.5–6.5 µm, 1–3-seriate, inequilateral-fusiform, brown to grayish, 1-septate, constricted at the septum, without any mucilaginous sheaths and appendages | Guizhou, China | Dead bamboo | Terrestrial | [10] |
| O. kunmingensis | 220–260 × 110–150 μm, ellipsoidal, black, ostiolate | 110–140(–160) × 13–16.5 μm, cylindric-clavate, with a short to long pedicel | 32–40 × 5–6.5 μm, 3–4-seriate, fusiform, brown, 1-septate, constricted at the septum, without any mucilaginous sheaths and appendages | Yunnan, China | Submerged bamboo | Freshwater | [13] |
| 350–550 × 170–220 μm, ampulliform, dark brown to black, minutely papillate ostiole | (76.4–)83–106(–115) × (11–)12–14(–15) μm, cylindric-clavate to clavate, with a short pedicel or subsessile | (30–)34–36(–37.5) ×(4.5–)5–6 μm, 1–2-seriate, fusiform, brown to dark brown, 1–(3)-septate, slightly constricted at the septum, lacking a gelatinous sheath | Yunnan, China | Dead bamboo | Terrestrial | This study | |
| O. maolanensis | 544–600 µm diam., subglobose to slightly conical, dark brown, papillate ostiole | (66–)77–85(–94) × 17–20(–24) µm, broadly cylindrical to clavate, with a short pedicel | 25–31 × 8–10 µm, 2–4-seriate, inequilateral-fusiform, light brown, 1-septate, slightly constricted at the septum, without any mucilaginous sheaths and appendages | Guizhou, China | Dead bamboo | Terrestrial | [10] |
| O. pustula | 200–300 ×150–200 μm, conical, black, ostiolate | 80–105 × 8–12 μm, cylindrical, with a short furcate pedicel | 22–25 × 5–5.5 μm, 2–3-seriate, slightly broad-fusiform, hyaline to pale brown, 1-septate, not constricted at the septum, with sheath | Chiang Rai, Thailand | Dead bamboo | Terrestrial | [4] |
| 320–350 ×190–220 μm, ellipsoidal, black, papillate ostiole | (60–)78–125 × 12.5–15.5 μm, mostly broadly clavate or sometimes narrowly clavate, with a short or long pedicel | 22–29 × 6–8 μm, 1–2-seriate, fusiform, pale brown and 1-septate when young, dark brown and 3-septate when mature, constricted at the septa, without sheath | Yunnan, China | Submerged wood | Freshwater | [13] | |
| Species Name | Sexual Morph | Asexual Morph | References | ||||
|---|---|---|---|---|---|---|---|
| Ascostromata | Asci | Ascospores | Conidiomata | Conidiogenous Cells | Conidia | ||
| Seriascoma bambusae | N/A | N/A | N/A | 170–380 μm diam., 110–150 μm high, uni- to bi-loculate | 5.6–7.2 × 1.6–3.5 μm | 3.5–4 × 2–2.3 μm, subglobose to ellipsoidal | This study |
| S. didymosporum | 1000–1900 μm diam., 150–320 μm high, multi-loculate | (56–)60–75(−80) × 8–11(−13) μm | 11–12(−14.5) × 3–4 μm, clavate to fusiform, with upper cell shorter and wider than lower cell | 250–470 μm diam., 110–170 μm high, uni-loculate | 4–7(−8) × 1.5–3 μm | 4–5.5 × 1.5–2 μm, oblong, with rounded to obtuse ends | [4] |
| 200–250 μm diam., 120–170 μm high, uni-loculate | 70–95 × 9–11 μm | 10.5–14.5 × 3.5–5 μm, clavate to fusiform, with upper cell shorter and wider than lower cell | N/A | N/A | N/A | [13] | |
| S. yunnanense | 275–400 μm diam., 175–205 μm high, uni-loculate | 44–83 × 10–20 μm | 22–30 × 5–7.2 μm, slightly broad and fusiform, with upper cell larger than lower cell, surrounded by a gelatinous sheath | N/A | N/A | N/A | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.-B.; Phookamsak, R.; Hyde, K.D.; Mortimer, P.E.; Xu, J.-C.; Kakumyan, P.; Karunarathna, S.C.; Kumla, J. A Taxonomic Appraisal of Bambusicolous Fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with New Collections from Yunnan Province, China. Life 2021, 11, 932. https://doi.org/10.3390/life11090932
Jiang H-B, Phookamsak R, Hyde KD, Mortimer PE, Xu J-C, Kakumyan P, Karunarathna SC, Kumla J. A Taxonomic Appraisal of Bambusicolous Fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with New Collections from Yunnan Province, China. Life. 2021; 11(9):932. https://doi.org/10.3390/life11090932
Chicago/Turabian StyleJiang, Hong-Bo, Rungtiwa Phookamsak, Kevin D. Hyde, Peter E. Mortimer, Jian-Chu Xu, Pattana Kakumyan, Samantha C. Karunarathna, and Jaturong Kumla. 2021. "A Taxonomic Appraisal of Bambusicolous Fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with New Collections from Yunnan Province, China" Life 11, no. 9: 932. https://doi.org/10.3390/life11090932