UPLC-MS-Based Metabolomics Profiling for α-Glucosidase Inhibiting Property of Parkia speciosa Pods

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Plant Collection
2.3. Sample Drying and Extraction
2.4. Estimation of α-Glucosidase Inhibitory Activity
2.5. UHPLC-QTOF-MS
2.6. Multivariate Data Analysis
2.7. Statistical Analysis
3. Results
3.1. α-Glucosidase Inhibitory Activity
3.2. UHPLC-QTOF-MS/MS Identification
3.3. Multivariate data analysis (MVDA)
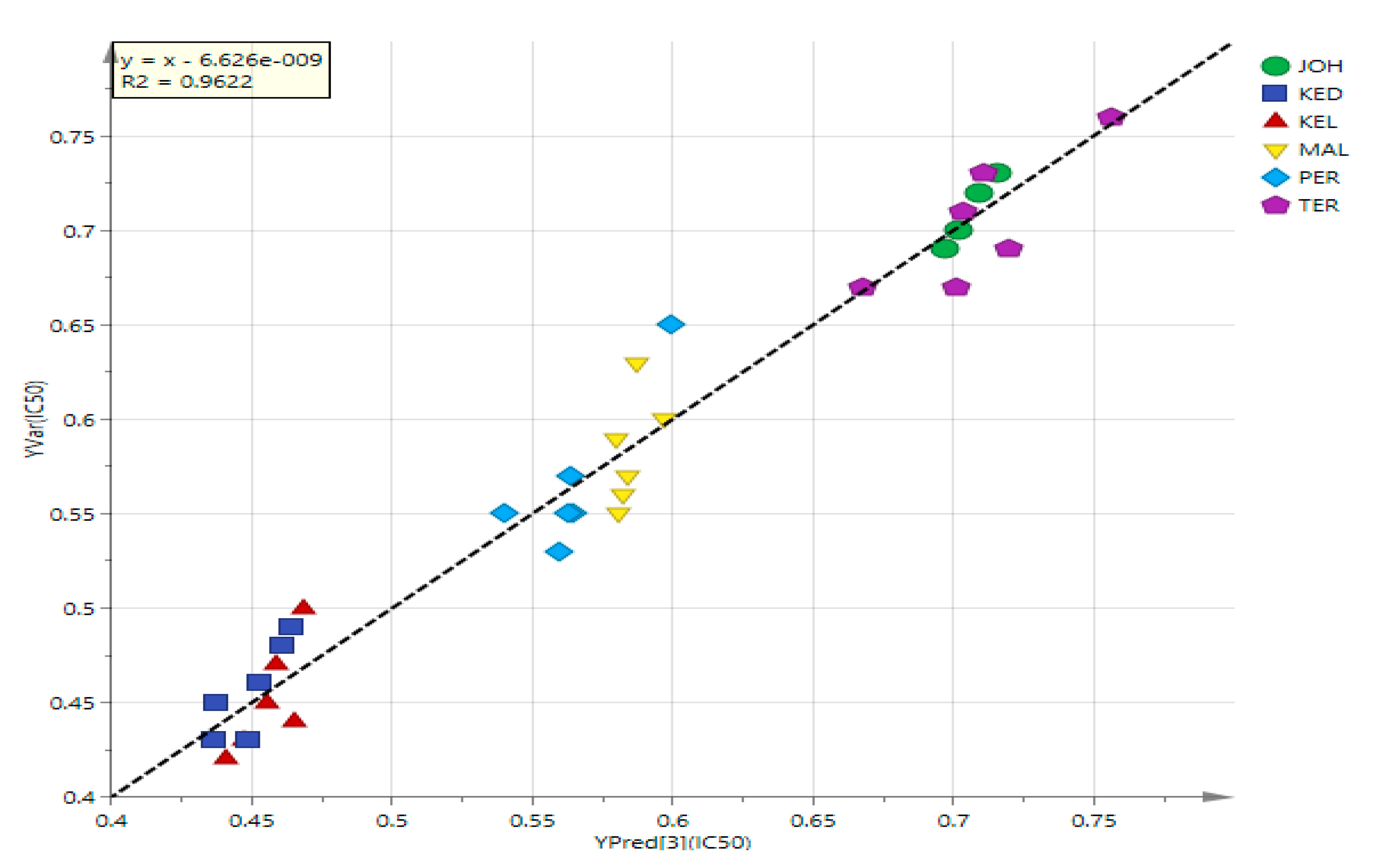
3.4. PLS Analysis of UHPLC-MS Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Definition and Diagnoses of Diabetes Mellitus and Intermediate Hyperglycaemia. Report of a WHO/IDF Consultation; WHO Press: Geneva, Switzerland, 2006. [Google Scholar]
- WHO. Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 30 November 2020).
- Hedrington, M.S.; Davis, S.N. Considerations when using alpha-glucosidase inhibitors in the treatment of type 2 diabetes. Expert Opin. Pharmacother. 2019, 20, 2229–2235. [Google Scholar] [CrossRef]
- Vinholes, J.; Vizzotto, M. Synergisms in alpha-glucosidase inhibition and antioxidant activity of Camellia sinensis L. Kuntze and Eugenia uniflora L. Ethanolic Extracts. Pharmacogn. Res. 2017, 9, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Gerber, P.A.; Rutter, G.A. The role of oxidative stress and hypoxia in pancreatic beta-cell dysfunction in diabetes mellitus. Antioxidants Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.S.M.; Siddiqui, M.J.; Mediani, A.; Ahmed, Q.U.; So’ad, S.Z.M.; Saidi-Besbes, S.; Elnaem, M.H.; Othman, H.A.; Ismail, N.H. Modulation of metabolic alterations of obese diabetic rats upon treatment with Salacca zalacca fruits extract using 1H NMR-based metabolomics. Food Res. Int. 2020, 137, 109547. [Google Scholar] [CrossRef] [PubMed]
- Jamaluddin, F.; Mohameda, S.; Lajis, M.N. Hypoglycaemic effect of stigmast-4-en-3-one, from Parkia speciosa empty pods. Food Chem. 1995, 54, 9–13. [Google Scholar] [CrossRef]
- Trinh, P.T.N.; Giang, B.L.; Tuan, N.T.; Hang, H.T.T.; Thuy, N.T.L.; Tuan, N.N.; Dung, L.T. Alfa glucosidase inhibitory, anti inflammatory activities and a new furanocoumarin derivative of Ruellia tuberosa. Nat. Prod. Res. 2019, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Pham, B.; Le, L. Bioactive compounds in anti-diabetic plants: From herbal medicine to modern drug discovery. Biology 2020, 9, 252. [Google Scholar] [CrossRef] [PubMed]
- Kamisah, Y.; Zuhair, J.S.F.; Juliana, A.H.; Jaarin, K. Parkia speciosa empty pod prevents hypertension and cardiac damage in rats given N(G)-nitro-L-arginine methyl ester. Biomed. Pharmacother. 2017, 96, 291–298. [Google Scholar] [CrossRef]
- Lim, T.K. Parkia speciosa . In Edible Medicinal And Non-Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 798–803. [Google Scholar]
- Saleh, M.S.M.; Jalil, J.; Zainalabidin, S.; Asmadi, A.Y.; Mustafa, N.H.; Kamisah, Y. Genus Parkia: Phytochemical, medicinal Uses, and pharmacological properties. Int. J. Mol. Sci. Rev. 2021, 22, 618. [Google Scholar] [CrossRef]
- Kamisah, Y.; Othman, F.; Qodriyah, H.M.S.; Jaarin, K. Parkia speciosa Hassk.: A potential phytomedicine. Evid. Based Complement. Altern. Med. 2013, 2013, 709028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Bukhori, M.F.M.; Rahmat, M.H.; Rahmat, A. Assessment and comparison of phytochemical constituents and biological activities of bitter bean (Parkia speciosa Hassk.) collected from different locations in Malaysia. Chem. Cent. J. 2018, 12, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, H.J.; Ang, L.H.; Ng, L.T. Antioxidant activities and polyphenolic constituents of bitter bean Parkia speciosa. Int. J. Food Prop. 2014, 17, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Parizad, S.; Dizadji, A.; Habibi, M.K.; Winter, S.; Kalantari, S.; Movi, S.; Lorenzo Tendero, C.; Alonso, G.L.; Moratalla-Lopez, N. The effects of geographical origin and virus infection on the saffron (Crocus sativus L.) quality. Food Chem. 2019, 295, 387–394. [Google Scholar] [CrossRef]
- Saleh, M.S.M.; Siddiqui, M.J.; Mat So’ad, S.Z.; Roheem, F.O.; Saidi-Besbes, S.; Khatib, A. Correlation of FT-IR fingerprint and α-glucosidase inhibitory activity of salak (Salacca zalacca) fruit extracts utilizing orthogonal partial least square. Molecules 2018, 23, 1434. [Google Scholar] [CrossRef] [Green Version]
- Murugesu, S.; Perumal, V.; Balan, T.; Fatinanthan, S.; Khatib, A.; Arifin, N.J.; Shukri, N.S.S.M.; Saleh, M.S.M.; Hin, W.L. The investigation of antioxidant and antidiabetic activities of Christia vespertilionis leaves extracts. South Afr. J. Bot. 2020, 133, 227–235. [Google Scholar] [CrossRef]
- Abd Ghafar, S.Z.; Mediani, A.; Maulidiani, M.; Rudiyanto, R.; Mohd Ghazali, H.; Ramli, N.S.; Abas, F. Complementary NMR- and MS-based metabolomics approaches reveal the correlations of phytochemicals and biological activities in Phyllanthus acidus leaf extracts. Food Res. Int. 2020, 136, 109312. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Mariod, A.A. Wild Fruits: Composition, Nutritional Value and Products; Springer Nature: Berlin/Heidelberg, Germany, 2019; ISBN 9783030318840. [Google Scholar]
- Meng, Y.; Su, A.; Yuan, S.; Zhao, H.; Tan, S.; Hu, C.; Deng, H.; Guo, Y. Evaluation of total flavonoids, myricetin, and quercetin from Hovenia dulcis Thunb. As inhibitors of α-amylase and α-glucosidase. Plant Foods Hum. Nutr. 2016, 71, 444–449. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Lerdsuwankij, O.; Poputtachai, U.; Minipun, A.; Suparpprom, C. Inhibitory activity of cinnamon bark species and their combination effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Plant Foods Hum. Nutr. 2011, 66, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J.; Fern, D.; Mielgo-ayuso, J.; Arora, I.; Sharma, M.; Tollefsbol, T.O.; et al. Dietary polyphenols, Mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxidative Med. Cell. Longev. 2017, 2017, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Stobiecki, M. Application of mass spectrometry for identification and structural studies of flavonoid glycosides. Phytochemistry 2000, 54, 237–256. [Google Scholar] [CrossRef]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Wu, L.; Lin, X.; Hu, X.; Wang, L. Phenolic profiles and screening of potential α-glucosidase inhibitors from Polygonum aviculare L. leaves using ultra-filtration combined with HPLC-ESI-qTOF-MS/MS and molecular docking analysis. Ind. Crop. Prod. 2020, 154, 112673. [Google Scholar] [CrossRef]
- Abd Ghafar, S.Z.; Mediani, A.; Maulidiani; Ramli, N.S.; Abas, F. Antioxidant, α-glucosidase, and nitric oxide inhibitory activities of Phyllanthus acidus and LC–MS/MS profile of the active extract. Food Biosci. 2018, 25, 134–140. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Sari, S.; Šoral, M.; Barut, B.; Özel, A.; Liptaj, T. Potential of Potentilla inclinata and its polyphenolic compounds in α-glucosidase inhibition: Kinetics and interaction mechanism merged with docking simulations. Int. J. Biol. Macromol. 2018, 108, 81–87. [Google Scholar] [CrossRef]
- Roheem, F.O.; Ahmed, Q.U.; So, S.Z.M.; Shah, S.A.A.; Latip, J.; Alhassan, A.M.; Mohammad, S.N.A.S. Assessment of Free radical scavenging and digestive enzyme inhibitory activities of extract, fractions and isolated compounds from Tetracera macrophylla leaves. J. Herb. Med. 2020, 22, 100351. [Google Scholar] [CrossRef]
- Rocchetti, G.; Senizza, B.; Zengin, G.; Senkardes, I.; Bibi Sadeer, N.; Fawzi Mahomoodally, M.; Lucini, L. Metabolomics-based profiling with chemometric approach to delineate the bio-pharmaceutical properties of fruit extracts from Ligustrum vulgare L. Ind. Crop. Prod. 2019, 140, 111635. [Google Scholar] [CrossRef]
- Alhassan, A.M.; Ahmed, Q.U.; Latip, J.; Adnan, S.; Shah, A. A new sulphated flavone and other phytoconstituents from the leaves of Tetracera indica Merr. and their alpha-glucosidase inhibitory activity. Nat. Prod. Res. 2018, 33, 1–8. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Yield (%) | α-Glucosidase Inhibitory Activity (IC50) (µg/mL) |
|---|---|---|
| TER | 16.497 ± 0.702 a | 0.705 ± 0.036 b |
| JOH | 17.393 ± 0.632 a | 0.707 ± 0.016 b |
| KEL | 16.833 ± 1.048 a | 0.443 ± 0.036 d |
| MAL | 16.700 ± 0.605 a | 0.583 ± 0.029 c |
| PER | 16.087 ± 0.784 a | 0.567 ± 0.043 c |
| KED | 16.875 ± 0.996 a | 0.457 ± 0.025 d |
| Quercetin | - | 3.550 ± 0.130 a |
| No. | RT (min) | [M–H]− (m/z) | m/z | MW | Formula | MS/MS | ppm | Tentative Identification | TER | JOH | KEL | MAL | PER | KED |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.196 | 169.0142 | 170.0219 | 170.1200 | C7 H6 O5 | 125.0245, 107.0132 | −2.23 | Gallic acid | + | + | - | - | - | + |
| 2 | 1.705 | 609.1249 | 610.1295 | C30 H26 O14 | 483.0907, 441.0813, 423.0708, 305.0658, 303.0506, 273.0417, 177.0192, 137.0258, 125.0244, 137.0258 | −0.52 | Gallocatechin-(4α→8)- epigallocatechin | + | - | - | + | + | + | |
| 3 | 2.298 | 760.6424 | 716.1379 | 716.6040 | C36 H28 O16 | 609.1227,591.1129, 423.0714, 305.0657 | −2.2 | Theaflavin-3-gallate isomers | - | - | - | + | + | + |
| 2.705 | 1.67 | + | - | + | + | + | + | |||||||
| 2.891 | −2.18 | - | - | - | + | + | + | |||||||
| 5.246 | −1.93 | + | - | + | + | + | + | |||||||
| 4 | 3.419 | 457.0785 | 458.372 | 458.3720 | C15 H14 O7 | 331.0458, 305.0666, 287.0561, 269.0463, 193.0146, 169.0152, 125.0254 | −1.72 | Epigallocatechin | + | - | - | - | - | - |
| 5 | 3.594 | 912.6556 | 914.1546 | 914.7000 | C44 H34 O22 | 761.1333, 743.1231, 591.1131M 573.1024, 423.0714, 177.0198 | −0.02 | Theasinensin A | + | - | + | + | + | + |
| 6 | 4.017 | 457.0778 | 458.0841 | 458.3720 | C22 H18 O11 | 331.0458, 305.0665, 287.0558, 269.0471, 193.0144, 169.0148, 125.0255 | −1.48 | Epigallocatechin gallate isomers | + | - | - | + | + | + |
| 4.305 | −2.96 | - | - | + | + | + | + | |||||||
| 4.505 | −3.18 | + | + | - | + | + | + | |||||||
| 7 | 4.139 | 343.1032 + (HCOO)- | 298.29 | 298.1050 | C14H18O7 | 253.073, 223.0601, 208.0385, 206.021, 195.0657181.0497, 177.02 | 2-Phenylethanol glucuronide | - | + | - | - | - | - | |
| 8 | 6.836 | 479.0814 | 480.0946 | 480.4000 | C21H20O13 | 461.0666, 316.0213, 287.0178, 271.0245, 243.0276, 214.0279, 179.0002, 151.0034, 124.0171 | 1.65 | Gossypetin 8-glucoside | + | + | - | - | - | + |
| 9 | 6.748 | 463.0857 | 464.096 | 464.4000 | C21 H20 O12 | 343.0425, 301.0335, 271.0243, 255.0283, 243.0261, 178.9978, 151.002 | 1.14 | Gossypetin 8-rhamnoside isomers | + | + | - | - | - | + |
| 7.104 | 0.67 | + | + | - | - | + | + | |||||||
| 7.04 | 0.64 | + | + | - | + | - | + | |||||||
| 7.688 | 0.49 | + | + | + | + | + | + | |||||||
| 10 | 7.955 | 463.0882 | 464.0961 | 464.3700 | C21 H20 O12 | 445.0769, 445.0769, 343.0423, 300.0274, 271.0243, 255.0298, 243.0298, 178.9985, 151.0045 | −1.24 | Myricitrin | + | + | - | - | - | - |
| 11 | 8.345 | 595.2027 + (HCOO)- | 550.6020 | 550.6000 | C27 H34 O12 | 595.2022, 475.1579, 431.1332, 369.114, 329.1004, 311.0916, 205.0503, 190.0283 | 1.16 | Eucommin A | - | + | - | + | + | + |
| 12 | 8.408 | 448.1012 | 447.0940 | 447.4000 | C21 H20 O11 | 301.0349, 272.0317, 255.0297, 243.0291, 227.0349, 151.0058, 105.0203 | 0.56 | 6-c-Galactosylluteolin isomers | + | - | - | - | - | - |
| 8.76 | ||||||||||||||
| −0.7 | - | + | - | + | + | + | ||||||||
| 13 | 10.068 | 75.1763 + (M + HCOO) | 530.1778 | 530.5000 | C27 H30 O11 | 559.1445, 279.9182, 170.8336 | 1.92 | Curcumin monoglucoside | + | - | - | - | - | - |
| 14 | 10.088 | 301.0351 | 302.0426 | 302.23OO | C15H10O7 | 273.0399, 257.0437, 245.0428, 192.0079, 178.9975, 164.0087, 151.0037, 121.0286, 107.0143 | 0.19 | Herbacetin | - | + | - | - | - | - |
| 15 | 11.042 | 573.1608 + (M + HCOO)− | 528.1632 | 528.5000 | C27 H28 O11 | 409.0912, 325.0684, 307.0597, 283.0623, 117.0364 | 1.92 | Tremulacin | + | - | - | + | + | - |
| 16 | 13.858 | 293.1753 | 294.1828 | 294.4000 | C17 H26 O4 | 263.1631, 257.1775, 237.1073, 235.1324, 221.1541, 190.1001 | 0.16 | Embelin | - | + | - | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, M.S.M.; Jalil, J.; Mustafa, N.H.; Ramli, F.F.; Asmadi, A.Y.; Kamisah, Y. UPLC-MS-Based Metabolomics Profiling for α-Glucosidase Inhibiting Property of Parkia speciosa Pods. Life 2021, 11, 78. https://doi.org/10.3390/life11020078
Saleh MSM, Jalil J, Mustafa NH, Ramli FF, Asmadi AY, Kamisah Y. UPLC-MS-Based Metabolomics Profiling for α-Glucosidase Inhibiting Property of Parkia speciosa Pods. Life. 2021; 11(2):78. https://doi.org/10.3390/life11020078
Chicago/Turabian StyleSaleh, Mohammed S. M., Juriyati Jalil, Nor Hidayah Mustafa, Fitri Fareez Ramli, Ahmad Yusof Asmadi, and Yusof Kamisah. 2021. "UPLC-MS-Based Metabolomics Profiling for α-Glucosidase Inhibiting Property of Parkia speciosa Pods" Life 11, no. 2: 78. https://doi.org/10.3390/life11020078