
Characterization of New Small-Spored Alternaria Species Isolated from Solanaceae in Algeria
by
 ,
,
Nabahat Bessadat
1,2 ,
,
Bruno Hamon
1,
Nelly Bataillé-Simoneau
1,
Kihal Mabrouk
2 and
Philippe Simoneau
1,* 1
UMR IRHS, University Angers, Agrocampus-Ouest, INRAE, SFR QUASAV, 49071 Beaucouzé, France
2
Laboratoire de Microbiologie Appliquée, Université Oran1 Ahmed Ben Bella, BP1524 El M’naouer, Oran 31000, Algeria
*
Author to whom correspondence should be addressed.
Life 2021, 11(12), 1291; https://doi.org/10.3390/life11121291
Submission received: 31 October 2021
/
Revised: 20 November 2021
/
Accepted: 21 November 2021
/
Published: 24 November 2021
(This article belongs to the Special Issue Biodiversity and Ecology of Fungi in Terrestrial and Marine Ecosystems)
Abstract
:Although large-spored Alternaria species of the section Porri are considered to be the major agents responsible for leaf spot and blight of Solanaceae, small-spored Alternaria species are also frequently isolated from symptomatic tissues. A survey of the north-western regions of Algeria during the 2017–2018 growing seasons revealed that amongst the 623 Alternaria isolates from tomato, potato, pepper, eggplant and black nightshade, 8% could not be morphologically assigned to either section Porri or section Alternaria. In order to more precisely determine the taxonomic position of these isolates, detailed morphological characterizations and multi-locus phylogenetic analyses were performed. Based on these analyses, the isolates were grouped into four main clades: section Ulocladioides, section Infectoriae, including two new species, section Embellisioides, and section Eureka, including one new species. These isolates were also characterized for their virulence under green-house conditions. They were able to produce leaf spot symptoms on tomato plants but with variable levels.
1. Introduction
Alternaria foliar diseases are important factors that decrease yield in solanaceous crops [1,2]. Of these foliar diseases, early blight and brown spot are very common [3,4]. Early blight starts with the formation of small, brownish spots on lower leaves that enlarge and develop concentric rings. Leaves turn yellow and plants may defoliate. Brown spot also initially develops on lower leaves with dark-brown lesions but the foliar lesions never develop the dark concentric rings; instead they coalesce across large veins until whole leaves turn brown and hang from the plant. The pathogens spread progressively to other plants or parts of the same plant such as the stems, fruits and tubers, where they may initiate new infections [5,6,7,8,9].
Many Alternaria species have been documented to cause these foliar diseases on Solanaceae. Among large-spored species belonging to the section Porri, A. solani has long been recognized as the major species responsible for early blight [10,11]. However, it has been shown that A. linariae, A. grandis and A. protenta can also infect both tomato (Solanum lycopersicum L.) and potato (Solanum tuberosum L.) [12,13,14,15]. Moreover, several small-spored species have been reported to cause necrotic lesions on Solanaceae, e.g., A. alternata, A. arborescens, A. tenuissima, A. dumosa, A. interrupta [8,14,16,17,18,19]. However, the exact number of species responsible for Solanaceae leaf spot diseases should be taken with caution due to the recent redefinition of species within the genus Alternaria, based on multi-gene phylogeny. For instance, Woudenberg et al. [20] synonymized some of the small-spored species (A. tenuissima, A. dumosa and A. interrupta) under A. alternata and showed that members of the A. arborescens species constitute a species complex. Nevertheless, concerning tomato, members of at least seven out of the 28 sections described within the genus Alternaria [21,22] have been associated with foliar diseases. Besides the large-spored species of the section Porri and the members of the section Alternaria (A. alternata, A. arborescens, A. tomato), this includes the following species: A. mimicula (section Brassicicola) [23], A. consortialis (section Ulocladioides) [14], A. infectoria (section Infectoriae) [24], A. chlamydosporigena (section Embellisia) [25] and more recently A. telliensis (section Japonicae) [26].
Solanaceae, such as potato and tomato, are the most important crops and the second main source of food in Algeria, after cereals. Annual surveys across the north-western regions of the country revealed the presence of a high rate of Alternaria diseases on several plant species. In fact, early blight is considered to be a major threat for Solanaceae production since A. grandis and A. protenta were observed on tomato and A. linariae on potato for the first time [27,28,29]. The north-western regions of Algeria are characterized by a warm Mediterranean climate (Köppen climate classification) where tomato can be cultivated throughout the year [30]. In these regions, other solanaceous cultivated plants, i.e., potato, eggplant (Solanum melongena), and pepper (Capsicum annuum), are often found in the near vicinity of tomato fields or even used in crop rotation with tomato. Climatic conditions and agricultural practices are therefore highly favorable for the development of epidemics and the appearance of new virulent genotypes. Thus, the composition of Alternaria species that is associated with tomato foliar diseases is probably rapidly evolving in Algeria and may be more complex than previously recognized. Moreover, except for the members of the section Alternaria [18,31], small-spored Alternaria spp. have been poorly studied and were the primary focus of this study. The objective was thus to estimate the diversity of small-spored species associated with necrotic lesions on Solanaceae in Algeria using morphological, multi-locus phylogenetic and pathological approaches. This study revealed that small-spored isolates belonging to four sections of Porri, other than section Alternaria, were isolated from symptomatic tissues of Solanaceae in northern Algeria.
2. Materials and Methods
2.1. Sampling and Isolation
The current study was part of a long-term project on the species diversity of the Solanaceae–Alternaria pathosystem in the north-west region of Algeria [14]. During the 2017 and 2018 growing seasons, samples with typical symptoms of early blight and brown spot were collected from several solanaceous plants in the cropping area of the Mostaganem regions. Weeds such as black nightshade (Solanum nigrum) and jimson weed (Datura stramonium) with suspected foliar symptoms of Alternaria infection were also collected (Table 1). Samples were separately placed in paper bags and transported to the laboratory. The isolation of fungi was performed immediately. Small pieces (1 cm2) of both diseased and apparently healthy vegetable tissue (leaf, stem, fruit) were collected, and their surfaces were disinfected by dipping them in 2% sodium hypochlorite solution for 2 min and rinsing them with sterile distilled water three times. Samples were placed in Petri plates containing potato carrot agar medium (PCA) and incubated at room temperature in ambient light. Multiple fungal colonies that were growing up from the margins or surfaces of each lesion produced conidia within one to three weeks. These colonies were examined under a light stereomicroscope at 20× and 40× and further transferred onto new PCA plates. The colonies with Alternaria characteristics were purified by transferring single spores onto fresh potato dextrose agar medium (PDA) plates. After adequate growth, mycelial plugs of these isolates were cut and stored in 30% glycerol at −80 °C to maintain a fungal collection.
2.2. Species Identification and Morphological Characterization
Pure cultures of Alternaria isolates with different morphologies and representing small-spored species were preliminarily identified according to the standard method [23]. Isolates were cultured on 9 cm PCA plates and incubated at 22–28 °C under a daily fluorescent 8 light/16 dark cycle for 5 to 7 days. After this period, the arrangement of conidia on conidiophores and the patterns of chain branching were examined at 40× and 100× without disrupting the colony. For the examination of microscopic details, mounts were prepared in pure lactic acid without added dye. The conidial color, shape, size (length and width) and number of septa (longitudinal, oblique or transverse), as well as the presence or absence of secondary conidiophores were recorded from observations that were made at 400X magnifications. Dimensions are based on the observation of 50 conidia and 25 conidiophores per isolate. In general, sporulation patterns were compared with the representative species described in the literature (Simmons 2004; 2007) or with the reference strain of A. cumini (CBS 121329). For macro-morphological characterization, the color of the upper and reverse side of the colonies, colony margin, texture and diameter were recorded on PCA and PDA. Color notations were rated according to the color charts of Kornerup and Wanscher [32]. New species were also described on PDA, Oatmeal Agar (OA) and Malt Extract Agar (MEA). The effect of temperature on growth was studied at different temperatures, i.e., 4 °C, 25 °C, 30 °C, 35 °C and 40 °C. Petri dishes (Ø = 90 mm) containing 15 mL of each media (PCA, PDA, MEA, OA) were inoculated with 5 mm mycelial discs of 15 days-old culture. Colony diameter was measured after 7 days of incubation. Specimen for each new species (NB65, NB530, NB560, NB562, NB568, NB660) were deposited in the Westerdijk Fungal Biodiversity Institute (Utrecht, The Netherlands) under the reference codes CBS 146567, CBS 148413, CBS 148415, CBS 148414, CBS 148416, and CBS 146566, respectively. Nomenclatural data were deposited in MycoBank (MB 835037: A. cuminicola, MB 840829: A. dahraensis, MB 840845: A. pseudohumuli).
2.3. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification and Sequencing
Alternaria isolates were grown on PDA for 7 days; mycelium was scraped from the culture surface and transferred into a microtube. Lysis buffer (50 mM Tris-HCl pH 7.5, 50 mM EDTA, 3% SDS, 1% 2-mercaptoethanol) was added and the nucleic acids were extracted according to the microwave mini-prep procedure described by Goodwin and Lee [33]. The final DNA pellet was dissolved into 100 μL TE buffer (10 mM Tris-HCl pH 8.0, 0.1 mM EDTA).
As an initial step of identification, the internal transcribed spacer region of ribosomal DNA (ITS rDNA) and a portion of the glyceraldehyde-3-phosphate dehydrogenase (gpd) gene were amplified from the isolates with the primers ITS1/ITS4 [34] and gpd1-gpd2 [35], respectively. Since it was impossible to discriminate between closely related species solely by using the ITS rDNA and gpd sequences, portions of the genes of the translation elongation factor 1-alpha (tef1), RNA polymerase’s second largest subunit (rpb2), plasma membrane ATPase (ATPase), actin (act), a major Alternaria allergen (Alt a1), and a mating type protein (MAT1-2-1) were amplified with the primer pairs EF1-728F/EF1-986R [36], RPB2–5F2/fRPB2–7cR [37,38], ATPDF1/ATPDR1, ACTDF1/ACTDR1 [39], Alt-for/Alt-rev [40] and MAT1-2F/MAT1-2R [41], respectively. PCR amplification was carried out in a total volume of 50 μL containing 75 mM Tris-HCl pH 9.0, 20 mM (NH4)2SO4, 0.01% (w/v) Tween 20, 1.5 mM MgCl2, 200 μM of each deoxyribonucleotide triphosphate, 1 unit of thermostable DNA polymerase (GoTaq, Promega) and 400 nM of each relevant oligonucleotide primer. The thermocycling parameters were the same as described in the references provided above. DNA amplification products were analyzed by electrophoresis in 1.2% (w/v) agarose in 0.5 × TAE buffer and sent for sequencing at GATC Lab (Germany).
2.4. Sequence Alignment and Phylogenetic Analysis
DNA sequences were concatenated and aligned by the MUSCLE algorithm using MEGA 7 [42]. Phylogenetic analysis was performed using the maximum likelihood (ML) and Bayesian inference (BI) approaches under IQTree v.1.6. [43] and MrBayes v.3.2.1 [44], respectively. The best-fit evolutionary models for each dataset were calculated by ModelFinder [45] under the Bayesian Information Criterion (BIC) selection procedure. The ML analysis was carried out with 1,000 ultrafast bootstrap replicates and only values above 70% were considered significant. BIs were performed in order to estimate the posterior probabilities (PP) of tree topologies based on the Markov Chain Monte Carlo (MCMC) analysis with four chains, 1M generations, sampling intervals of 1,000 generations. Burn-in was set to 25% and only PP values above 0.95 were considered significant.
2.5. Pathogenicity Tests
In order to determine the virulence of the isolates, representative isolates selected from among the identified phylogenetic taxa were used (Table 1). Conidia of these isolates that were produced on 7 to 10 day-old PCA cultures were used to prepare conidial suspensions. Conidia were collected by rubbing them gently from the surface of the medium with a rubber spatula and placing them in a test tube with sterile distilled water containing 0.01% Tween 80; the conidial suspensions were then filtered through a mousseline membrane to remove mycelial fragments. Concentrations of the spore suspensions were estimated using a hemocytometer and adjusted to 105 conidia per mL. Tomato var. Saint Pierre and potato var. Désirée were used in this study. Disinfected tomato seeds and potato tubers were grown, after germination, in plastic pots containing sterilized soil (3/4 potting soil, 1/4 sand mixture). Six Apiaceae (coriander, carrot var. Super Muscade, parsley, fennel, celery, cumin) plant species were included in the pathogenicity tests in order to assess the host range of some isolates from section Eureka. Three plants from each species were used in this study. The plants were kept in a greenhouse for six weeks. Greenhouse temperatures were approximately 25–32 °C during the day and 19–25 °C at night. Inoculation was done by spraying the plants with 10 mL of the spore suspension. Five strains, three from section Alternaria (A. alternata, A. arborescens), one from section Eureka (A. cumini, CBS 121329) and one from section Radicina (A. petroselini CBS 109383) were used as positive controls on tomato and Apiaceae plants, respectively. The negative controls were sprayed with sterile distilled water. Plants were covered with polyethylene plastic bags for two days to maintain a high level of humidity. The percentage of the leaves’ surfaces that were covered with dark lesions was estimated for each plant 7, 14 and 21 days after inoculation (DAI). Each inoculation experiment was done in triplicate.
3. Results
3.1. General Survey
A large survey was carried out in 2017 and 2018 in the Mostaganem region (Algeria) on symptomatic leaves of different cultivated solanaceous crops (tomato, eggplants, potato, pepper) or neighboring wild species belonging to the Solanaceae (jimson weed Datura stramonium, black nightshade Solanum nigrum). A total of 367 leaf samples representing brown spot and early blight symptoms were collected, and 1061 isolates were obtained and observed for their conidia size and morphology. Based on these observations, the isolated fungi were classified into four categories: small-spored Alternaria spp. (48%), large-spored Alternaria spp. (11%), Stemphyllium spp. (33%) and fungi belonging to other genera (8%). Among the small-spored Alternaria species, 92% had typical sporulation patterns of Alternaria sect. Alternaria and were present in all sampling locations, while 8% could not be unambiguously identified at the section level based only on their morphological characteristics. A selection of 20 small-spored isolates that produced conidia that were different from the isolates of the members of the section Alternaria were subjected to monospore purification for further characterization (Table 1). The initial characterization of isolates from this collection was performed by sequencing the ITS rDNA and gpd loci. The latter locus allowed a good resolution of the 28 sections described in the genus Alternaria. The resulting phylogenetic tree (Figure 1) revealed that the isolates belonged to four sections: Ulocladioides, Infectoriae, Embellisioides and Eureka.
3.2. Species Identification
Further identification of these isolates was obtained through multilocus sequence analyses and detailed morphological characterization.
3.2.1. Section Ulocladioides
The two isolates assigned to this section (NB558, NB559) shared similar morphological characteristics. They both had a slow radial growth rate (ca. 8.67 ± 0.89 mm/day on PDA medium) and formed dark green, powdery colonies. Microscopic observations (Figure 2a,b) showed that they produced obovoid, beakless, small-size conidia (mean length and width: (15)25–36(41) µm and (8)10–13(15) µm, respectively), with 1–5 transverse septa and 0–4 longitudinal septa. Conidiophores arose near the agar surface and were (25)30–50(77) µm long and 3–4(5) µm wide and had 1–13 successive, apical sympodial conidiogenous sites. Secondary conidiophores were scarcely formed. A multilocus phylogeny based on gpd, Alt a1 and MAT1-2-1, which has been previously used to resolve the phylogenetic relationships between species that were formerly included in the genus Ulocladium [48], was selected to identify the two isolates at the species level. Sequences obtained for NB558 and NB559 at these 3 loci were compared with those from 16 of the 21 recognized species belonging to section Ulocladioides [48,49], for which the sequences were available in GenBank. Sequences of an isolate of A. alternata (CBS 916.96) were used as the outgroup. The combined gene dataset included 1,849 characters (including gaps) and 135 were parsimony-informative. The best-fit model (TN+F+I+G4) was selected for the analysis. The topology of the resulting trees that were inferred from the two phylogenetic methods (ML and BI) was similar. The ML phylogenetic tree shown in Figure 3 did not allow for the complete separation of all of the species within section Ulocladioides, and the isolates NB508 and NB509 could not be separated from two species, i.e., A. cucurbitae and A. brassicae-pekinensis. Additional attempts to resolve this group of strains by including two additional loci (rpb2 and tef1) were unsuccessful (supplementary Figure S1).
3.2.2. Section Infectoriae
Twelve of the small-spored isolates considered here were included in the section Infectoriae based on gpd phylogeny (Figure 1). They all shared morphological characteristics that are typical for members of this section, i.e., they often have mostly mycelial colonies in the culture, with poor sporulation until hyphae were scarified, conidiophores with several conidiogenous loci, resulting in a three-dimensional sporulation pattern, and many conidia with elongated apical regions (“false beaks”) [50]. Primary conidiophores were mostly simple or branched ((12)30–75(130) × 3–4(5) µm) and had 1–4 conidiogenous loci. Conidia were obclavate, ellipsoid to ovoid and reached a size of (15)25–44(72) × (6)8–12(17) µm with 2–11 transverse septa and 0-5 longitudinal septa (Figure 2c–g). However, due to the high similarity between isolates, it was not possible to delineate reliable morpho-groups based solely on morphology within the members of this section. Phylogenetic analyses at five loci (ITS rDNA, gpd, rpb2, tef1 and ATPase) have been used to separate species within the section Infectoriae [51,52,53,54,55]. Sequences were obtained at these five loci for the 12 isolates assigned to section Infectoriae and compared with corresponding sequences from 37 recognized species from section Infectoriae. Sequences of A. abundans CBS 534.83 (section Chalastospora) were used as the outgroup. The combined dataset included 2,566 characters (including gaps) and 193 were parsimony-informative. The best-fit evolutionary model was TN+F+R4. The resulting ML tree confirmed that all of the isolates clustered in section Infectoriae (Figure 4). However, most of the isolates appeared as lineages distinct from the 37 types of strains of the species included in this study and independently formed separated branches (e.g., NB561, NB540, NB539, NB538, NB550). Isolates NB562, NB530 and NB545 clustered together, forming a highly supported clade that was distinct from all the other strains included in that analysis. Similarly, NB560 and NB568 grouped in a clade together with A. humuli. However, the phylogenetic relationship between these two isolates and this species was uncertain due to the low support of internal branches. Based on these observations, we propose two new species in the section Infectoriae named Alternaria dahraensis and Alternaria pseudohumuli, respectively (see description below in the “Taxonomy” paragraph).
3.2.3. Sections Embellisioides and Eureka
As shown in Figure 1, six isolates, all collected from the Mostaganem region, grouped into two phylogenetically related sections, i.e., section Embellisioides and section Eureka, that both contain species formerly assigned to the genus Embellisia [21,46]. Five isolates (NB65, NB542, NB557, NB660, NB666), clustering with members of the section Eureka based on gpd phylogeny, shared similar morphology. These isolates developed greyish green, velvety colonies and produced short, beakless, ovoid conidia ((12)25–42(52) × (6)10–20(25) µm). The conidia were formed on long primary conidiophores ((155)430–760(1097) × 3-5(6) µm). These isolates usually produced short conidial chains (3–4 units) and secondary conidiophores ((17)25–55(158) × 3–5 µm), which arose from apical, lateral or basal cells of the conidia. The tertiary conidiophores were common and produced single conidia. Sporulation patterns were almost the same compared with the A. cumini reference strain CBS 212,329, with the exception of the conidia size ((30)35–50(70) × (12)15–17(21) µm) and the abundance of tertiary conidiophores. Isolate NB354 had different morphological characteristics (Figure 2h) and produced ellipsoid, ovoid, conical or cylindrical conidia with broadly rounded bases (mean length and width (30)40–48(61) × (10)12–15(25) µm, respectively). Conidia were solitary (rarely in short chains of 2–3 spores) or produced in a bouquet, with mostly transverse septa (4–8) that distinctly contrasted with the external wall. Conidiophores were of moderate length, (32)45–57(89) µm, each bearing 1–8(14) conidiogenous sites. These morphological characteristics were in agreement with the assignation of this isolate to the section Embellisioides. In order to better resolve these isolates at the species level within their respective sections, a phylogenetic analysis based on additional loci was performed. In accordance with Woudenberg et al. [46], who first resolved the taxonomic position of strains that were formerly grouped in the genus Embellisia, a combined gpd-rpb2-tef1 phylogeny was used. Sequences were obtained at these three loci for the six isolates that were assigned to sections Eureka and Embellisioides and compared with the corresponding sequences from the 12 recognized species from these sections. Sequences of A. embellisia CBS 339.71 (section Embellisia) were used as the outgroup. The combined dataset included 1,533 characters (including gaps) and 217 were parsimony-informative. The best-fit evolutionary model was TNe+G4. The resulting ML tree (Figure 5) revealed that isolate NB354 formed a well-supported branch close to A. lolii within section Embellisioides. As the morphology of this isolate matched the published description of Embellisia lolii from E.G. Simmons & C.F. Hill [56], isolate NB354 was considered to be a member of this species. By contrast, the five isolates belonging to section Eureka formed a separate clade that was closely related to A. cumini. Comparison of sequences from these isolates with those from the reference strain of A. cumini at additional loci (Act, ATPase, Alt a1) confirmed that they formed a distinct cluster (Figure S2). As described above, discrete morphological characteristics distinguished these isolates from A. cumini and they were therefore considered to be a new species named Alternaria cuminicola (see detailed description below in the “Taxonomy” paragraph).
3.2.4. Taxonomy
- MycoBank: MB 840829
Etymology: name refers to the Dahra range located in the northern part of Algeria where the fungus was isolated.
- Culture ex-type specimen: NB 530 = CBS 148413
- Other specimen examined: NB545, NB 562 = CBS 148414
Description: Colonies on PCA are subhyaline to brown, arachnoid to loosely wooly, approximately 76 mm diameter at day 7 (Figure 6i); abundant sporulation but not densely crowded at the aerial hyphae within the 3 cm central radius of the NB350 colony. However, colonies of strain NB562 were mostly mycelial, with a meager sporulation. Zones of growth and sporulation were not well defined. Aerial arachnoid hypha arose early over the center of the colony and gradually over the entire colony; they mainly produced scattered chains of conidia. Conidia chains that were viewed at 40X had a conspicuous appearance of open-angled tufts of 7–15(34) conidia. Chains were simple in young colonies (3–6 conidia) (Figure 6k); they became branched with open angles to the primary chain upon aging (Figure 6m). These chains included 1–2(3) lateral branches of 1–3(5) conidia. Primary conidiophores that arose near the agar surface were 57 ± 25 × 5 ± 1 (15–113 × 4–6) µm in size, with 3–8(18) transverse septa, were usually simple, occasionally branched, and incorporated 1–3 conidiogenous loci. Secondary conidiophores were usually short and often (but not always) the dominant element in the chain structure; they were frequently longer than conidium body (Figure 6k). These lateral or apical secondary conidiophores (Figure 6b,e) were single-celled; they had either straight, geniculate, or angular extensions ca 56 ± 25 × 5 ± 1 (5–90(118) × 4–7) µm, with 1 to 3 conidiogenous loci. The apical or lateral secondary conidiophore of the primary conidia was as long as the primary conidiophore and became conidiogenous. Simple secondary conidiophores of exceptional length were often present within most of aerial spore clusters. Conidia were ovoid or ellipsoid, medium brown, smooth or ornamented, and each was beakless with either a bluntly tapered or conical apical cell (Figure 6a,d). Longitudinal septa were common within the total maturing population. The largest conidia in the mature colony were mostly basal in chains near 39 ± 10 × 11 ± 2 (18–68 × 8–23) µm with 3–8(10) transverse septa and 1–4 longitudinal septa, although some short, ovoid, primary conidia were present. Most secondary conidia were of the same length or shorter ca 36 ± 6 × 14 ± 2 (23–53 × 9–18) µm. A large number of conidia remained small within the chains (Figure 6f); they were commonly in the range of 29 ± 7 × 14 ± 2 (19–45 × 10–20) µm, with 1–4(6) transverse septa and 0–4 longitudinal septa.
- MycoBank: MB 840845
- Etymology: name refers to the closely related species A. humuli.
- Culture ex-type specimen: NB 560 = CBS 148415
- Other specimen examined: NB 568 = CBS 148416
Description: the colonies on PCA were often mostly mycelial, hyaline to subhyaline, loosely wooly, and reached 66 mm diameter after 7 days (Figure 7j); sporulation was meager or completely lacking until hypha were disturbed or scarified. Zones of growth and sporulation were not well defined. Clusters of a few conidia (up to 5 to 15) were already scattered on the aerial hyphae above the center of the colony after 4 days. By day 14, discrete clusters of openly branched conidial chains were produced on the agar substrate near the center of the colony, resulting in bushy clumps (Figure 7d,i). Primary conidiophores were solitary, straight or geniculate, brown, simple, occasionally branched, short ca 25–82 × 3–5 (53 ± 15 × 4 ± 1) μm with 1–7 transverse septa; some were long (with up to 8 transverse septa) ca 63–133 × 3–7 (95 ± 18 × 5 ± 0.5) μm, with one or several (2–6) conidiogenous loci (pores) at the apex and throughout their lengths (Figure 7i,l). Conidial chains were moderately long to long 4–7(13) and usually branched. Branching was produced by the occurrence of several conidiogenous loci on apical secondary conidiophores (Figure 7b,e) and lateral secondary conidiophores (Figure 7h); lateral branches of 2–3(5) conidia were present and the dominant portion of the sporulation pattern (in contrast to the pattern in A. dahraensis isolated from eggplant). Aged clumps of conidia that developed on a single conidiophore were comprised of 40–50(68) conidia (Figure 7d). Secondary conidiophores were usually short and not the dominant element in the chain structure (except on water agar substrate); they were occasionally longer than the conidium body 14–80 × 3–7 (42 ± 16 × 4 ± 1) μm, with 2–8(10) transverse septa that mainly formed at the beginning of the chain (Figure 7e). These lateral or apical secondary conidiophores were single and had straight, geniculate or angular extensions. Juvenile conidia were ovoid and short-beaked with a tapered apex, or with a bluntly rounded base and apex containing 1–2(3) septa (Figure 7c). Mature conidia were narrowly oblong and ellipsoid with a rounded apex, pale brown or yellowish to dark brown, and the spore wall was either smooth or conspicuously ornamented (Figure 7a). Primary conidia at the base of the chain were predominately long-ellipsoid 28–75 × 6–18 (46 ± 12 × 12 ± 3) μm in size; with 5–7(9) transverse septa and at most one longitudinal septum in 1–3 of the transverse segments, which were slightly constricted near some transverse septa. Mature conidia at the beginning of the chain often produced secondary conidiophores (Figure 7b). The conidia in chains often showed a gradual decrease in size from relatively large and multi-cellular near the base of the chain to small and 1–3 celled near the apex. Most of the secondary conidia were the same length or shorter ca 29 ± 8 × 10 ± 3 (15–50 × 6–18) µm, with 1–4(8) transverse and 0–5 longitudinal septa. Other conidia were shorter ca 26 ± 6 × 9 ± 2 (17–42 × 6–15) µm and produced at least 3 transverse septa in the conidium body, terminating in a chain that was ovoid or ellipsoid and had no secondary conidiophores, beaks or conidiogenous loci on the apical cell.
Note: the most obvious difference between A. humuli and A. pseudohumuli was found in primary conidiophores; in A. humuli, they became variously branched upon aging and bore 8–10 conidiogenous sites [23], while in A. pseudohumuli they remained simple, geniculate, or sometimes produced two branches with only a few conidiogenous sites. However, secondary conidiophores of exceptional length within A. humuli conidial clusters were also present in A. pseudohumuli at a few positions but in most of the conidial clusters. Conidia size constitutes another distinction between the two species. A. humuli produced smaller, more mature conidia (28–38 × 10–11 µm) [23] than A. pseudohumuli.
- MycoBank: MB 835037
- Etymology: name refers to the closely related species A. cumini but with smaller conidia.
- Culture ex-type specimen: NB 65 = CBS 146567
- Other specimen examined: NB666, NB660, NB542, NB557
Description: the colonies on PCA were velvety, approximately 60 mm in diameter without concentric rings of growth and sporulated according to standardized conditions [23] (Figure 8f). At 7 days, sporulation occupied the center of the colony in a small area (1.5 cm diameter with NB65); however, many non-sporulating vegetative elements above the substrate surface were observed. Conidiophores that were produced in the central area were up to ca 370–770 × 3–5 µm, often developing further through geniculate extensions (Figure 8k). Development outside the center of the colony consisted of moderate primary conidiophores ca 106–350 × 4–5 µm (7–24 transverse septa), with few lateral branches, bearing a single apical conidium. Primary conidiophores were abundant and became densely crowded upon aging (up to 14 days) reaching a size length of 820–1100 × 4–5 µm (Figure 8j), with 3–6 lateral branches (22–190 × 3.5–5 µm). Each conidiogenous apex frequently bore a chain of 2–4(5) conidia, mainly with additional short lateral or basal chains of 2–3 conidia forming clumps of 12–40 conidia. Chain branching typically occurred by the elongation of secondary conidiophores (ca 15–50 × 3.5–5 µm with 2–5 transverse septa) from distal terminal conidial cells (Figure 8c), and lateral ones (Figure 8i) and subsequently formed numerous secondary, tertiary, and quaternary chains upon aging. Conidia that were produced on PCA were brown, short ellipsoid, ovoid, and beakless with a broad rounded or conical apex. The dominant size range was 20–29 µm long × 12–20 μm width with 2–4 transverse septa and 0–4 longitudinal septa. Conidia that were produced in the aerial branches tended to be small ca 6.5–25 × 5–18 µm with 0–3 transverse septa and 0–3 longitudinal septa (Figure 8b). Maturing conidia reached a maximum size of 30–38 × 12.5–20 μm with 2–5 transverse septa and 1–4 longitudinal septa (Figure 8a). They became distinctly constricted at the main two transverse septa and had a smooth or sometimes punctate surface. The outer walls and some transverse and longitudinal septa became thick and darkly pigmented.
Note: A. cuminicola sp. nov. isolates were separable at high magnifications from A. cumini reference strain CBS 121329. They clearly exhibited different sporulation patterns and conidial morphology under recommended conditions [23]. Conidia that were formed by A. cuminicola sp. nov. were smaller and the sporulation axes with conidia branching were more complex in structure, forming dense tufts that were visible to the naked eye in the center of the colony at 7–14 days.
3.3. Effect of Culture Media and Temperature on Colony Morphology and Growth
The effect of media and temperature on colony morphology and growth of the three newly described species was determined by the colony diameter method using PCA, PDA, MEA and OA medium. Growth was recorded at five temperatures (from 4 °C to 40 °C). Colony aspect and color varied with the type of culture medium. All fungi grew best on PCA and PDA at 25 °C (Table 2). In general, no growth was observed at 40 °C on any media except for NB530 (A. dahraensis) on PDA, MEA, OA and NB568 (A. pseudohumuli) on OA at 40 °C.
3.4. Pathogenicity Tests
Tomato plants were inoculated with 18 Alternaria isolates from the sections Infectoriae, Ulocladioides, Eureka and Embellisioides. Three isolates from the section Alternaria, i.e., A. alternata (isolates NB529 and NB553) and A. arborescens (NB555) were used as positive controls. However, regardless of the isolate origin or identity, similar necrotic symptoms were observed at 21 DAI on all inoculated plants, but with various intensities (Figure 9). No symptoms were seen in the negative control plants. The lesions initially developed on the basal leaves, the tips and along the margins of the leaf petiole. Symptoms also caused the development of brown, circular spots that turned into dark brown, necrotic spots surrounded by yellowish margins within 7 DAI. The infected lower leaves often began yellowing and browning before falling out. Lesions that were produced by aggressive isolates enlarged and coalesced, causing a blight of the leaves and progressing upwards. Two isolates from section Infectoriae (NB538, NB562) had a high severity on both tomato plants inducing severe blight and spot symptoms at 21 DAI (Figure 9A). The percentage of the leaf necrotic area on tomato plants was slightly higher with the reference isolates from section Alternaria (32.17 ± 9.53%) compared with those from section Infectoriae (16.67 ± 9.30%). Isolates belonging to sections Eureka and Embellisioides had the lowest severity (Figure 9B). Similar trends of aggressiveness were recorded for the potato plants, although in this case five strains from the section Infectoriae were found to be more aggressive than the two A. alternata strains that were used as positive controls (Figure S3).
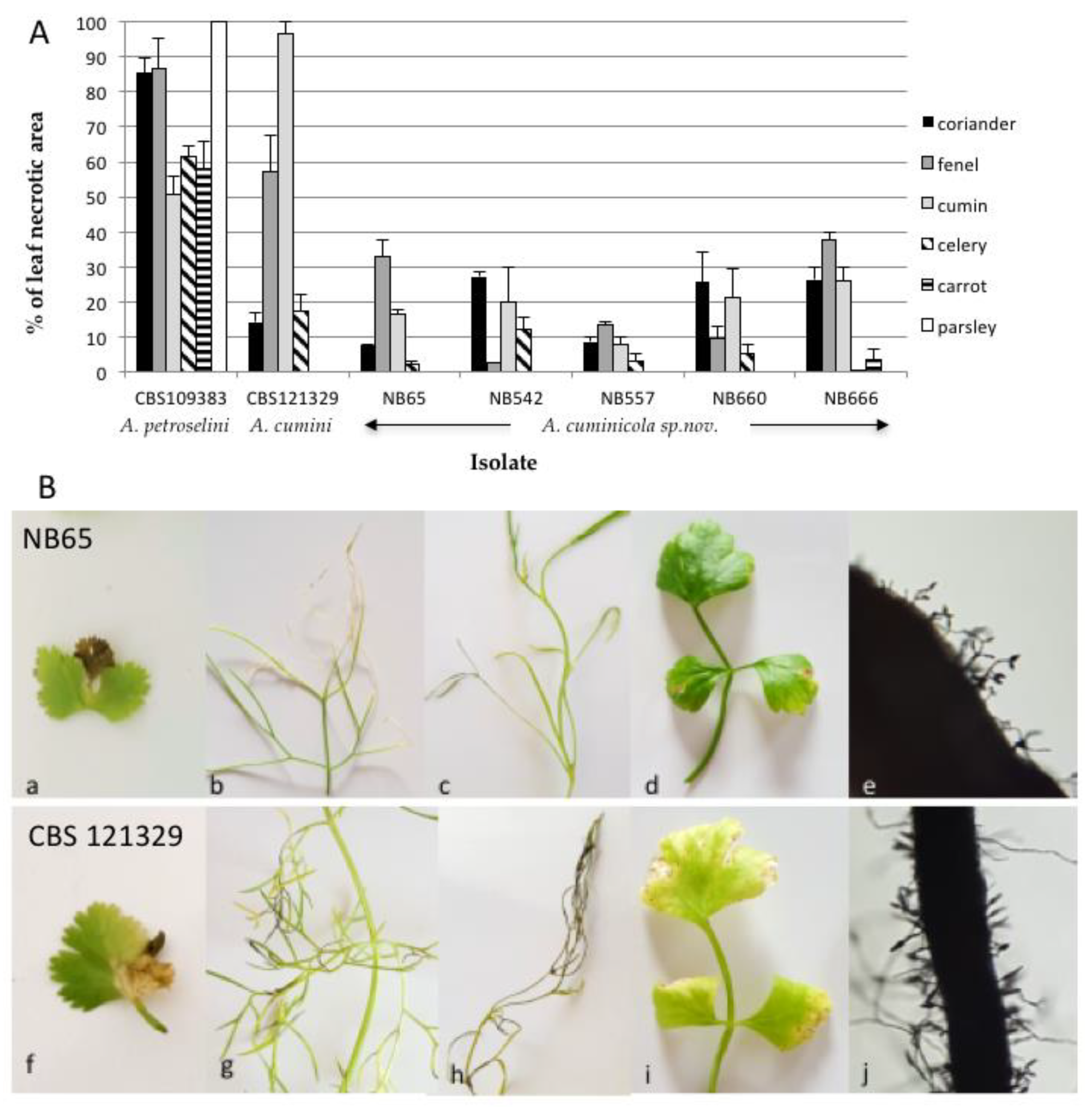
To assess the host range of isolates from section Eureka, six species from Apiaceae, i.e., coriander, carrot, parsley, fennel, celery, cumin, were inoculated with five isolates (NB65, NB542, NB557, NB660 and NB666). Strains CBS 109383 (A. petroselini) and CBS 121329 (A. cumini) were used as positive controls. As expected, the most susceptible plant species were parsley with A. petroselini and cumin with A. cumini (Figure 10). The latter species produced symptoms on all tested plant species except for parsley and carrot. Symptoms were also observed on coriander, fennel and cumin plants with the five A. cuminicola sp. nov. isolates. Discrete symptoms were also observed on celery, except with NB666. However, these isolates were only weakly aggressive towards these plants, and non-pathogenic to parsley and carrot at 21 DAI. The inoculated leaves of cumin showed distinct blighted tips on lower leaves at 21 DAI, and sporulation was present on lesions after four weeks (Figure 10B).
4. Discussion
This study identified small-spored Alternaria spp. belonging to four sections other than those already described for isolates from symptomatic tissues of Solanaceae in north Algeria. Previous studies suggested a high diversity within the Alternaria component of the Algerian mycoflora associated with Solanaceae. At least five large-spored species from section Porri, i.e., A. solani, A. linariae, A. protenta, A. grandis and A. crassa, and two small-spored species from section Alternaria have been characterized from symptomatic tomato tissues [14,30,57]. Recently, A. telliensis, a new species from section Japonicae was also isolated from tomato [26]. The importance of the small-spored Alternaria species of section Alternaria in the development of foliar diseases in Solanaceae has been demonstrated in Algeria [14,18]. Small-spored Alternaria species were detected in about half of the sampled leaves with clear early blight and spot symptoms. Microscopic examinations showed that members of section Alternaria were dominant, representing more than 90% of the small-spored isolates, whereas species from other sections occurred at a lower frequency. Similar trends have been reported in the US [58] and Russia [24]. However, based on their morphology, nearly 8% of the small-spored isolates could not be considered to be members of section Alternaria. The present work focused on the diversity of these new small-spored Alternaria isolates that were associated with foliar lesions on Solanaceae in the north-western regions of the country. The combined use of morphological characteristics and phylogenetic analysis at the gpd locus revealed that these isolates could be classified into four different sections, i.e., sections Infectoriae, Ulocladioides, Embellisioides and Eureka. In agreement with other studies, although colony characteristics, such as color, texture, and sporulation patterns provided useful information for the preliminary separation of individuals into sections, their morphological characteristics were insufficient for accurate delineation at the species or even at the section level [21].
Two strains that were isolated from eggplant and black nightshade were assigned to the section Ulocladioides. This section has been introduced by Woudenberg et al. [46], who synonymized the genus Ulocladium with Alternaria and split the members of this genus into three sections based on gpd-rpb2-tef1 multilocus phylogeny. This section is comprised of 21 recognized species [49], although sequence data for the taxonomy-informative loci are not available for all of them. The two isolates that were assigned to this section could not be differentiated on the basis of morphological characteristics. However, morphological characteristics of species formerly included in the genera Ulocladium are subject to variation according to growth conditions and can cause difficulty in establishing new taxa [48]. Members of this section were already described for tomato plants in Algeria and were identified as A. consortialis based on ITS rDNA sequence analysis [14]. This locus has limited value for distinguishing species within the Alternaria genus and this identification is therefore questionable. In fact, it was impossible to discriminate between the two isolates from section Ulocladioides that were under investigation in the present study and a group of phylogenetically related isolates using combined gpd-rpb2-tef1 phylogeny. To better resolve the individuals within this section, Dang et al. [48] proposed the use of a gpd-Alt a1-MAT 1-2-1-AGA1 phylogeny. Despite many attempts, we were unable to amplify at the latter locus. Sequence comparisons at the gpd, Alt a1 and MAT 1-2-1 were performed but they failed to discriminate the two isolates that were under study from two other phylogenetically related species (A. cucurbitae and A. brassicae-pekinensis).
More than half of the small-spored isolates from our strain collection were assigned to section Infectoriae. This section is characterized by the fact it includes species known to produce a sexual state by contrast to other Alternaria sections, along with additional characteristics (e.g., production of unique metabolites). A recent study even recommended the consideration of a taxonomic re-evaluation of the section Infectoriae which, due to the morphology, sexuality, genetic and mycotoxin profile of the species, could be defined as different fungal genus from Alternaria [59]. However, the gpd phylogeny that was used in our study to delineate sections confirmed that the section Infectoriae constitutes a well-defined group that is phylogenetically related to other groups defined within the genus Alternaria [46]. Most species within the section Infectoriae exhibited noticeable morphological similarities that could be resolved using phylogenetic analyses [52]. However, such analyses also revealed the high genetic variability of this section compared to others [59]. Using the combined datasets of ITS rDNA, gpd, rpb2, tef1 and ATPase, new species have recently been defined within the section Infectoriae [51,55]. Using similar loci and comparisons with all of the species for which the corresponding sequences were available in databank, we showed that several isolates included in this section clustered independently with little to no statistical support for their phylogenetic position, which is similar to previous observations [54]. These authors recommended that species names should only be applied to isolates that possess a strongly supported phylogenetic position within the section Infectoriae. Five isolates from section Infectoriae could be clearly distinguished from each other by analyzing the above-mentioned set of sequences. They formed two well-supported groups, one at the upper part of the tree that we propose to include in a newly described species named A. dahrensis sp. nov., and another closely related but statistically different to A. humuli that formed a sister species named A. pseudohumuli sp. nov.
Alternaria species from section Infectoriae were detected as common species in food, the indoor environment and human cutaneous infections [23,51,60,61,62]. Recent studies have reported the incidence of species belonging to this section on potato plants in the Pacific North-west [58], in Pakistan [63], in Iran [17], in Russia [64], and our inoculation assays have indeed confirmed that the species from this section could be pathogenic to this host as well as to tomatoes.
The combined phylogenetic analysis from the gpd, tef1 and rpb2 genes indicated that isolates of the phylogenetically related sections Eureka and Embellisioides were isolated from Solanaceae in Algeria. A species belonging to the latter section (i.e., A. chlamydosporigena) was isolated recently from a tomato leaf spot in Iran [25]. The strain that was isolated from tomato leaves in Algeria (NB354) produced longer conidia, no chlamydospores, and grouped with A. lolli. Although the separation between NB354 and this species was statistically supported, as a single field isolate was available, we identified NB354 as a member of the species A. lolii (Woudenb. & Crous, Syn. Embellisia lolii). This species was described in Lolium perenne [56], a perennial ryegrass which was frequently observed in growing fields with many other weeds and wild Solanaceae such as black nightshade and jimson weed [28,57]. This may suggest that weeds may be an important source of Alternaria inoculum, contributing to the establishment of Alternaria spp. population in Solanaceae crops. Concerning the five members of section Eureka, all formed a well-supported group closely related to A. cumini, which has previously been reported on cumin in India [23] and Japan [65]. These isolates were considered as belonging to a new species based on morphological traits and multi-locus phylogenetic analysis and was circumscribed as Alternaria cuminicola sp. nov. Although all of these strains were isolated from tomato and black nightshade, they did not cause severe symptoms in the inoculated plants. Due to the phylogenetic relationship with A. cumini, the five isolates of Alternaria cuminicola sp. nov. were inoculated onto six Apiaceae species, comparing their host range with other pathogenic species in this family. Our results showed that A. cuminicola isolates were able to infect coriander, fennel and cumin, albeit with low disease severity. This suggests that the preferred host of A. cuminicola probably belongs to another plant family as it was demonstrated recently for A. telliensis, which a species that was initially isolated in Algeria from Solanaceae and was recently found to be responsible for cabbage leaf spot in Iran [66]. Indeed, Alternaria spores are known as dry air spores that are well adapted for wind dispersal [67]. It is thus possible that this species has a broad host range and moves through the air from one crop to another. Areas with large fluctuations in humidity and temperature, and where agricultural activities are prevalent, may have the potential to significantly affect host–pathogen dynamics and epidemic development in natural plant–pathogen communities [68]. Northern Algeria, which is characterized by a Mediterranean climate and where cultural practices (potato and tomato fields in close proximity or even in rotation, solanaceous weed species often present near cultivated areas) may favor host jumps and the emergence of new virulence, and could therefore be considered as a potential hot spot of diversity for the genus Alternaria.
Our data indeed revealed a huge diversity of Alternaria spp. associated with leaf spot symptoms on Solanaceae in Algeria. In addition to the five large-spored species belonging to section Porri as well as members of the two species-complex alternata and arborescens from section Alternaria that have already been described in this country, here we documented the isolates belonging to four other sections and described three new small-spored Alternaria species whose role in the development of leaf spot disease has to be clarified. Indeed, small-spored Altenaria species are often considered as secondary plant pathogens that are able to either infect plants, together with large-spored Alternaria species, or to live on necrotic lesions caused by other plant pathogens. However, we previously showed the ability of strains of A. alternata and A. arborescens to provoke tomato and potato leaf blight and spot. These species may also develop synergistic interactions in mixed infections with moderately aggressive isolates of A. linariae [14]. The data reported here strongly suggest that some of the newly identified small-spored Alternaria species such as A. dahraensis sp. nov. and A. pseudohumuli sp. nov. might be able to infect tomato and/or potato plants and provoke significant leaf blight symptoms, at least under greenhouse conditions.
In conclusion, our results highlight the complexity of the etiology of early blight and brown spot of Solanaceae in Algeria and illustrate the necessity to better understand the pathogen population structure before devising a successful strategy of disease management. This might necessitate the characterization of additional phylogenetically informative molecular markers for the better delineation of species within some sections such as the sections Infectoriae and Ulocladioides.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/life11121291/s1, Figure S1: Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd, tef1, rpb2, Alt a1 and MAT1-2-1 of Alternaria isolates from section Ulocladioides for which these sequences are available. Figure S2: Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of ATPase, Act, Alt a1 1 of Alternaria isolates from section Eureka. Figure S3: Pathogenicity testing of Alternaria isolates on Solanum tuberosum var. var. Désirée.
Author Contributions
Conceptualization, N.B. and P.S.; Data curation, P.S and B.H.; Formal analysis, P.S.; Investigation and methodology, N.B. and N.B.-S.; Validation, N.B., B.H., K.M., N.B.-S., and P.S.; Resources, N.B.-S., B.H. and N.B.; Supervision, P.S. and K.M.; Visualization, writing—original draft preparation, writing—review and editing N.B., and P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequences obtained during this study have been deposited in GenBank and their accession numbers are provided. Specimen for newly described species have been deposited in the Westerdijk Fungal Biodiversity Institute (Utrecht, The Netherlands) in compliance with the Nagoya protocol. All isolates described in this study are freely available upon request to the authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tsedaley, B. Review on early blight (Alternaria spp.) of potato disease and its management options. J. Biol. Agric. Health 2014, 4, 191–198. [Google Scholar]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current Status of Early Blight Resistance in Tomato: An Update. Int. J. Mol. Sci. 2017, 18, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamgain, A.; Roychowdhury, R.; Tah, J. Alternaria pathogenicity and its strategic controls. Res. J. Biol. 2013, 1, 1–9. [Google Scholar]
- Zheng, H.H.; Zhao, J.; Wang, T.Y.; Wu, X.H. Characterization of Alternaria species associated with potato foliar diseases in China. Plant. Pathol. 2015, 64, 425–433. [Google Scholar] [CrossRef]
- Grogan, R.G.; Kimble, K.A.; Misaghi, I. A stem canker disease of tomato caused by Alternaria alternata f. sp. lycopersici. Phytopathology 1975, 65, 880–886. [Google Scholar] [CrossRef]
- Droby, S.; Dinoor, A.; Prusky, D.; Barkai-Golan, R. Pathogenicity of Alternaria alternata on potato in Israel. Phytopathology 1984, 74, 537–542. [Google Scholar] [CrossRef]
- Akhtar, K.P.; Saleem, M.Y.; Asghar, M.; Haq, M.A. New report of Alternaria alternata causing leaf blight of tomato in Pakistan. Plant Pathol. 2004, 53, 816. [Google Scholar] [CrossRef]
- Raja, P.; Reddy, A.V.R.; Allam, U.S. First report of Alternaria tenuissima causing leaf spot and fruit rot on eggplant (Solanum melongena) in India. Plant Pathol. 2006, 55, 579. [Google Scholar] [CrossRef]
- Van der Waals, J.E.; Pitsi, B.E.; Marais, C.; Wairuri, C.K. First report of Alternaria alternata causing leaf blight of potatoes in South of Africa. Plant Dis. 2011, 95, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Neergaard, P. Danish Species of Alternaria and Stemphylium: Taxonomy, Parasitism, Economic Significance; Oxford University Press: Oxford, UK, 1945; 560p. [Google Scholar]
- Ellis, M.B.; Gibson, I.A.S. Alternaria Solani No. 45 Set 48; Common Wealth Mycological Institute: Kew, UK, 1975. [Google Scholar]
- Rodrigues, T.T.M.S.; Berbee, M.L.; Simmons, E.G.; Cardoso, C.R.; Reis, A.; Maffia, L.A.; Mizubuti, E.S.G. First report of Alternaria tomatophila and A. grandis causing early blight on tomato and potato in Brazil. New Dis. Rep. 2010, 22, 28. [Google Scholar] [CrossRef] [Green Version]
- Bessadat, N.; Hamon, B.; Henni, J.E.; Simoneau, P. First report of tomato early blight caused by Alternaria grandis in Algeria. Plant. Dis. 2016, 102, 2651. [Google Scholar] [CrossRef] [Green Version]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Bataille-Simoneau, N.; Benichou, S.; Mabrouk, K.; Heni, D.E.; Simoneau, P. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in northwestern Algeria. Eur. J. Plant. Pathol. 2017, 148, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Landschoot, S.M.; De Baets, B.; Höfte, M.; Audenaert, K.; Haesaert, G. Identification of A. arborescens, A. grandis and A. protenta as new members of the European Alternaria population on potato. Fungal Biol. 2017, 121, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Taheri, A.S.; Sharif, N.B.; Zare, R.; Abbasi Moghadam, A. Alternaria interrupta, a new pathogen causing potato early blight in Iran. Rostaniha 2009, 10, 72–73. [Google Scholar]
- Ardestani, S.T.; Sharifnabi, B.; Zare, R.; Moghadam, A.A. New Alternaria species associated with potato leaf spot in various potato growing regions of Iran. Iran. J. Plant Pathol. 2010, 45, 83–86. [Google Scholar]
- Bessadat, N.; Benichou, S.; Mabrouk, K.; Henni, J.E. Aggressiveness and morphological variability of small spore Alternaria sp. isolated from Algeria. J. Exp. Biol. Agric. Res. 2014, 2, 265–278. [Google Scholar]
- Ren, Z.J.; Cao, A.C.; Li, J.; Guo, M.X.; Wang, Q.X.; Yan, D.D.; Ouyang, C.B.; Li, Y. First Report of Alternaria alternata Causing Black Spot on Tomato (Solanum lycopersicum) in Tongzhou, China. Plant Dis. 2017, 101, 1680. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidi, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes. Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and taxonomy of the pleomorphic genus Alternaria. Mycol. Prog. 2016, 15, 1–22. [Google Scholar] [CrossRef]
- Al Ghafri, A.; Maharachchikumbura, S.; Hyde, K.; Al-Saady, N.; Al-Sadi, A. A new section and a new species of Alternaria encountered from Oman. Phytotaxa 2019, 405, 279–289. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria. An identification manual. In CBS Biodiversity Series 6; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007; pp. 456–642. [Google Scholar]
- Kokaeva, L.Y.; Belosokhov, A.F.; Doeva, L.Y.; Skolotneva, E.S.; Elansky, S.N. Distribution of Alternaria species on blighted potato and tomato leaves in Russia. J. Plant Dis. Prot. 2017, 125, 205–212. [Google Scholar] [CrossRef]
- Sadeghi, B.; Mirzaei, S. First report of Alternaria Leaf spot caused by Alternaria chlamydosporigena on tomato in Iran. Plant Dis. 2018, 102, 1175. [Google Scholar] [CrossRef]
- Bessadat, N.; Hamon, B.; Bataillé-Simoneau, N.; Mabrouk, K.; Simoneau, P. Alternaria telliensis sp. nov., a new species isolated from Solanaceae in Algeria. Phytotaxa 2020, 440, 89–100. [Google Scholar] [CrossRef]
- Bessadat, N.; Hamon, B.; Bataillé-Simoneau, N.; Kihal, M.; Simoneau, P. Alternaria foliar diseases of solanaceous crops in Algeria: A multi-species threat? Acta Hortic. 2019, 1257, 63–72. [Google Scholar] [CrossRef]
- Ayad, D.; Leclerc, S.; Hamon, B.; Kedad, A.; Bouznad, Z.; Simoneau, P. First report of early blight caused by Alternaria protenta on potato in Algeria. Plant Dis. 2017, 101, 836. [Google Scholar] [CrossRef] [Green Version]
- Ayad, D.; Aribi, D.; Hamon, B.; Kedad, A.; Simoneau, P.; Bouznad, Z. Distribution of large-spored Alternaria species associated with early blight of potato and tomato in Algeria. Phytopathol. Mediterr. 2019, 58, 139–149. [Google Scholar] [CrossRef]
- Snoussi, S.A. Étude de Base sur la Tomate en Algérie. Programme Régional de Gestion Intégrée des Ravageurs Pour le Proche-Orient. 2009. Available online: http://www.ipm-neareast.com (accessed on 28 April 2017).
- Bessadat, N.; Simoneau, P.; Benichou, S.; Setti, B.; Mabroukl, K.; Henni, J.E. Morphological, physiological and pathogenic variability of small-spore Alternaria causing leaf blight of Solanaceae in Algeria. Afr. J. Microbiol. Res. 2014, 8, 3422–3434. [Google Scholar] [CrossRef] [Green Version]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; pp. 16–225. [Google Scholar]
- Goodwin, D.C.; Lee, S.B. Microwave miniprep of total genomic DNA from fungi, plants, protists and animals for PCR. BioTechniques 1993, 15, 438–444. [Google Scholar] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: Formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal. Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Li, Z.; Xia, L.Y.; Wang, Q.; Hu, X.M.; Zhang, X.G. Characterization and phylogenetic analysis of the mating-type loci in the asexual ascomycete genus Ulocladium. Mycologia 2014, 106, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA 7, Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-Tree: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 532, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3, Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [Green Version]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debeta, A.J.M.; Bilanenko, E.N. On the diversity of fungi from soda soils. Fungal Div. 2016, 76, 27–74. [Google Scholar] [CrossRef] [Green Version]
- Dang, J.L.; Gleason, M.L.; Li, L.N.; Wang, C.; Niu, C.K.; Zhang, R.; Sun, G.Y. Alternaria malicola sp. nov., a New Pathogen Causing Fruit Spot on Apple in China. Plant Dis. 2018, 102, 1273–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannibal, P.B.; Lawrence, D.P. Distribution of Alternaria species among sections. 6. Species formerly assigned to genus Ulocladium. Mycotaxon 2018, 133, 293–299. [Google Scholar] [CrossRef]
- Pryor, B.M.; Michailides, T.J. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria late blight of pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iturrieta-Gonzalez, I.; Pujol, I.; Iftimie, S.; Garcia, D.; Morente, V.; Queralt, R.; Guevara-Suarez, M.; Alastruey-Izquierdo, A.; Ballester, F.; Hernandez-Restrepo, M.; et al. Polyphasic identification of three new species in Alternaria section Infectoriae causing human cutaneous infection. Mycoses 2020, 63, 212–224. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Dugan, F.M.; Pryor, B.M. Characterization of Alternaria isolates from the infectoria species-group and a new taxon from Arrhenatherum, Pseudoalternaria arrhenatheria sp. nov. Mycol. Prog. 2014, 13, 257–276. [Google Scholar] [CrossRef]
- Gannibal, P.B.; Lawrence, D.P. Distribution of Alternaria species among sections. 3. Sections Infectoriae and Pseudoalternaria. Mycotaxon 2016, 131, 781–790. [Google Scholar] [CrossRef]
- Poursafar, A.; Ghosta, Y.; Orina, A.S.; Gannibal, P.B.; Javan-Nikkhah, M.; Lawrence, D.P. Taxonomic study of Alternaria sections Infectoriae and Pseudoalternaria associated with black (sooty) head mold of wheat and barley in Iran. Mycol. Prog. 2018, 17, 343–356. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; Garcia, D.; Gene, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef] [PubMed]
- Simmons, E.G. Novel dematiaceous hyphomycetes. Stud. Mycol. 2004, 50, 109–118. [Google Scholar]
- Bessadat, N.; Hamon, B.; Bataillé-Simoneau, N.; Château, C.; Mabrouk, K.; Simoneau, P. Occurrence of leaf spot disease caused by Alternaria crassa (Sacc.) Rands on Jimson weed and potential additional host plants in Algeria. Plant. Pathol. J. 2020, 36, 179–184. [Google Scholar] [CrossRef]
- Tymon, L.S.; Peever, T.L.; Johnson, A.D. Identification and enumeration of small-spored Alternaria associated with potato in the U.S. Northwest. Plant Dis. 2016, 100, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somma, S.; Amatulli, M.T.; Masiello, M.; Moretti, A.; Logrieco, A.F. Alternaria species associated to wheat black point identified through a multilocus sequence approach. Int. J. Food Microbiol. 2019, 293, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Castañares, E.; Cabral, L.C.; Dinolfo, M.I.; Andersen, B.; Stenglein, S.A.; Patriarca, A. Alternaria in malting barley: Characterization and distribution in relation with climatic conditions and barley cultivars. Int. J. Food Microbiol. 2021, 357, 109367. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, A.; Fernández Pinto, V. Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–8. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; van der Merwe, N.A.; Jurjević, Ž.; Groenewald, J.Z.; Crous, P.W. Diversity and movement of indoor Alternaria alternata across the mainland USA. Fungal Genet. Biol. 2015, 81, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, U.; Mushtaq, S.; Akhtar, N. First report of Alternaria metachromatica from Pakistan causing leaf spot of tomato. Pak. J. Agric. Sci. 2014, 51, 305–308. [Google Scholar]
- Belosokhov, A.F.; Belov, G.L.; Chudinova, E.M.; Kokaeva, L.Y.; Elansky, S.N. Alternaria spp. and Colletotrichum coccodes in potato leaves with early blight symptoms. Sixt. Euroblight Workshop Spec. Rep. 2017, 18, 181–190. [Google Scholar]
- Nishikawa, J.; Nakashima, C. Japanese species of Alternaria and their species boundaries based on host range. Fungal Syst. Evol. 2020, 5, 197–281. [Google Scholar] [CrossRef] [PubMed]
- Poursafar, A.; Youbert, G.; Razmig, A.B. Alternaria telliensis, a new causal agent of cabbage leaf spot in Iran. Mycol. Iran. 2020, 7, 231–239. [Google Scholar] [CrossRef]
- Chou, H.H.; Wu, W.S. Phylogenetic analysis of internal transcribed spacer regions of the genus Alternaria, and the significance of filament-beaked conidia. Mycol. Res. 2002, 106, 164–169. [Google Scholar] [CrossRef]
- Burdon, J.J.; Thrall, P.H. The demography and genetics of host–pathogen interactions. In Integrating Ecology and Evolution in a Spatial Context; Silvertown, J., Antonovics, J., Eds.; Blackwell Science: Oxford, UK, 2001; pp. 197–217. [Google Scholar]
Figure 1.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd sequences of 20 small-Scheme 75% are indicated by arrows. The GenBank acc. no. of sequences of the reference strains were from [46] except for members of the section Soda from [47] and section Omanenses from [22].
Figure 1.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd sequences of 20 small-Scheme 75% are indicated by arrows. The GenBank acc. no. of sequences of the reference strains were from [46] except for members of the section Soda from [47] and section Omanenses from [22].

Figure 2.
Conidia and conidiophores of Alternaria sp. sect. Ulocladioides NB559 (a), NB558 (b), Alternaria sp. sect. Infectoriae: NB545 (c), NB540 (d), NB569 (e), NB543 (f), NB561 (g), Alternaria sp. sect. Embellisioides NB354 (h). Bars = 50 µm.
Figure 2.
Conidia and conidiophores of Alternaria sp. sect. Ulocladioides NB559 (a), NB558 (b), Alternaria sp. sect. Infectoriae: NB545 (c), NB540 (d), NB569 (e), NB543 (f), NB561 (g), Alternaria sp. sect. Embellisioides NB354 (h). Bars = 50 µm.

Figure 3.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd, Alt a1 and MAT1-2-1 of Alternaria isolates from section Ulocladioides. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [48].
Figure 3.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd, Alt a1 and MAT1-2-1 of Alternaria isolates from section Ulocladioides. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [48].
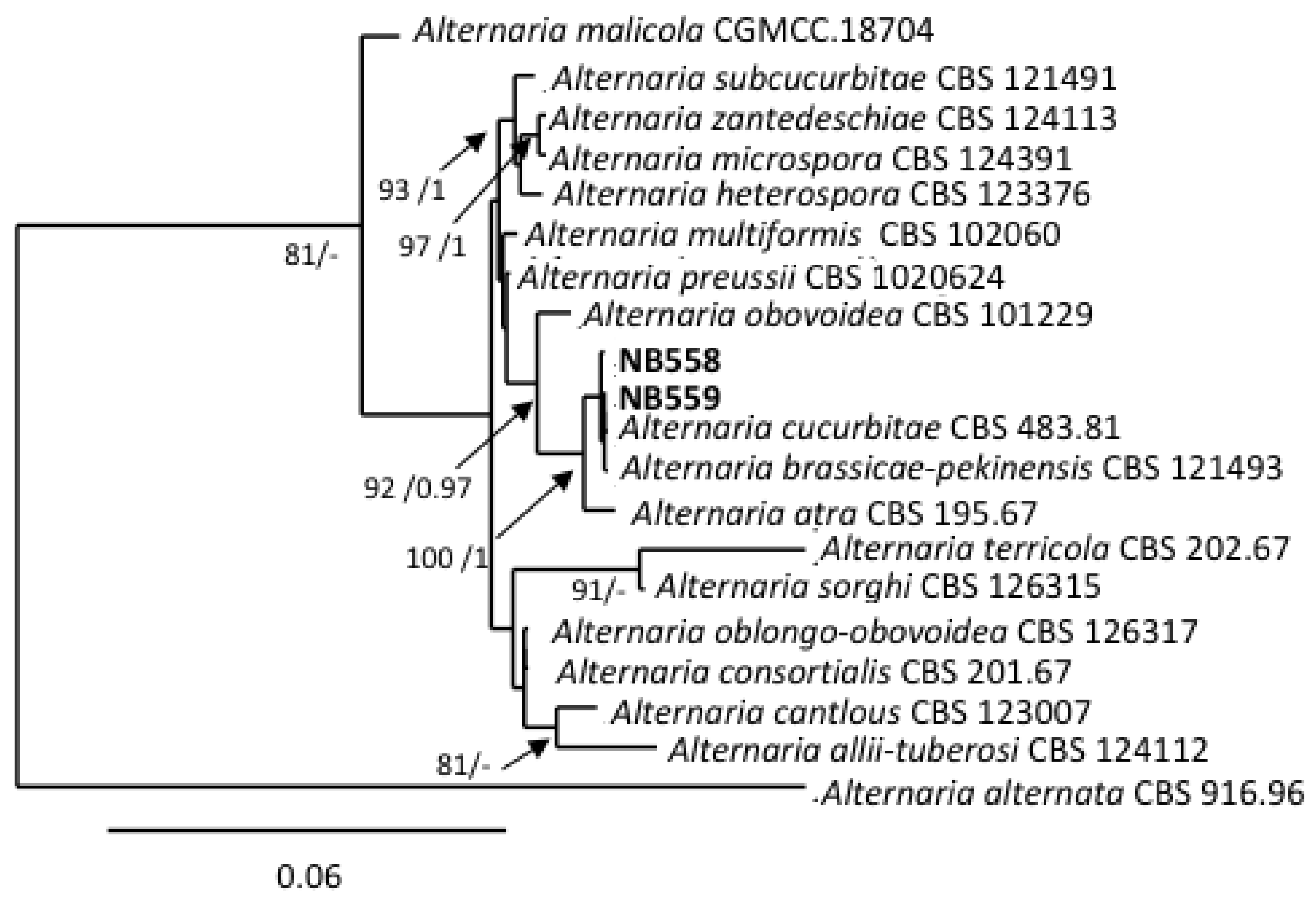
Figure 4.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of ITS rDNA, gpd, rpb2, tef1 and ATPase of Alternaria isolates from section Infectoriae. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [51,55].
Figure 4.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of ITS rDNA, gpd, rpb2, tef1 and ATPase of Alternaria isolates from section Infectoriae. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [51,55].
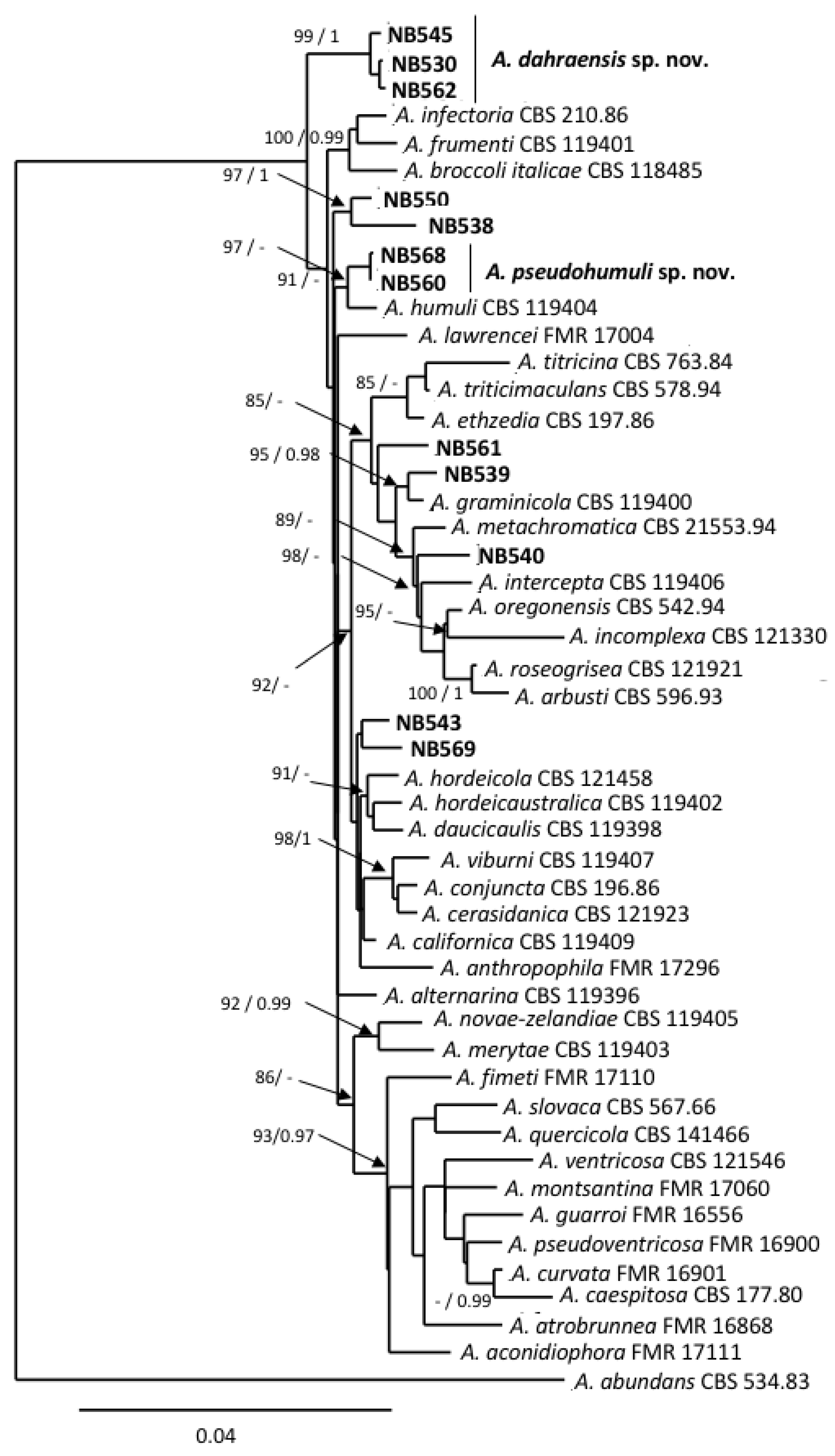
Figure 5.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd, rpb2, tef1 of Alternaria isolates from sections Embellisioides and Eureka. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [46].
Figure 5.
Phylogenetic tree reconstructed by the maximum likelihood method from the alignment of gpd, rpb2, tef1 of Alternaria isolates from sections Embellisioides and Eureka. Bootstrap support values greater than 75% and Bayesian posterior probabilities greater than 0.95 are indicated near nodes. The GenBank acc. no. of sequences of the strains included as references were retrieved from [46].
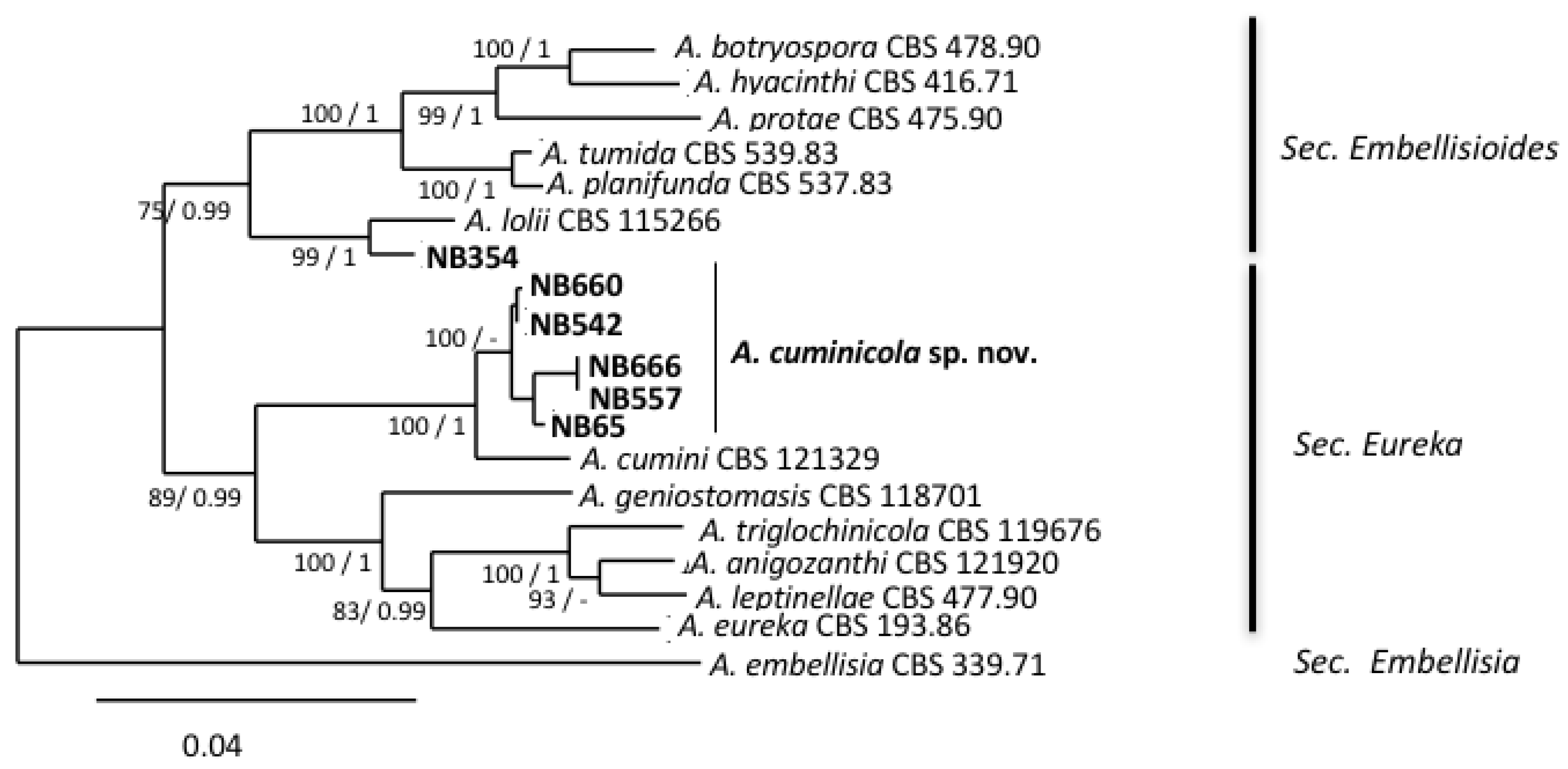
Figure 6.
Alternaria dahraensis sp. nov. (a) Primary mature conidia. (b) Conidia with short and long apical secondary conidiophore (bars = 25 µm). (c) Short conidia chains produced on fertile aerial hyphae (bar = 100 µm). (d) Juvenile conidia with short-beak (right) and tapered apex (left), (bars = 25 µm). (e) Conidia with apical and lateral secondary conidiophores (bar = 25 µm). Colony aspect on: (g) Oatmeal Agar (OA), (h) Malt Extract Agar (MEA), (i) Potato carrot Agar (PCA), (j) Potato Dextrose Agar (PDA) after 7 days of incubation at 25 °C (bar = 10 mm). (k) Conidia and conidiophores produced in the central area of PCA colony (bar = 50 µm), (l) Conidiophores and conidia produced on periphery of PCA colony at 7–14 days (bars = 50 µm). Sporulation pattern on PCA at: (m) 14 and (f) 21 days (bar = 50 µm).
Figure 6.
Alternaria dahraensis sp. nov. (a) Primary mature conidia. (b) Conidia with short and long apical secondary conidiophore (bars = 25 µm). (c) Short conidia chains produced on fertile aerial hyphae (bar = 100 µm). (d) Juvenile conidia with short-beak (right) and tapered apex (left), (bars = 25 µm). (e) Conidia with apical and lateral secondary conidiophores (bar = 25 µm). Colony aspect on: (g) Oatmeal Agar (OA), (h) Malt Extract Agar (MEA), (i) Potato carrot Agar (PCA), (j) Potato Dextrose Agar (PDA) after 7 days of incubation at 25 °C (bar = 10 mm). (k) Conidia and conidiophores produced in the central area of PCA colony (bar = 50 µm), (l) Conidiophores and conidia produced on periphery of PCA colony at 7–14 days (bars = 50 µm). Sporulation pattern on PCA at: (m) 14 and (f) 21 days (bar = 50 µm).
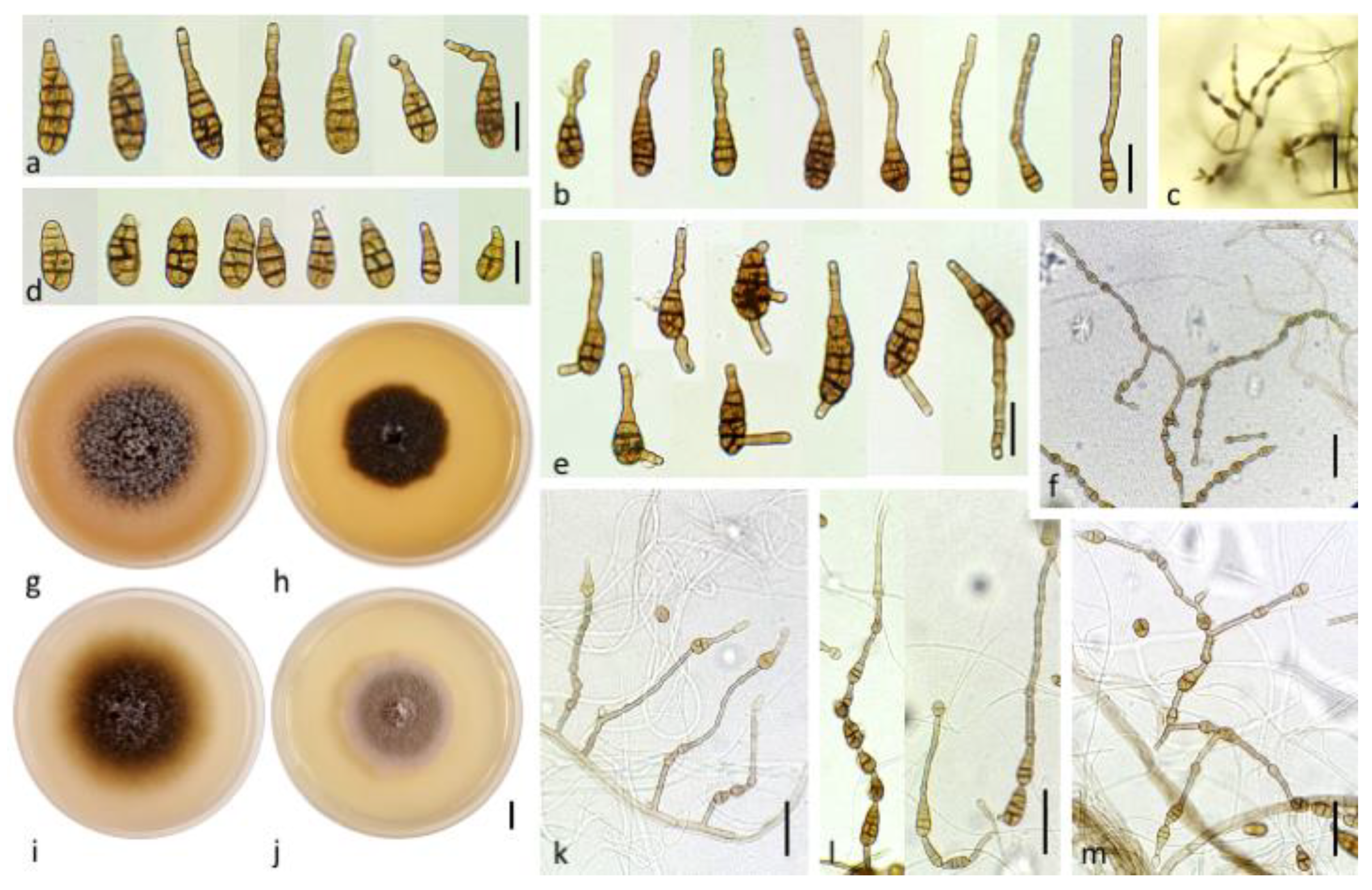
Figure 7.
Alternaria pseudohumuli sp. nov. (a) Mature beakless conidia. Conidia with: (b) short and (e) long apical secondary conidiophore. (c) Juvenile conidia with short-beak (right) and tapered apex (left), (bars = 25 µm). Colony aspect on: (f) Oatmeal Agar (OA), (g) Malt Extract Agar (MEA), (j) Potato carrot Agar (PCA), (k) Potato Dextrose Agar (PDA) after 7 days of incubation at 25 °C (bar = 10 mm). (h) Conidia with lateral secondary conidiophores (bar = 25 µm). Sporulation pattern on water agar 2% at: (l) 7, (i) 14 and (d) 21 days, (bar = 50 µm).
Figure 7.
Alternaria pseudohumuli sp. nov. (a) Mature beakless conidia. Conidia with: (b) short and (e) long apical secondary conidiophore. (c) Juvenile conidia with short-beak (right) and tapered apex (left), (bars = 25 µm). Colony aspect on: (f) Oatmeal Agar (OA), (g) Malt Extract Agar (MEA), (j) Potato carrot Agar (PCA), (k) Potato Dextrose Agar (PDA) after 7 days of incubation at 25 °C (bar = 10 mm). (h) Conidia with lateral secondary conidiophores (bar = 25 µm). Sporulation pattern on water agar 2% at: (l) 7, (i) 14 and (d) 21 days, (bar = 50 µm).

Figure 8.
Alternaria cuminicola sp. nov. (a) Medium and mature conidia, (b) small conidia produced in aerial branches (bars = 10 µm), (c) conidia with one apical secondary conidiophore (bar = 50 µm). Colony aspect on: (d) Oatmeal Agar (OA), (e) Malt Extract Agar (MEA), (f) Potato carrot Agar (PCA), (g) Potato Dextrose Agar (PDA) after 7 days of incubation at 20 °C (bar = 10 mm). (h) Lateral branches of primary conidiophore (bar = 50 µm), (i) conidia with one or two lateral secondary conidiophores (bar = 10 µm), (j) Primary conidiophore with lateral branches and short conidia chains (bar = 100 µm), (k) sporulation pattern on PCA at 14 days (bar = 50 µm).
Figure 8.
Alternaria cuminicola sp. nov. (a) Medium and mature conidia, (b) small conidia produced in aerial branches (bars = 10 µm), (c) conidia with one apical secondary conidiophore (bar = 50 µm). Colony aspect on: (d) Oatmeal Agar (OA), (e) Malt Extract Agar (MEA), (f) Potato carrot Agar (PCA), (g) Potato Dextrose Agar (PDA) after 7 days of incubation at 20 °C (bar = 10 mm). (h) Lateral branches of primary conidiophore (bar = 50 µm), (i) conidia with one or two lateral secondary conidiophores (bar = 10 µm), (j) Primary conidiophore with lateral branches and short conidia chains (bar = 100 µm), (k) sporulation pattern on PCA at 14 days (bar = 50 µm).

Figure 9.
Pathogenicity testing of Alternaria isolates on tomato var. St Pierre. (A) Symptoms at 21 DAI on tomato leaves inoculated with A. dahraensis (NB562), A. pseudohumuli (NB560), A. cuminicola (NB 557), A. lolii (NB 354) and Alternaria sp section Ulocladioides (NB558); (B) Percentage of leaf necrotic area recorded at 21 DAI on plants inoculated with 18 Alternaria isolates plus 3 strains from section Alternaria (NB529, NB553, NB555).
Figure 9.
Pathogenicity testing of Alternaria isolates on tomato var. St Pierre. (A) Symptoms at 21 DAI on tomato leaves inoculated with A. dahraensis (NB562), A. pseudohumuli (NB560), A. cuminicola (NB 557), A. lolii (NB 354) and Alternaria sp section Ulocladioides (NB558); (B) Percentage of leaf necrotic area recorded at 21 DAI on plants inoculated with 18 Alternaria isolates plus 3 strains from section Alternaria (NB529, NB553, NB555).
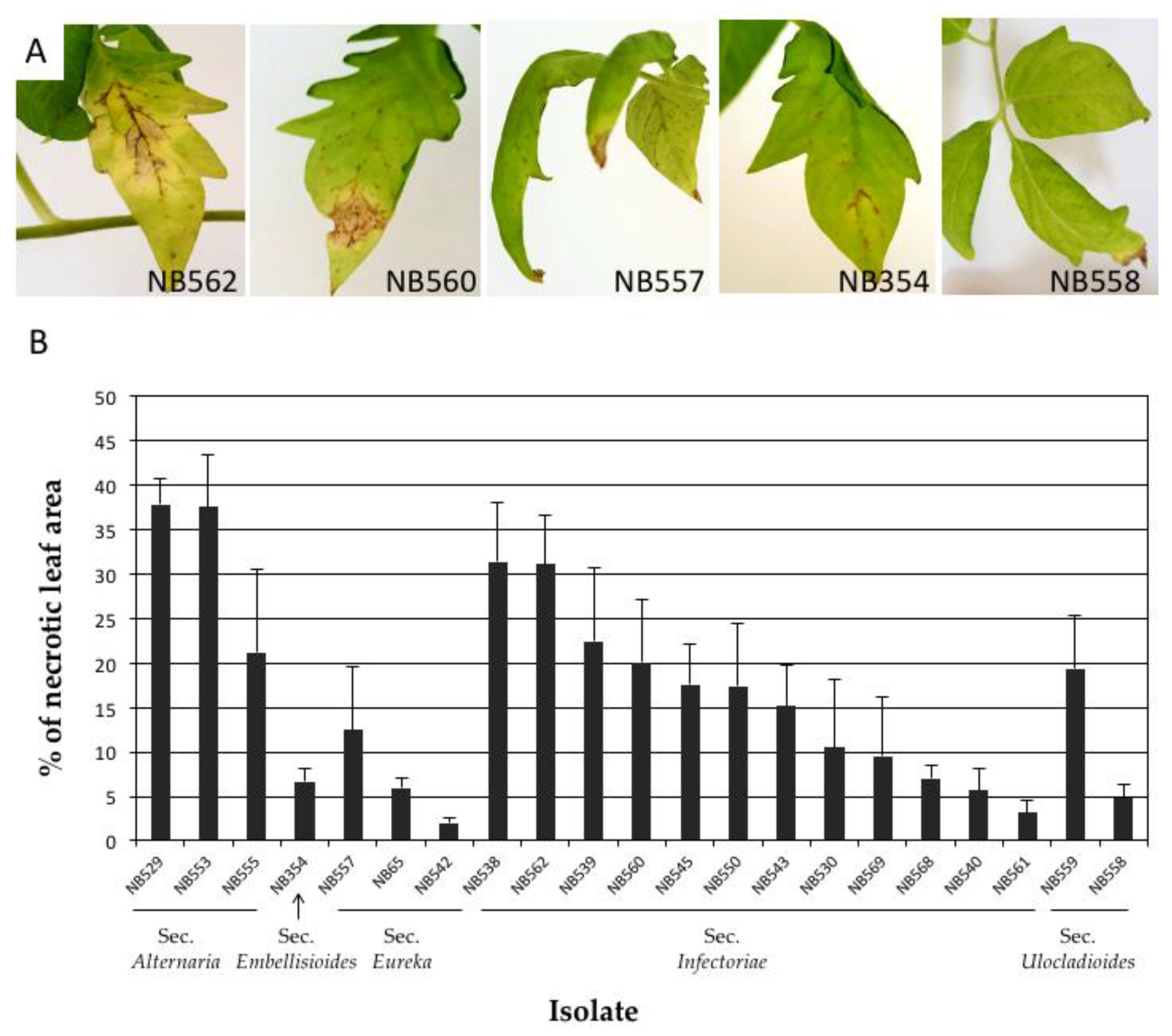
Figure 10.
Pathogenicity testing of Alternaria cuminicola sp. nov. on Apiaceae plant species. (A) Percentage and mean of leaf necrotic area recorded at 21 DAI on Apiaceae leaves inoculated by isolates NB65, NB542, NB557, NB660 and NB666. The Alternaria cumini type strain CBS 121329 and Alternaria petroselini type strain CBS 109383 were included for comparison. (B) Symptoms of Alternaria cuminicola sp. nov. on Apiaceae at 21DAI. A Isolate NB65 on (a) coriander, (b) fennel, (c) cumin and (d) celery, (e) Spores of NB65 produced on cumin leaflet at 28 DAI. Isolate CBS 121329 (A. cumini) on (f) coriander, (g) fennel, (h) cumin and (i) celery, (j) Spores of CBS 121329 produced on cumin leaflet at 28 DAI.
Figure 10.
Pathogenicity testing of Alternaria cuminicola sp. nov. on Apiaceae plant species. (A) Percentage and mean of leaf necrotic area recorded at 21 DAI on Apiaceae leaves inoculated by isolates NB65, NB542, NB557, NB660 and NB666. The Alternaria cumini type strain CBS 121329 and Alternaria petroselini type strain CBS 109383 were included for comparison. (B) Symptoms of Alternaria cuminicola sp. nov. on Apiaceae at 21DAI. A Isolate NB65 on (a) coriander, (b) fennel, (c) cumin and (d) celery, (e) Spores of NB65 produced on cumin leaflet at 28 DAI. Isolate CBS 121329 (A. cumini) on (f) coriander, (g) fennel, (h) cumin and (i) celery, (j) Spores of CBS 121329 produced on cumin leaflet at 28 DAI.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of Alternaria spp. isolates selected for this study.
| Isolate | Host | Symptoms b | Section | GenBank Accession # | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ITS | gpd | ATPase | rpb2 | tef1 | Alt a1 | Mat | ||||
| NB65 | Tomato | BS | Eureka | OK353786 | MK904515 | MK913533 | MK904529 | MK904538 | MK940316 | / |
| NB354 a | Tomato | BS | Embellisioides | OK353787 | MK904520 | / | MK904534 | MK904543 | / | / |
| NB530a | Eggplant | BS | Infectoriae | OK353801 | MK904502 | MK913517 | OK358873 | OK358885 | / | / |
| NB538 a | Pepper | BS | Infectoriae | OK353802 | MK904503 | MK913518 | OK358874 | OK358886 | / | / |
| NB539 a | Pepper | BS | Infectoriae | OK353803 | MK904509 | MK913524 | OK358875 | OK358887 | / | / |
| NB540 a | Eggplant | BS | Infectoriae | OK353804 | MK904510 | MK913525 | OK358876 | OK358888 | / | / |
| NB542a | Black nightshade | BS | Eureka | OK353788 | MK904516 | OK358909 | MK904530 | MK904539 | OK358902 | / |
| NB543 a | Eggplant | EB/BS | Infectoriae | OK353805 | MK904513 | MK913528 | OK358877 | OK358889 | / | / |
| NB545a | Potato | EB/BS | Infectoriae | OK353806 | MK904504 | MK913519 | OK358878 | OK358890 | / | / |
| NB550 a | Pepper | EB/BS | Infectoriae | OK353807 | MK904505 | MK913520 | OK358879 | OK358891 | / | / |
| NB557a | Tomato | BS | Eureka | OK353789 | MK904517 | OK358910 | MK904531 | MK904540 | OK358903 | / |
| NB558 a | Black nightshade | BS | Ulocladioides | OK353790 | MH232169 | / | MK904526 | MK904545 | MN473190 | OK358907 |
| NB559 a | Eggplant | BS | Ulocladioides | OK353791 | MH287762 | / | MK904527 | MK904546 | MN473191 | OK358908 |
| NB560a | Eggplant | BS | Infectoriae | OK353808 | MK904507 | MK913522 | OK358880 | OK358892 | / | / |
| NB561 a | Potato | EB/BS | Infectoriae | OK353809 | MK904512 | MK913527 | OK358881 | OK358893 | / | / |
| NB562a | Tomato | EB/BS | Infectoriae | OK353810 | MK904506 | MK913521 | OK358882 | OK358894 | / | / |
| NB568a | Eggplant | BS | Infectoriae | OK353811 | MK904508 | MK913523 | OK358883 | OK358895 | / | / |
| NB569 a | Eggplant | EB/BS | Infectoriae | OK353812 | MK904514 | MK913529 | OK358884 | OK358896 | / | / |
| NB660 | Tomato | BS | Eureka | OK353792 | MK904518 | OK358911 | MK904532 | MK904541 | OK358904 | / |
| NB666 | Tomato | BS | Eureka | OK353793 | MK904519 | OK358912 | MK904533 | MK904542 | OK358905 | / |
a Isolates selected for morphological characterizations; b BS = black spot, EB = early blight. Isolates identified as new species are indicated in bold characters. # Stands for Numbers.
Table 2.
Cultural characteristics and temperature effect on new species growth after 7 days of incubation on PCA, PDA, MEA and OA.
Table 2.
Cultural characteristics and temperature effect on new species growth after 7 days of incubation on PCA, PDA, MEA and OA.
| Colony Diameter (mm) at | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | Medium | Colony Type | Colony Color | Sporulation at 25 °C | 4 °C | 25 °C | 30 °C | 35 °C | 40 °C |
| A. cuminicola | PCA | Velvety, appressed | Greyish green (30E6) | +++ | 5.4 ± 0.5 | 68.9 ± 1.7 | 54.3 ± 2.2 | 9.8 ± 1.0 | - |
| PDA | Cottony, dense | Greyish green (27E3) | +++ | 8.3 ± 0.3 | 65.6 ± 1.9 | 47.5 ± 1.1 | 7.4 ± 0.8 | - | |
| OA | Velvety, fluffy | Dull green (30D4) | ++ | 7.1 ± 0.3 | 64.6 ± 0.9 | 42.1 ± 0.9 | 5.5 ± 0.4 | - | |
| MEA | Cottony, dense | greyish green (28ED5) | ++ | 8.3 ± 0.5 | 55.0 ± 1.4 | 44.4 ± 1.1 | 6.4 ± 0.6 | - | |
| A. dahraensis | PCA | Arachnoid to loosely woolly | Olive brown (4E4) | +++ | 8.7 ± 1.7 | 76.1 ± 1.2 | 51.5 ± 1.0 | 9.5 ± 0.6 | - |
| PDA | Cottony compact | Olive (3D3) to orange grey (5B2) | ++ | 6.5 ± 3.7 | 53.2 ± 2.2 | 51.8 ± 1.5 | 10.0 ± 0.4 | 5.3 ± 0.3 | |
| OA | Cottony. fluffy | Olive (3E4) to greyish yellow (3C2) | ++ | 8.1 ± 0.6 | 73.5 ± 1.0 | 60.3 ± 2.9 | 8.9 ± 1.0 | 5.4 ± 0.5 | |
| MEA | Velvety | Olive (3E5)/(3E6) | +++ | 8.6 ± 0.8 | 50.0 ± 1.6 | 48.5 ± 0.6 | 9.8 ± 0.6 | 5.1 ± 0.3 | |
| A. pseudohumuli | PCA | Loosely woolly | Yellowish grey (4B2) | ++ | 10.6 ± 0.5 | 79.4 ± 0.5 | 71.0 ± 0.8 | 13.0 ±1.8 | - |
| PDA | Cottony | Olive brown (4D3) center | + | 10.5 ± 0.6 | 78.5 ± 0.7 | 54.6 ± 0.6 | 19.3 ± 1.0 | - | |
| OA | Cottony. woolly | Greyish beige (4C2) center | + | 11.8 ± 1.0 | 66.8 ± 0.4 | 74.0 ± 1.4 | 17.3 ± 1.0 | 5.3 ± 0.3 | |
| MEA | Cottony dense | Olive brown (4D4) center | + | 10.3 ± 1.0 | 67.6 ± 0.5 | 64.0 ± 0.8 | 13.1 ± 1.3 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bessadat, N.; Hamon, B.; Bataillé-Simoneau, N.; Mabrouk, K.; Simoneau, P. Characterization of New Small-Spored Alternaria Species Isolated from Solanaceae in Algeria. Life 2021, 11, 1291. https://doi.org/10.3390/life11121291
AMA Style
Bessadat N, Hamon B, Bataillé-Simoneau N, Mabrouk K, Simoneau P. Characterization of New Small-Spored Alternaria Species Isolated from Solanaceae in Algeria. Life. 2021; 11(12):1291. https://doi.org/10.3390/life11121291
Chicago/Turabian StyleBessadat, Nabahat, Bruno Hamon, Nelly Bataillé-Simoneau, Kihal Mabrouk, and Philippe Simoneau. 2021. "Characterization of New Small-Spored Alternaria Species Isolated from Solanaceae in Algeria" Life 11, no. 12: 1291. https://doi.org/10.3390/life11121291
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.