DnaK3 Is Involved in Biogenesis and/or Maintenance of Thylakoid Membrane Protein Complexes in the Cyanobacterium Synechocystis sp. PCC 6803



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. SDS-PAGE and Immunoblot Analysis
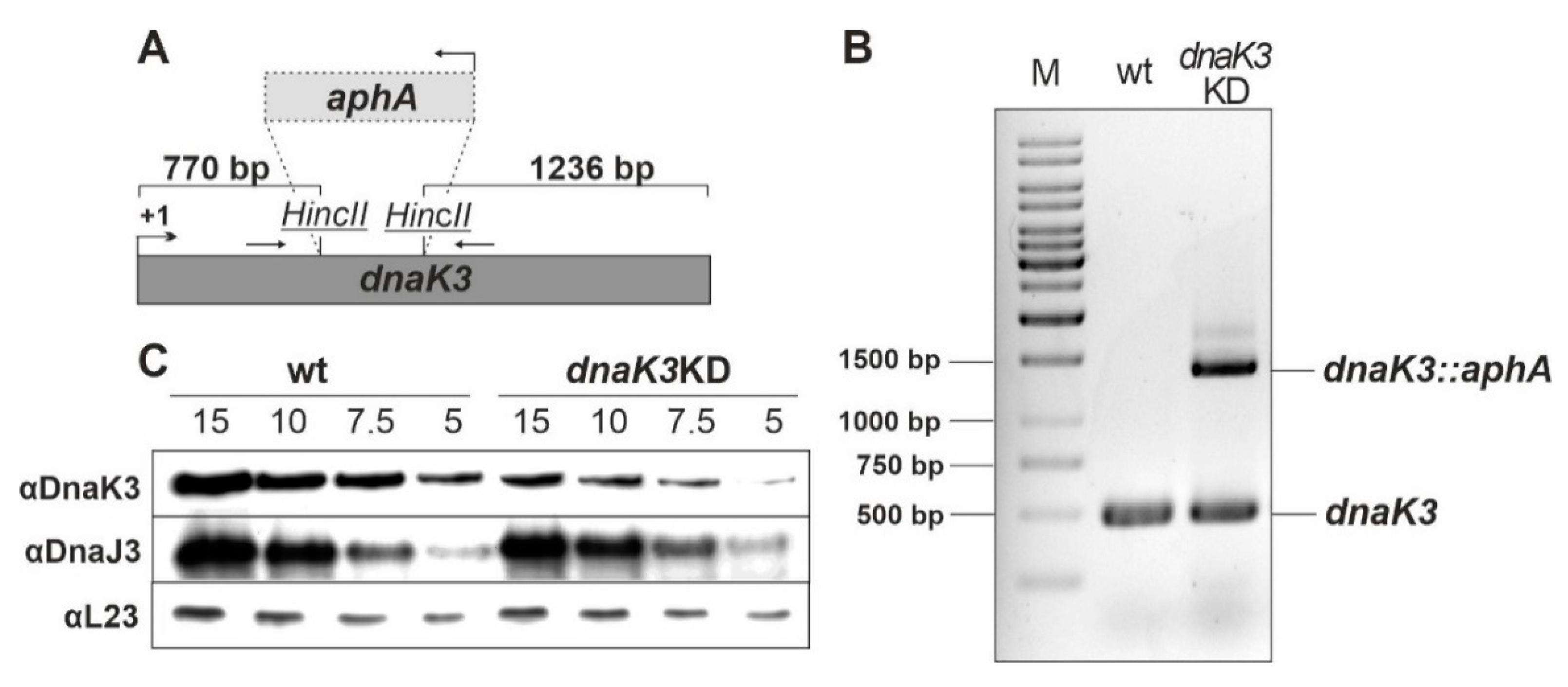
2.3. Complete Deletion of DnaK3 in Synechocystis Cells Grown under LAHG Conditions
2.4. Cell Number and Chlorophyll Concentration Determination
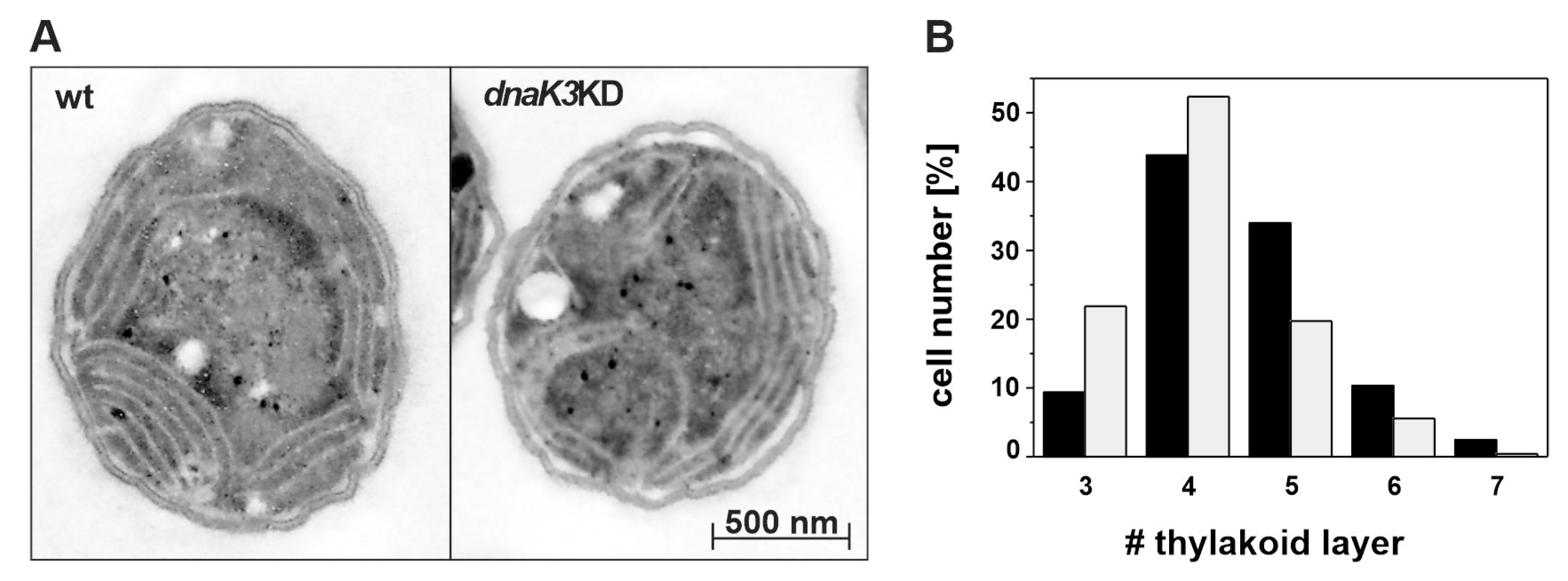
2.5. Electron Microscopy
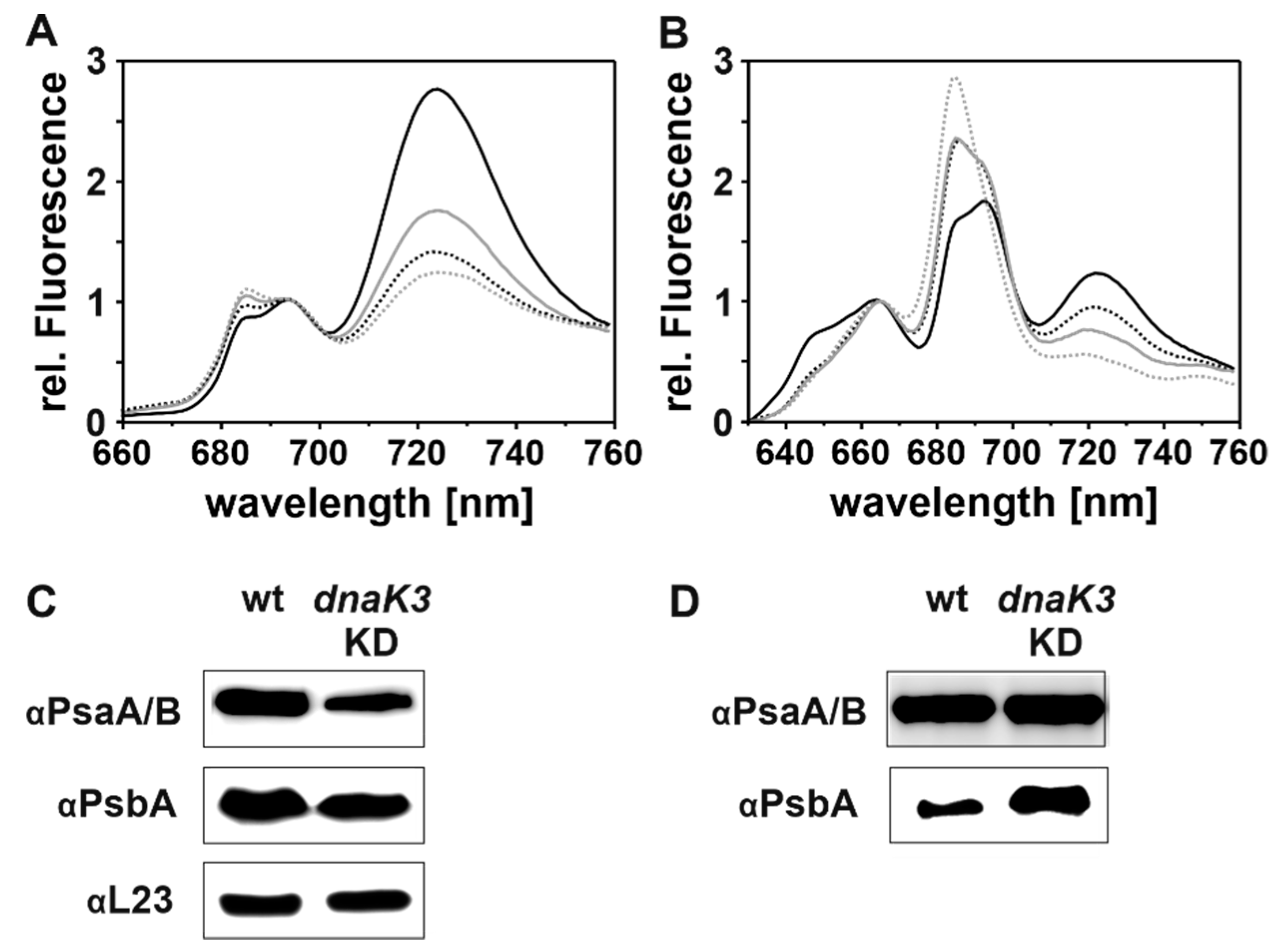
2.6. Absorbance and Low Temperature (77K) Fluorescence Spectra
2.7. Oxygen Evolution
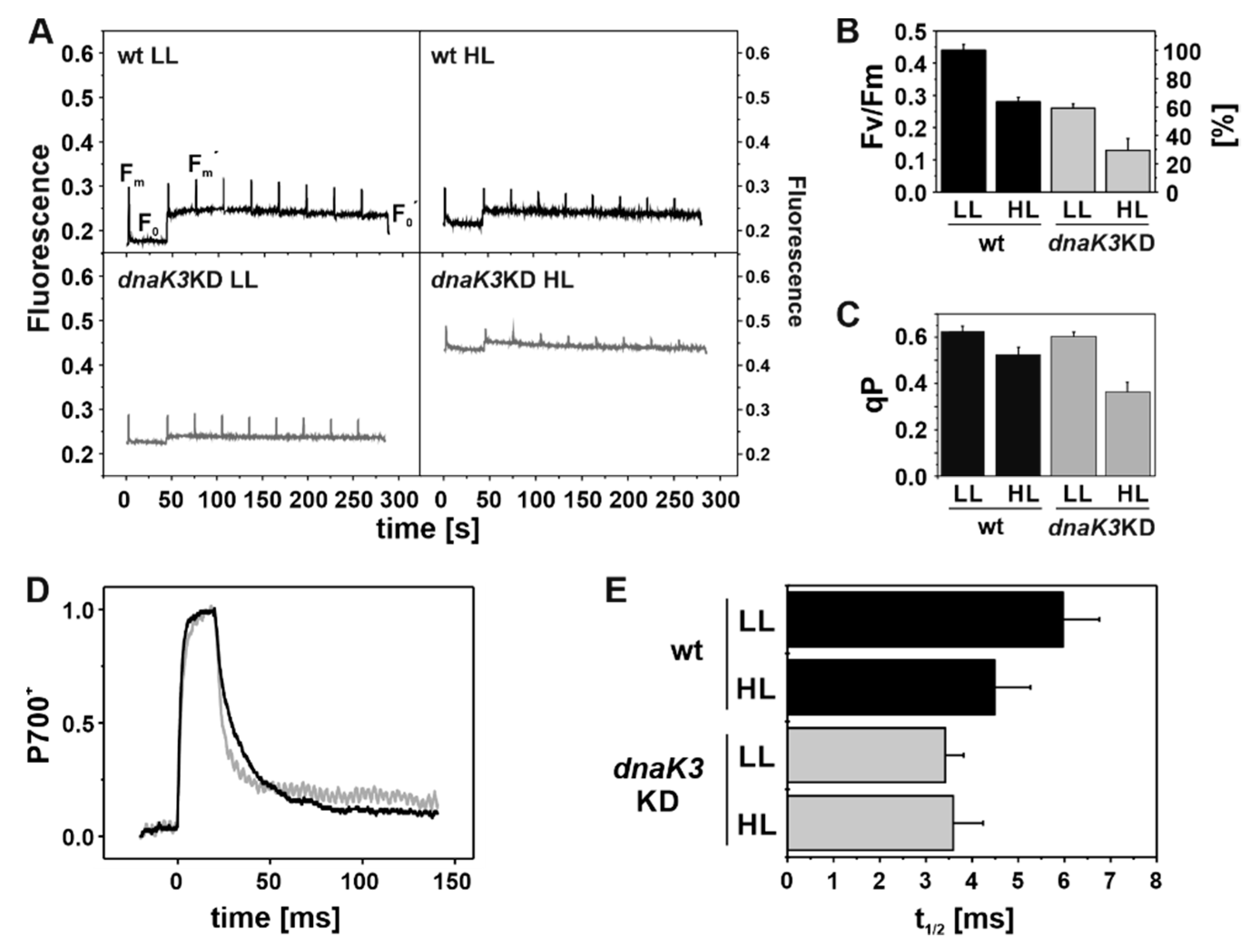
2.8. Chlorophyll Fluorescence Induction Curves
2.9. P700 Re-Reduction Kinetics
3. Results
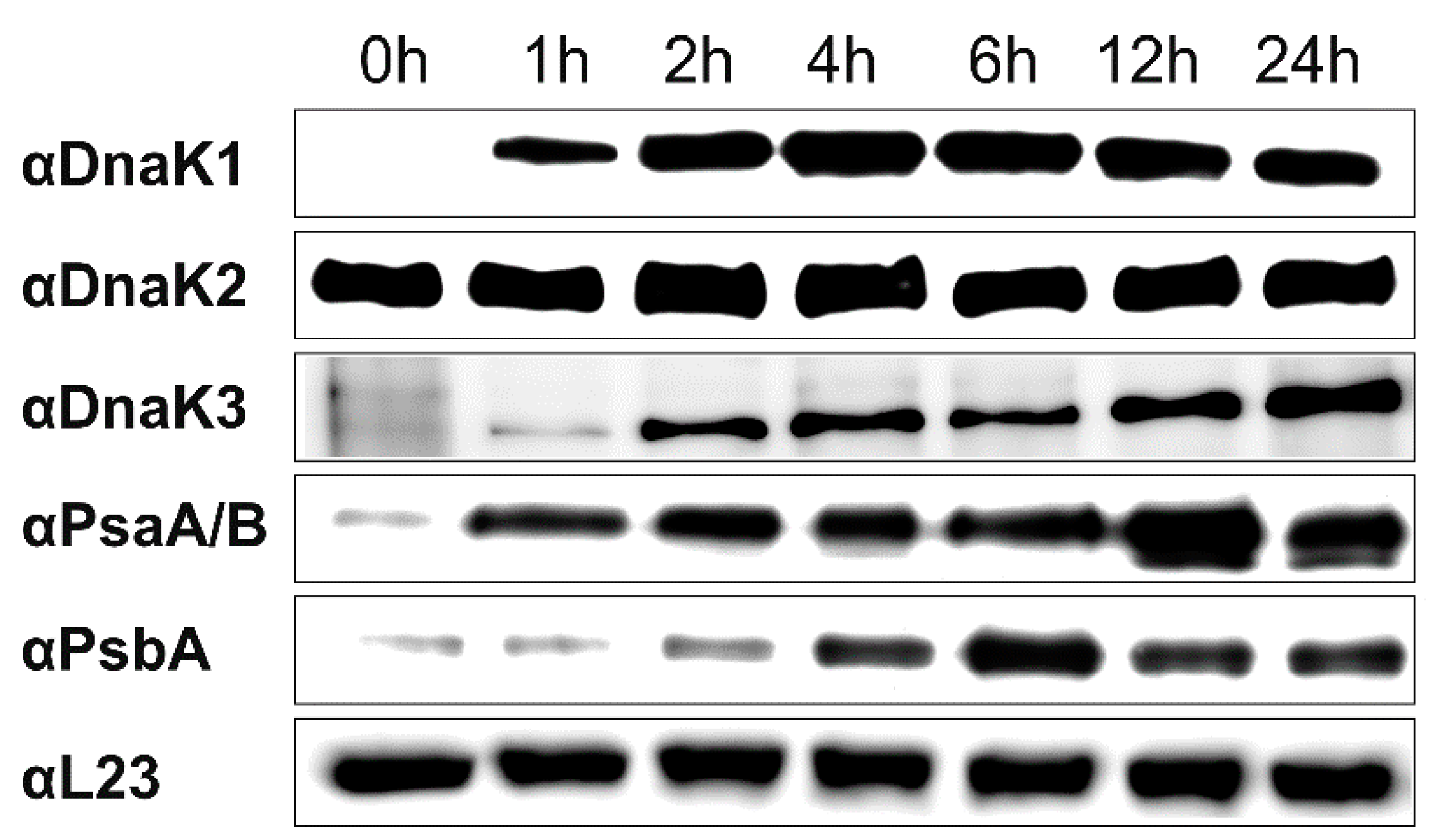
3.1. DnaK3 Synthesis is Light-Induced and Essential in the Dark
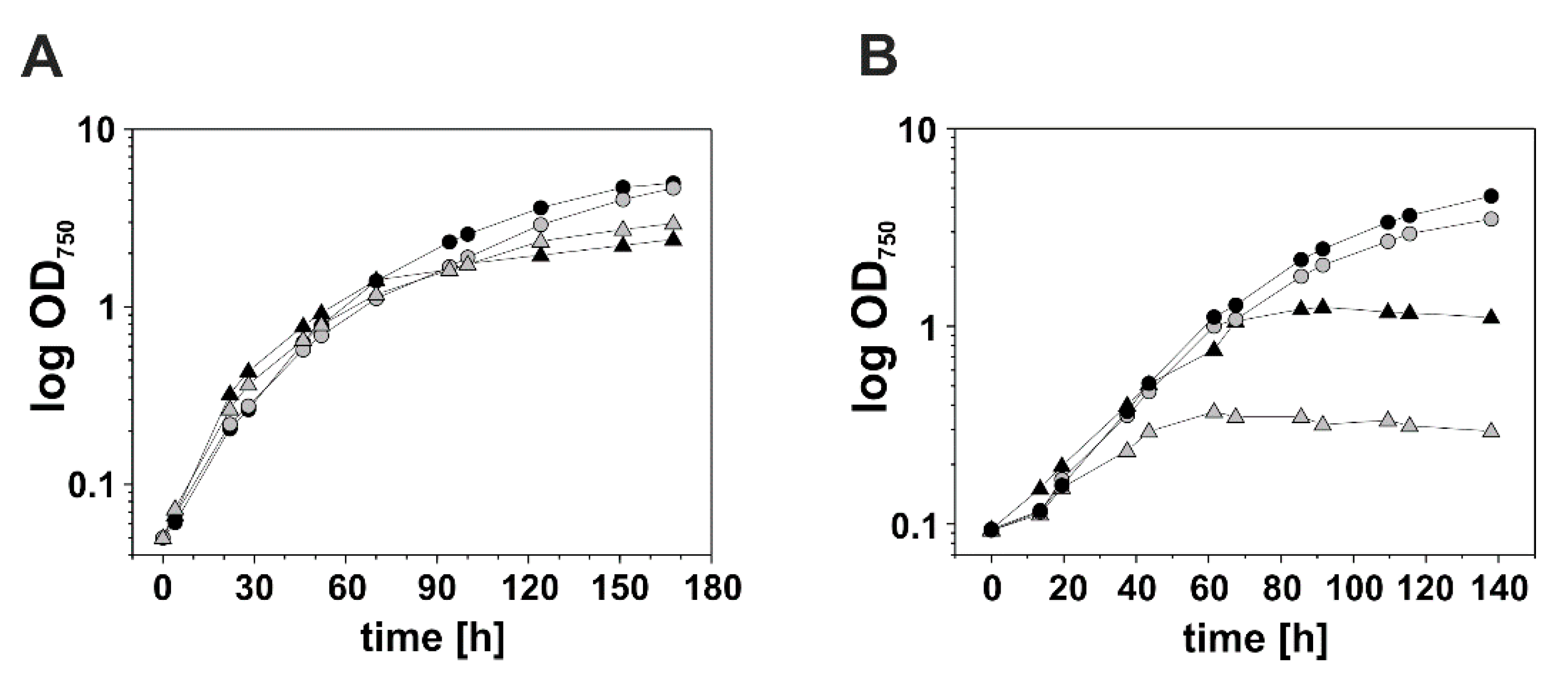
3.2. Reducing the DnaK3 Content Affects Cell Growth under Heat Stress Conditions
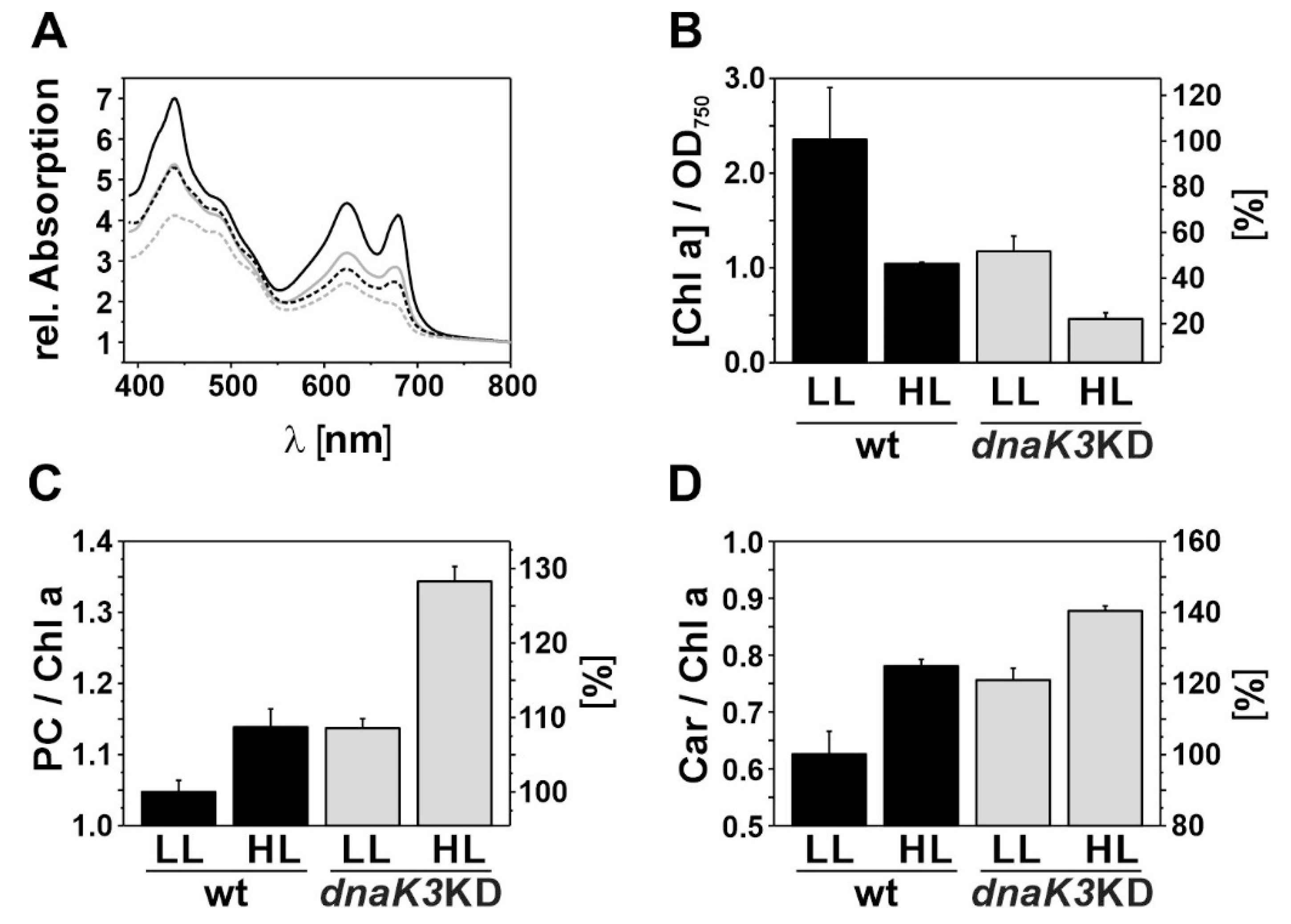
3.3. The dnaK3KD Strain Has a Reduced Pigment Content
3.4. Reducing the DnaK3 Content Results in an Altered PS I-to-PS II Ratio
3.5. The Photosynthetic Activity is Impaired in the DnaK3 Depleted Strain
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rast, A.; Heinz, S.; Nickelsen, J. Biogenesis of thylakoid membranes. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vothknecht, U.C.; Westhoff, P. Biogenesis and origin of thylakoid membranes. Biochim. Biophys. Acta 2001, 1541, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, P.; López-Juez, E. Biogenesis and homeostasis of chloroplasts and other plastids. Nat. Rev. Mol. Cell Biol. 2013, 14, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, C.; Jin, X.; Barajas-López, J.d.D.; Hewitt, T.C.; Tanz, S.K.; Dobrenel, T.; Schröder, W.P.; Hanson, J.; Pesquet, E.; Grönlund, A.; et al. Establishment of Photosynthesis through Chloroplast Development Is Controlled by Two Distinct Regulatory Phases. Plant Physiol. 2018, 176, 1199–1214. [Google Scholar] [CrossRef]
- Vernotte, C.; Picaud, M.; Kirilovsky, D.; Olive, J.; Ajlani, G.; Astier, C. Changes in the Photosynthetic Apparatus in the Cyanobacterium Synechocystis sp Pcc-6714 Following Light-to-Dark and Dark-to-Light Transitions. Photosyn. Res. 1992, 32, 45–57. [Google Scholar] [CrossRef]
- Anderson, S.L.; Mcintosh, L. Light-activated heterotrophic growth of the cyanobacterium Synechocystis sp. strain PCC 6803: A blue-light-requiring process. J. Bacteriol. 1991, 173, 2761–2767. [Google Scholar] [CrossRef] [Green Version]
- Barthel, S.; Bernat, G.; Seidel, T.; Rupprecht, E.; Kahmann, U.; Schneider, D. Thylakoid Membrane Maturation and PSII Activation Are Linked in Greening Synechocystis sp PCC 6803 Cells. Plant Physiol 2013, 163, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Mechela, A.; Schwenkert, S.; Soll, J. A brief history of thylakoid biogenesis. Open Biol. 2019, 9, 180237. [Google Scholar] [CrossRef] [Green Version]
- Spence, E.; Bailey, S.; Nenninger, A.; Moller, S.G.; Robinson, C. A homolog of Albino3/OxaI is essential for thylakoid biogenesis in the cyanobacterium Synechocystis sp PCC6803. J. Biol. Chem. 2004, 279, 55792–55800. [Google Scholar] [CrossRef] [Green Version]
- Thurotte, A.; Brüser, T.; Mascher, T.; Schneider, D.; Brueser, T. Membrane chaperoning by members of the PspA/IM30 protein family. Commun. Integr. Biol. 2017, 10, e1264546. [Google Scholar] [CrossRef]
- Heidrich, J.; Thurotte, A.; Schneider, D. Specific interaction of IM30/Vipp1 with cyanobacterial and chloroplast membranes results in membrane remodeling and eventually in membrane fusion. Biochim. Biophys. Acta Biomembr. 2017, 1859, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Siebenaller, C.; Junglas, B.; Schneider, D. Functional Implications of Multiple IM30 Oligomeric States. Front. Plant Sci. 2019, 10, 1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jilly, R.; Khan, N.Z.; Aronsson, H.; Schneider, D. Dynamin-Like Proteins Are Potentially Involved in Membrane Dynamics within Chloroplasts and Cyanobacteria. Front. Plant Sci. 2018, 9, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, S.J.; Burroughs, N.J.; Evered, C.; Sacharz, J.; Nenninger, A.; Mullineaux, C.W.; Spence, E.M. Loss of the SPHF Homologue Slr1768 Leads to a Catastrophic Failure in the Maintenance of Thylakoid Membranes in Synechocystis sp PCC 6803. PLoS ONE 2011, 6, e19625. [Google Scholar] [CrossRef] [Green Version]
- Rupprecht, E.; Gathmann, S.; Fuhrmann, E.; Schneider, D. Three different DnaK proteins are functionally expressed in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 2007, 153, 1828–1841. [Google Scholar] [CrossRef] [Green Version]
- Katano, Y.; Nimura-Matsune, K.; Yoshikawa, H. Involvement of DnaK3, one of the three DnaK proteins of cyanobacterium Synechococcus sp PCC7942, in translational process on the surface of the thylakoid membrane. Biosci. Biotechnol. Biochem. 2006, 70, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- Frick, G.; Su, Q.X.; Apel, K.; Armstrong, G.A. An Arabidopsis porB porC double mutant lacking light-dependent NADPH: protochlorophyllide oxidoreductases B and C is highly chlorophyll-deficient and developmentally arrested. Plant J. 2003, 35, 141–153. [Google Scholar] [CrossRef]
- Hagio, M.; Sakurai, I.; Sato, S.; Kato, T.; Tabata, S.; Wada, H. Phosphatidylglycerol is essential for the development of thylakoid membranes in Arabidopsis thaliana. Plant Cell Physiol. 2002, 43, 1456–1464. [Google Scholar] [CrossRef] [Green Version]
- Paddock, T.N.; Mason, M.E.; Lima, D.F.; Armstrong, G.A. Arabidopsis protochlorophyllide oxidoreductase A (PORA) restores bulk chlorophyll synthesis and normal development to a porB porC double mutant. Plant Mol. Biol. 2010, 72, 445–457. [Google Scholar] [CrossRef]
- Sato, N.; Hagio, M.; Wada, H.; Tsuzuki, M. Requirement of phosphatidylglycerol for photosynthetic function in thylakoid membranes. Proc. Natl. Acad. Sci. USA 2000, 97, 10655–10660. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genevaux, P.; Georgopoulos, C.; Kelley, W.L. The Hsp70 chaperone machines of Escherichia coli: A paradigm for the repartition of chaperone functions. Mol. Microbiol. 2007, 66, 840–857. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, H.; Chaurasia, A.K.; Apte, S.K. Cyanobacterial heat-shock response: Role and regulation of molecular chaperones. Microbiology 2014, 160, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, E.; Düppre, E.; Schneider, D. Similarities and Singularities of Three DnaK Proteins from the Cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2010, 51, 1210–1218. [Google Scholar] [CrossRef] [Green Version]
- Düppre, E.; Rupprecht, E.; Schneider, D. Specific and promiscuous functions of multiple DnaJ proteins in Synechocystis sp PCC 6803. Microbiology 2011, 157, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Nimura, K.; Takahashi, H.; Yoshikawa, H. Characterization of the dnaK multigene family in the cyanobacterium Synechococcus sp strain PCC7942. J. Bacteriol. 2001, 183, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, H.; Fujita, K.; Ohtaki, A.; Watanabe, S.; Narumi, S.; Maruyama, T.; Suenaga, E.; Misono, T.S.; Kumar, P.K.R.; Goloubinoff, P.; et al. Physical interaction between bacterial heat shock protein (Hsp) 90 and Hsp70 chaperones mediates their cooperative action to refold denatured proteins. J. Biol. Chem. 2014, 289, 6110–6119. [Google Scholar] [CrossRef] [Green Version]
- Oguchi, K.; Nimura, K.; Yoshikawa, H.; Takahashi, H. Sequence and analysis of a dnaJ homologue gene in cyanobacterium Synechococcus sp. PCC7942. Biochem. Biophys. Res. Commun. 1997, 236, 461–466. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Mimuro, M.; Miyashita, H.; Tsuchiya, T.; Sasamoto, S.; Watanabe, A.; Kawashima, K.; Kishida, Y.; et al. Complete genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks thylakoids. DNA Research 2003, 10, 137–145. [Google Scholar] [CrossRef]
- Rippka, R.; Waterbury, J.; Cohenbazire, G. Cyanobacterium Which Lacks Thylakoids. Arch. Microbiol. 1974, 100, 419–436. [Google Scholar] [CrossRef]
- Nimura, K.; Yoshikawa, H.; Takahashi, H. DnaK3, one of the three DnaK proteins of cyanobacterium Synechococcus sp. PCC7942, is quantitatively detected in the thylakoid membrane. Biochem. Biophys. Res. Commun. 1996, 229, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Aro, E.M.; Virgin, I.; Andersson, B. Photoinhibition of Photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Mulo, P.; Sakurai, I.; Aro, E.M. Strategies for psbA gene expression in cyanobacteria, green algae and higher plants: From transcription to PSII repair. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroda, M.; Vallon, O.; Wollman, F.A.; Beck, C.F. A chloroplast-targeted heat shock protein 70 (HSP70) contributes to the photoprotection and repair of photosystem II during and after photoinhibition. Plant Cell 1999, 11, 1165–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokthongwattana, K.; Chrost, B.; Behrman, S.; Casper-Lindley, C.; Melis, A. Photosystem II damage and repair cycle in the green alga Dunaliella salina: Involvement of a chloroplast-localized HSP70. Plant Cell Physiol. 2001, 42, 1389–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, E.; Gathmann, S.; Rupprecht, E.; Golecki, J.; Schneider, D. Thylakoid Membrane Reduction Affects the Photosystem Stoichiometry in the Cyanobacterium Synechocystis sp PCC 6803. Plant Physiol. 2009, 149, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, E.; Bultema, J.B.; Kahmann, U.; Rupprecht, E.; Boekema, E.J.; Schneider, D. The Vesicle-inducing Protein 1 from Synechocystis sp PCC 6803 Organizes into Diverse Higher-Ordered Ring Structures. Mol. Biol. Cell 2009, 20, 4620–4628. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous-Equations for Assaying Chlorophyll-a and Chlorophyll-B Extracted with 4 Different Solvents—Verification of the Concentration of Chlorophyll Standards by Atomic-Absorption Spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- van Kooten, O.; Snel, J.F. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosyn. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Salvucci, M.E.; Crafts-Brandner, S.J. Relationship between the heat tolerance of photosynthesis and the thermal stability of rubisco activase in plants from contrasting thermal environments. Plant Physiol. 2004, 134, 1460–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadulghani, M.; Suzuki, Y.; Nakamoto, H. Light plays a key role in the modulation of heat shock response in the cyanobacterium Synechocystis sp PCC 6803. Biochem. Biophys. Res. Commun. 2003, 306, 872–879. [Google Scholar] [CrossRef]
- Murakami, A.; Fujita, Y. Regulation of Stoichiometry between PsI and PsII in Response to Light Regime for Photosynthesis Observed with Synechocystis Pcc-6714-Relationship between Redox State of Cyt B6-F Complex and Regulation of Psi Formation. Plant Cell Physiol. 1993, 34, 1175–1180. [Google Scholar]
- Muramatsu, M.; Hihara, Y. Transcriptional regulation of genes encoding subunits of photosystem I during acclimation to high-light conditions in Synechocystis sp. PCC 6803. Planta 2003, 216, 446–453. [Google Scholar] [CrossRef]
- Hihara, Y.; Sonoike, K.; Ikeuchi, M. A novel gene, pmgA, specifically regulates photosystem stoichiometry in the cyanobacterium Synechocystis species PCC 6803 in response to high light. Plant Physiol 1998, 117, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Hihara, Y. Acclimation to high-light conditions in cyanobacteria: From gene expression to physiological responses. J. Plant Res. 2012, 125, 11–39. [Google Scholar] [CrossRef]
- Fraser, J.M.; Tulk, S.E.; Jeans, J.A.; Campbell, D.A.; Bibby, T.S.; Cockshutt, A.M. Photophysiological and photosynthetic complex changes during iron starvation in Synechocystis sp. PCC 6803 and Synechococcus elongatus PCC 7942. PLoS ONE 2013, 8, e59861. [Google Scholar] [CrossRef] [Green Version]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 angstrom resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef]
- Guskov, A.; Kern, J.; Gabdulkhakov, A.; Broser, M.; Zouni, A.; Saenger, W. Cyanobacterial photosystem II at 2.9-A resolution and the role of quinones, lipids, channels and chloride. Nat. Struct. Mol. Biol. 2009, 16, 334–342. [Google Scholar] [CrossRef]
- Rakhimberdieva, M.G.; Vavilin, D.V.; Vermaas, W.F.J.; Elanskaya, I.V.; Karapetyan, N.V. Phycobilin/chlorophyll excitation equilibration upon carotenoid-induced non-photochemical fluorescence quenching in phycobilisomes of the Synechocystis sp PCC 6803. Biochim. Biophys. Acta 2007, 1767, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamary, E.; Kiss, V.; Nevo, R.; Adam, Z.; Bernat, G.; Rexroth, S.; Rogner, M.; Reich, Z. Structural and functional alterations of cyanobacterial phycobilisomes induced by high-light stress. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Biggins, J.; Bruce, D. Regulation of Excitation-Energy Transfer in Organisms Containing Phycobilins. Photosyn. Res. 1989, 20, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Liu, X.; Li, Y.; Liu, C.-C.; Yang, F.; Zhao, J.; Sui, S.-F. Structural organization of an intact phycobilisome and its association with photosystem II. Cell Research 2015, 25, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Emlyn-Jones, D.; Ashby, M.K.; Mullineaux, C.W. A gene required for the regulation of photosynthetic light harvesting in the cyanobacterium Synechocystis 6803. Mol. Microbiol. 1999, 33, 1050–1058. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Emlyn-Jones, D. State transitions: An example of acclimation to low-light stress. J. Exp. Bot. 2005, 56, 389–393. [Google Scholar] [CrossRef]
- Kopecna, J.; Komenda, J.; Bucinska, L.; Sobotka, R. Long-term acclimation of the cyanobacterium Synechocystis sp. PCC 6803 to high light is accompanied by an enhanced production of chlorophyll that is preferentially channeled to trimeric photosystem I. Plant Physiol. 2012, 160, 2239–2250. [Google Scholar] [CrossRef] [Green Version]
- Komenda, J.; Tichy, M.; Prasil, O.; Knoppova, J.; Kuvikova, S.; de Vries, R.; Nixon, P.J. The exposed N-terminal tail of the D1 subunit is required for rapid D1 degradation during photosystem II repair in Synechocystis sp PCC 6803. Plant Cell 2007, 19, 2839–2854. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Thompson, E.; Bailey, S.; Kruse, O.; Mullineaux, C.W.; Robinson, C.; Mann, N.H.; Nixon, P.J. FtsH is involved in the early stages of repair of photosystem II in Synechocystis sp PCC 6803. Plant Cell 2003, 15, 2152–2164. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Niyogi, K.K.; Truong, T.B. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Curr. Opin. Plant Biol. 2013, 16, 307–314. [Google Scholar] [CrossRef]
- Campbell, D.; Bruce, D.; Carpenter, C.; Gustafsson, P.; Oquist, G. Two forms of the photosystem II D1 protein alter energy dissipation and state transitions in the cyanobacterium Synechococcus sp PCC 7942. Photosyn. Res. 1996, 47, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Santabarbara, S.; Villafiorita Monteleone, F.; Remelli, W.; Rizzo, F.; Menin, B.; Casazza, A.P. Comparative excitation-emission dependence of the FV/FM ratio in model green algae and cyanobacterial strains. Physiol. Plant 2019, 166, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Mikkat, S.; Huang, F.; Huckauf, J.; Marin, K.; Norling, B.; Hagemann, M. Proteome analysis of salt stress response in the cyanobacterium Synechocystis sp. strain PCC 6803. Proteomics 2006, 6, 2733–2745. [Google Scholar] [CrossRef] [PubMed]
- Mary, I.; Tu, C.J.; Grossman, A.; Vaulot, D. Effects of high light on transcripts of stress-associated genes for the cyanobacteria Synechocystis sp PCC 6803 and Prochlorococcus MED4 and MIT9313. Microbiology 2004, 150, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Prieto, M.A.; Futschik, M.E. CyanoEXpress: A web database for exploration and visualisation of the integrated transcriptome of cyanobacterium Synechocystis sp. PCC6803. Bioinformation 2012, 8, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Sonoike, K.; Hihara, Y.; Ikeuchi, M. Physiological significance of the regulation of photosystem stoichiometry upon high light acclimation of Synechocystis sp PCC 6803. Plant Cell Physiol. 2001, 42, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Parmryd, I.; Nilsson, F.; Persson, A.L.; Pakrasi, H.B.; Andersson, B.; Norling, B. Proteomics of Synechocystis sp. strain PCC 6803: Identification of plasma membrane proteins. Mol. Cell Proteomics 2002, 1, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Jansson, C.; Debus, R.J.; Osiewacz, H.D.; Gurevitz, M.; Mcintosh, L. Construction of an Obligate Photoheterotrophic Mutant of the Cyanobacterium Synechocystis 6803: Inactivation of the psbA Gene Family. Plant Physiol 1987, 85, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, C.W. Electron transport and light-harvesting switches in cyanobacteria. Front. Plant Sci. 2014, 5, 7. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thurotte, A.; Seidel, T.; Jilly, R.; Kahmann, U.; Schneider, D. DnaK3 Is Involved in Biogenesis and/or Maintenance of Thylakoid Membrane Protein Complexes in the Cyanobacterium Synechocystis sp. PCC 6803. Life 2020, 10, 55. https://doi.org/10.3390/life10050055
Thurotte A, Seidel T, Jilly R, Kahmann U, Schneider D. DnaK3 Is Involved in Biogenesis and/or Maintenance of Thylakoid Membrane Protein Complexes in the Cyanobacterium Synechocystis sp. PCC 6803. Life. 2020; 10(5):55. https://doi.org/10.3390/life10050055
Chicago/Turabian StyleThurotte, Adrien, Tobias Seidel, Ruven Jilly, Uwe Kahmann, and Dirk Schneider. 2020. "DnaK3 Is Involved in Biogenesis and/or Maintenance of Thylakoid Membrane Protein Complexes in the Cyanobacterium Synechocystis sp. PCC 6803" Life 10, no. 5: 55. https://doi.org/10.3390/life10050055
APA StyleThurotte, A., Seidel, T., Jilly, R., Kahmann, U., & Schneider, D. (2020). DnaK3 Is Involved in Biogenesis and/or Maintenance of Thylakoid Membrane Protein Complexes in the Cyanobacterium Synechocystis sp. PCC 6803. Life, 10(5), 55. https://doi.org/10.3390/life10050055