Influence of the Depth on the Shape and Thickness of Nacre Tablets of Pinctada margaritifera Pearl Oyster, and on Oxygen Isotopic Composition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
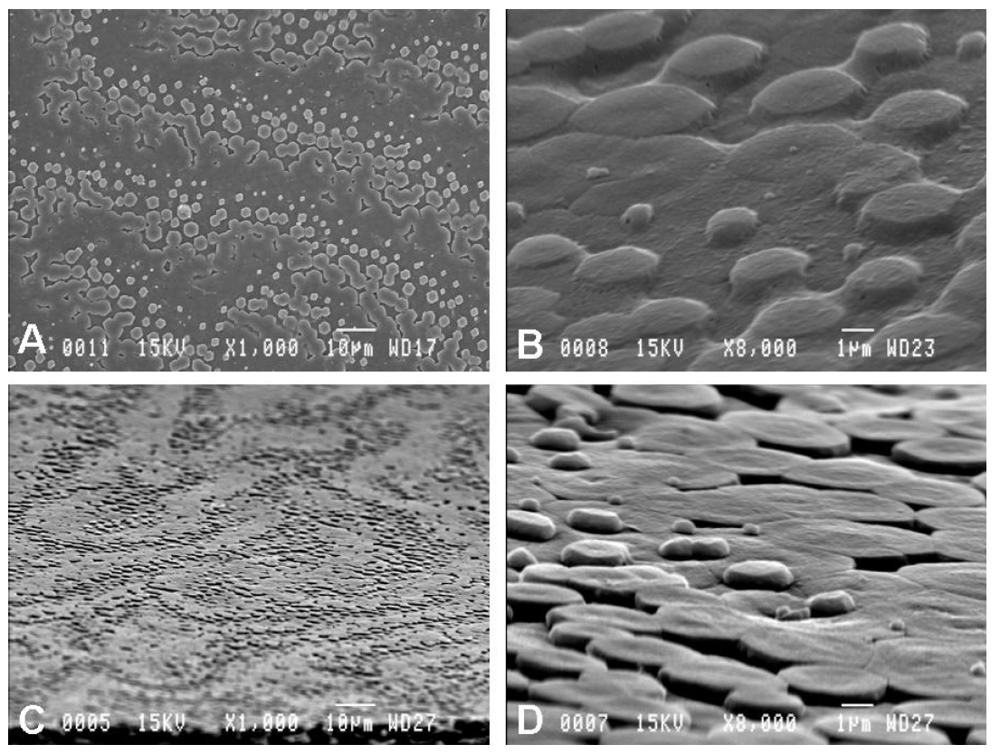
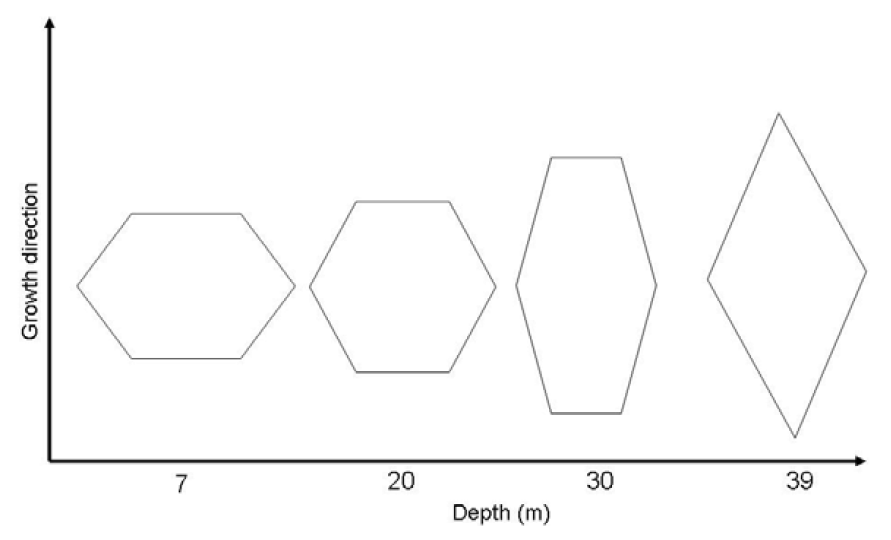
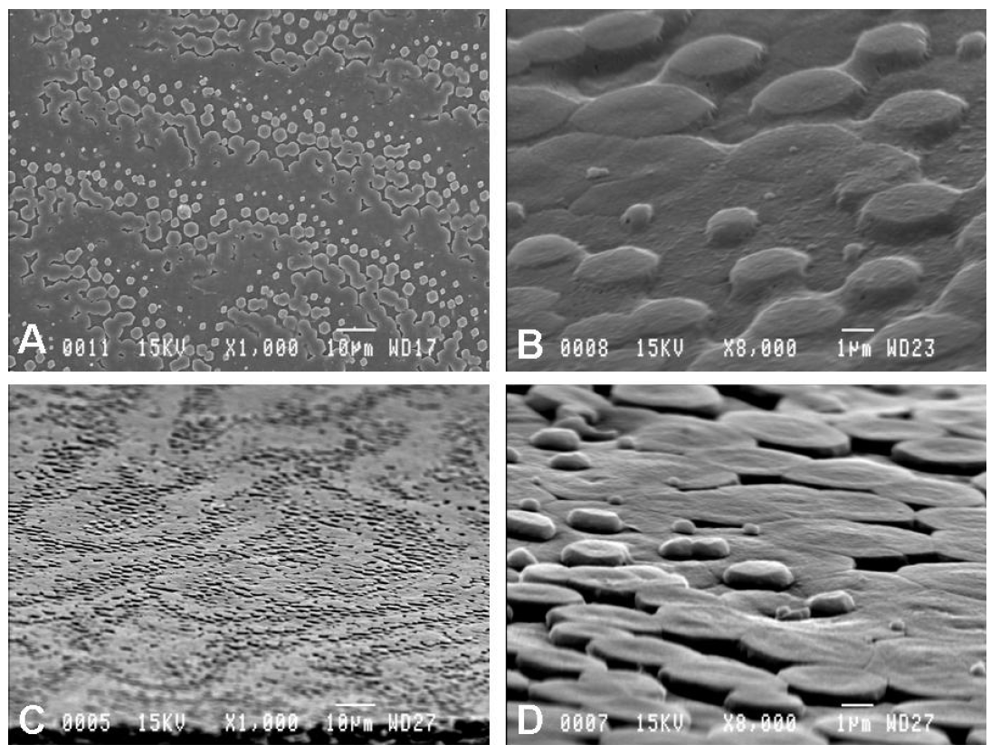
2.1. Scanning Electron Microscopy Observations


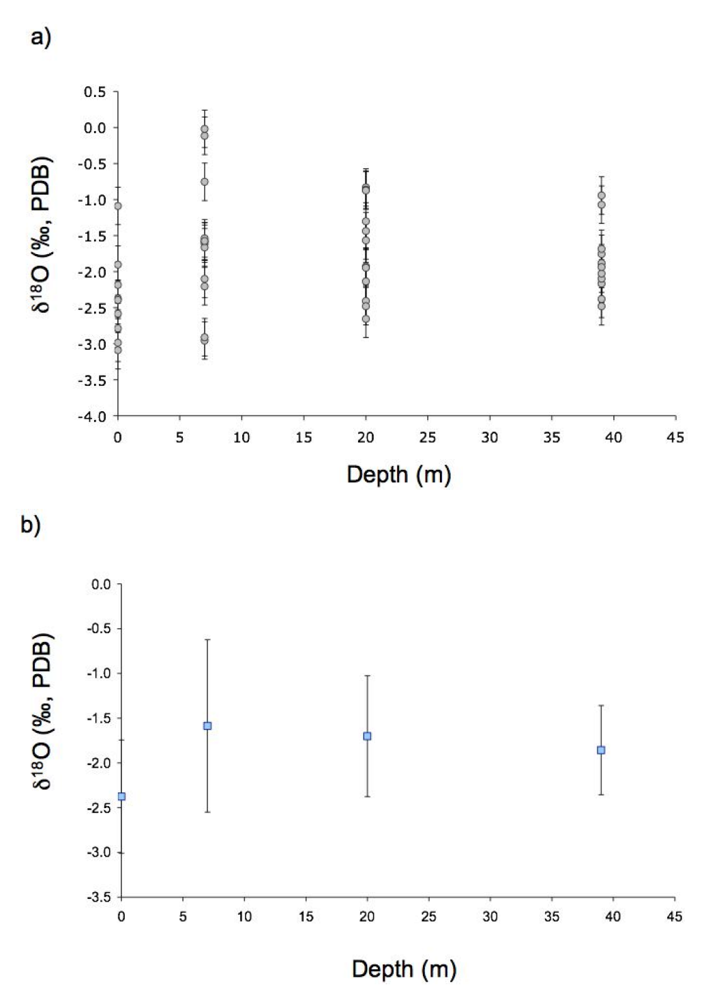
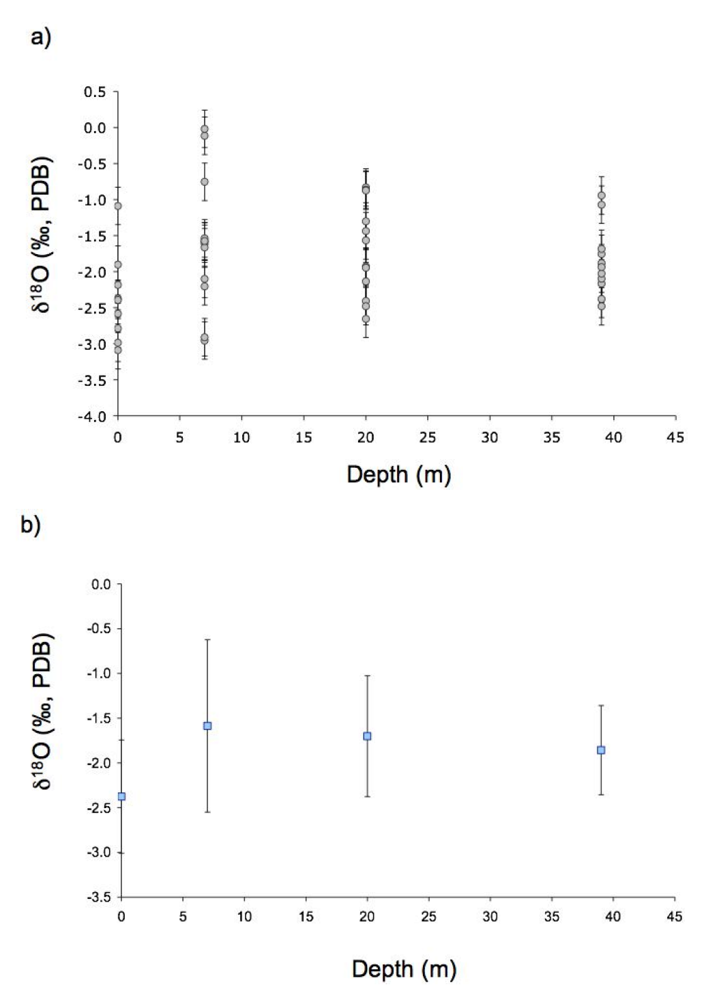
2.2. Oxygen Isotope Measurements
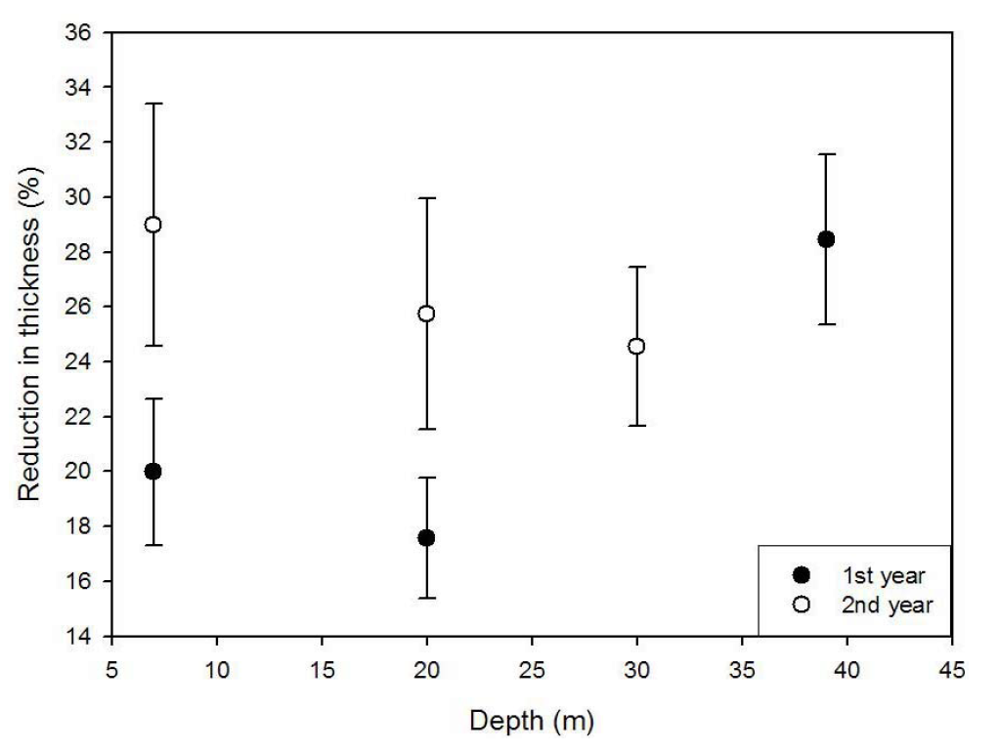
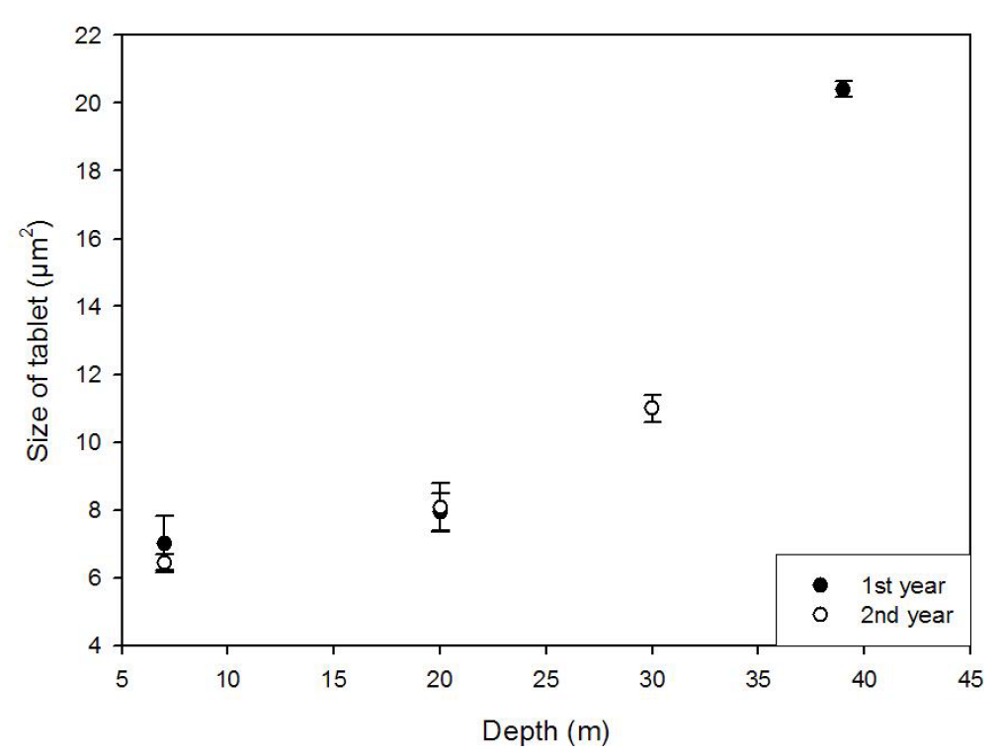
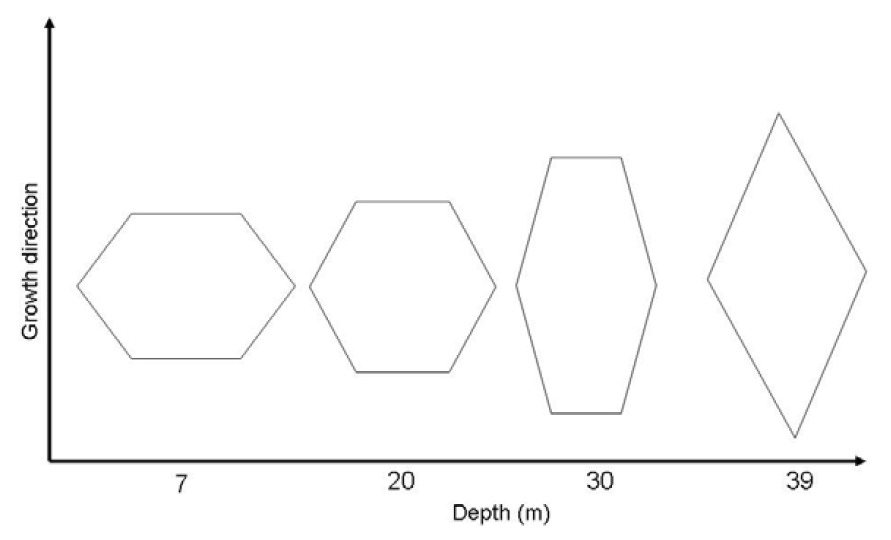
2.3. Modifications in Thickness and Size


3. Experimental Section
3.1. Experimental Testing
3.2. Scanning Electron Microscopy
3.3. Oxygen Isotope Measurements
4. Conclusions
Acknowledgements
References
- Hurwit, K.N.; Shigley, J.E.; Liu, Y. Iridescent colour of a shell mollusc Pinctada margaritifera caused by diffraction. Int. J. Optics 1999, 4(5), 177–182. [Google Scholar]
- Delesalle, B.; Charpy, L.; Le Borgne, R.; Adessi, L.; Legendre, L.; Sakka, A.; Pouvreau, S.; Niquil, N. Trophic web and carrying in a pearl oyster farming lagoon (Takapoto, French Polynesia). Aquat. Living Resour. 2001, 14, 165–174. [Google Scholar] [CrossRef]
- Prasil, V.; Pouvreau, S. Growth of the blacklip pearl oyster, Pinctada margaritifera, at nine culture sites of French Polynesia: synthesis of several sampling designs conducted between 1994 and 1999. Aquat. Living Resour 2001, 14(3), 155–163. [Google Scholar] [CrossRef]
- Wada, K. Nucleation and growth of aragonite crystals in the nacre of some bivalve molluscs. Biomineralization 1972, 6, 141–159. [Google Scholar]
- Urey, H.C. Thermodynamic properties of isotopic substances. J. Chem. Soc. 1947, 562–579. [Google Scholar] [CrossRef]
- Urey, H.C.; Lowenstam, H.A.; Buchsbaum, R.; Epstein, S. Carbonate-water isotopic temperature scale. Bull. Geol. Soc. Amer. 1951, 62, 417–426. [Google Scholar] [CrossRef]
- Urey, H.C.; Lowenstam, H.A.; Buchsbaum, R.; Epstein, S. Revised carbonate-water isotopic temperature scale. Bull. Geol. Soc. Amer. 1953, 64, 1315–1325. [Google Scholar] [CrossRef]
- McKinney, C.R.; Epstein, S.; Lowenstam, H.A.; Urey, H.C. Measurement of paleotemperatures and temperatures of the Upper Cretaceous of England, Denmark, and the Southeastern United States. Bull. Geol. Soc. Amer. 1951, 62, 399–416. [Google Scholar] [CrossRef]
- Bourrat, X.; Naslain, R.; Smith, D.C.; Mascarel, G.; Couté, A.; Lopez, E.; Rousseau, M. Sheet nacre growth mechanism: A Voronoi model. J. Structur. Biol 2005, 149, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Dove, P.M.; Weiner, S. An overview of biomineralization processes and the problem of the vital effec. In Biomineralization; Eds., Weiner, S., de Yoreo, J.J., Dove, P.M., Eds.; 2003; The Mineralogical Society of America: Washington, DC, USA. [Google Scholar]
- Ku, T.L.; Grossman, E.L. Oxygen and carbon isotope fractionation in biogenic aragonite—Temperature effects. Chem. Geol. 1986, 59, 59–74. [Google Scholar] [CrossRef]
- Dettman, D.L.; Schöne, B.R.; Flessa, K.W.; Goodwin, D.H. Cross-calibration of daily growth increments, stable isotope variation, and temperature in the Gulf of California bivalve mollusk Chione cortezi: Implication for paleoenvironmental analysis. Palaios 2001, 16, 387–398. [Google Scholar]
- Lopez, E.; Angellier, M.; Bourrat, X.; Gèze, M.; Meibom, A.; Rousseau, M. Dynamics of sheet nacre formation in bivalves. J. Structur. Biol. 2009, 165, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, M.L.; Pedrozo, H.A. Effects of hypergravity on statocyst development in embryonic Aplysia californica. Hear. Res. 1994, 79, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Rahmann, H.; Kappel, T.; Anken, R.H. Morphometry of fish inner ear otoliths after development at 3g hypergravity. Acta Oto-laryngol. 1998, 118, 534–539. [Google Scholar] [CrossRef]
- Becker, W.; Reelsen, O.; Marxen, J. Embryonic development of the freshwater snail Biomphalaria glabrata under µg conditions (STS-89 mission). J. Gravit. Physio. 2001, 8, 29–36. [Google Scholar]
- Rollion-Bard, C.; Blamart, D.; Cuif, J.-P.; Dauphin, Y. In situ measurements of oxygen isotopic composition in deep-sea coral, Lophelia pertusa: Re-examination of the current geochemical models of biomineralization. Geochim. Cosmochim. Acta 2010, 74, 1338–1349. [Google Scholar] [CrossRef]
- Champenois, M.; Mangin, M.; Rollion-Bard, C. Development and application of oxygen and carbon isotopic measurements of biogenic carbonates by ion microprobe. Geostand. Geoanal. Res. 2007, 31, 39–50. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rousseau, M.; Rollion-Bard, C. Influence of the Depth on the Shape and Thickness of Nacre Tablets of Pinctada margaritifera Pearl Oyster, and on Oxygen Isotopic Composition. Minerals 2012, 2, 55-64. https://doi.org/10.3390/min2010055
Rousseau M, Rollion-Bard C. Influence of the Depth on the Shape and Thickness of Nacre Tablets of Pinctada margaritifera Pearl Oyster, and on Oxygen Isotopic Composition. Minerals. 2012; 2(1):55-64. https://doi.org/10.3390/min2010055
Chicago/Turabian StyleRousseau, Marthe, and Claire Rollion-Bard. 2012. "Influence of the Depth on the Shape and Thickness of Nacre Tablets of Pinctada margaritifera Pearl Oyster, and on Oxygen Isotopic Composition" Minerals 2, no. 1: 55-64. https://doi.org/10.3390/min2010055