Symmetry Detection in Visual Impairment: Behavioral Evidence and Neural Correlates

Abstract
:
{kind=link}
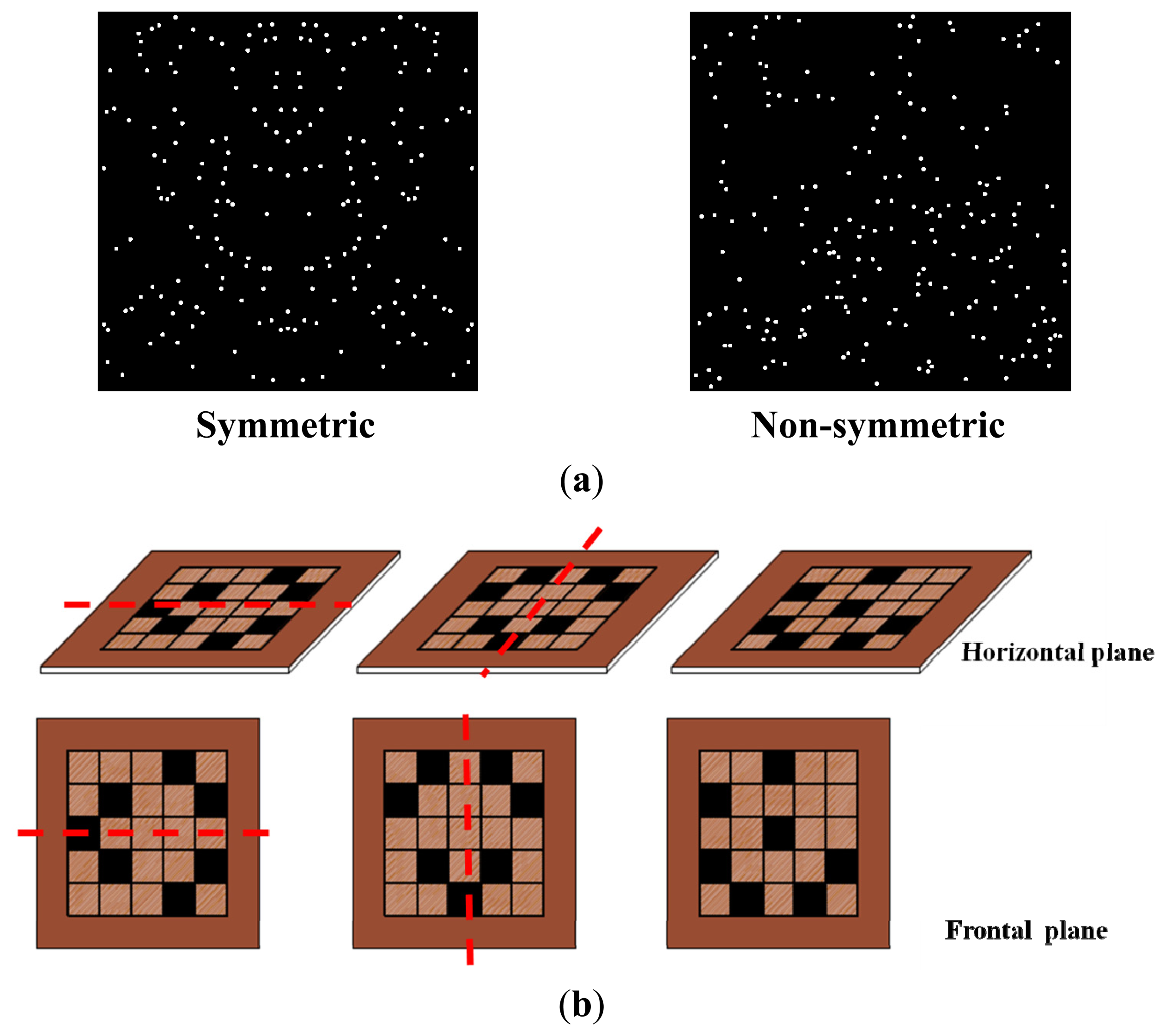{kind=link}
{kind=link}
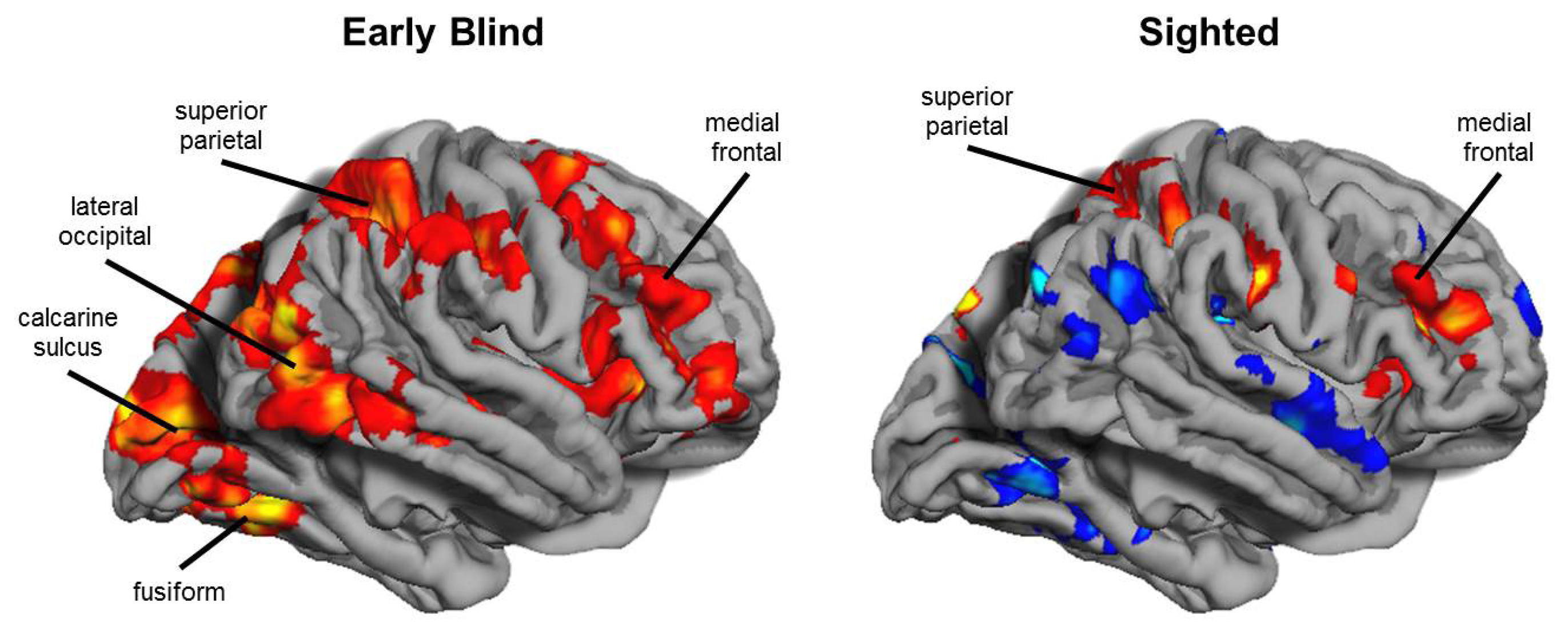{kind=link}
1. Introduction
2. Visual Symmetry Detection in Individuals with Impaired Binocular Vision
3. From Vision to Touch: Symmetry Detection in the Haptic Modality
3.1. Haptic Detection of Symmetry in Normally Sighted Participants
3.2. Haptic Detection of Symmetry in Blind Participants
4. Neural Correlates of Symmetry Detection in Sighted and Blind Individuals
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Treder, M.S. Behind the looking-glass: A review on human symmetry perception. Symmetry 2010, 2, 1510–1543. [Google Scholar]
- Barlow, H.B.; Reeves, B.C. The versatility and absolute efficiency of detecting mirror symmetry in random dot displays. Vis. Res. 1979, 19, 783–793. [Google Scholar]
- Driver, J.; Baylis, G.C.; Rafal, R.D. Preserved figure-ground segmentation and symmetry perception in visual neglect. Nature 1992, 360, 73–75. [Google Scholar]
- Wagemans, J. Parallel visual process in symmetry perception: Normality and pathology. Doc. Ophthalmol. 1998–1999, 95, 359–370. [Google Scholar]
- Machilsen, B.; Pauwels, M.; Wagemans, J. The role of vertical mirror symmetry in visual shape detection. J. Vis. 2009, 9, 1–11. [Google Scholar]
- Joung, W.; van der Zwan, R.; Latimer, C.R. Axes of symmetry produce tilt aftereffects. Spat. Vis. 1997, 13, 107–128. [Google Scholar]
- Joung, W.; Latimer, C.R. Tilt aftereffects generated by symmetrical dot patterns with two or four axes of symmetry. Spat. Vis. 2003, 16, 155–182. [Google Scholar]
- Dakin, S.C.; Herbert, A.M. The spatial region of integration for visual symmetry detection. Proc. Biol. Sci. 1998, 265, 659–664. [Google Scholar]
- Dakin, S.C.; Hess, R.F. The spatial mechanisms mediating symmetry perception. Vis. Res. 1997, 37, 2915–2930. [Google Scholar]
- Rainville, S.J.; Kingdom, F.A. The functional role of oriented spatial filters in the perception of mirror symmetry—Psychophysics and modeling. Vis. Res. 2000, 40, 2621–2644. [Google Scholar]
- Wade, T.J. The relationships between symmetry and attractiveness and mating relevant decisions and behavior: A review. Symmetry 2010, 2, 1081–1098. [Google Scholar]
- Wenderoth, P. The salience of vertical symmetry. Perception 1994, 23, 221–236. [Google Scholar]
- Herbert, A.M.; Humphrey, G.K. Bilateral symmetry detection: Testing a “callosal” hypothesis. Perception 1986, 25, 463–480. [Google Scholar]
- Gallace, A.; Spence, C. To what extent do Gestalt grouping principles influence tactile perception? Psychol. Bull. 2011, 137, 538–561. [Google Scholar]
- Wagemans, J. Detection of visual symmetries. Spat. Vis. 1995, 9, 9–32. [Google Scholar]
- Wong, A.M. New concepts concerning the neural mechanisms of amblyopia and their clinical implications. Can. J. Ophthalmol. 2012, 47, 399–409. [Google Scholar]
- Neville, H.; Bavelier, D. Human brain plasticity: Evidence from sensory deprivation and altered language experience. Prog. Brain Res. 2002, 138, 177–188. [Google Scholar]
- Martins Rosa, A.; Silva, M.F.; Ferreira, S.; Murta, J.; Castelo-Branco, M. Plasticity in the human visual cortex: An ophthalmology-based perspective. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Wenderoth, P. Monocular symmetry is neither necessary nor sufficient for the dichoptic perception of bilateral symmetry. Vis. Res. 2000, 40, 2097–2100. [Google Scholar]
- McKee, S.P.; Levi, D.M.; Movshon, J.A. The pattern of visual deficits in amblyopia. J. Vis. 2003, 3, 380–405. [Google Scholar]
- Levi, D.M.; Saarinen, J. Perception of mirror symmetry in amblyopic vision. Vis. Res. 2004, 44, 2475–2482. [Google Scholar]
- Hess, R.F.; McIlhagga, W.; Field, D.J. Contour integration in strabismic amblyopia: The sufficiency of an explanation based on positional uncertainty. Vis. Res. 1997, 37, 3145–3161. [Google Scholar]
- Mussap, A.J.; Levi, D.M. Amblyopic deficits in detecting a dotted line in noise. Vis. Res. 2000, 40, 3297–3307. [Google Scholar]
- Saarinen, J.; Levi, M.D. Perception of mirror symmetry reveals long-range interactions between orientation-selective cortical filters. Neuroreport 2000, 11, 2133–2138. [Google Scholar]
- Bona, S.; Herbert, A.; Toneatto, C.; Silvanto, J.; Cattaneo, Z. The causal role of the lateral occipital complex in visual mirror symmetry detection and grouping: An fMRI-guided TMS study. Cortex 2014, 51, 46–55. [Google Scholar]
- Cattaneo, Z.; Mattavelli, G.; Papagno, C.; Herbert, A.; Silvanto, J. The role of the human extrastriate visual cortex in mirror symmetry discrimination: A TMS-adaptation study. Brain Cogn. 2011, 77, 120–127. [Google Scholar]
- Sasaki, Y.; Vanduffel, W.; Knutsen, T.; Tyler, C.; Tootell, R. Symmetry activates extrastriate visual cortex in human and nonhuman primates. Proc. Natl. Acad. Sci. USA 2005, 102, 3159–3163. [Google Scholar]
- Tyler, C.W.; Baseler, H.A.; Kontsevich, L.L.; Likova, L.T.; Wade, A.R.; Wandell, B.A. Predominantly extra-retinotopic cortical response to pattern symmetry. Neuroimage 2005, 24, 306–314. [Google Scholar]
- Van der Zwan, R.; Leo, E.; Joung, W.; Latimer, C.; Wenderoth, P. Evidence that both area V1 and extrastriate visual cortex contribute to symmetry perception. Curr. Biol. 1998, 8, 889–892. [Google Scholar]
- Herbert, A.M.; Overbury, O.; Singh, J.; Faubert, J. Aging and bilateral symmetry detection. J. Gerontol. B Psychol. Sci. Soc. Sci. 2002, 57, 241–245. [Google Scholar]
- Habak, C.; Faubert, J. Larger effect of aging on the perception of higher-order stimuli. Vis. Res. 2000, 40, 943–950. [Google Scholar]
- Luu, J.Y.; Levi, D.M. Sensitivity to synchronicity of biological motion in normal and amblyopic vision. Vis. Res. 2013, 83, 9–18. [Google Scholar]
- Mansouri, B.; Hess, R.F. The global processing deficit in amblyopia involves noise segregation. Vis. Res. 2006, 46, 4104–4117. [Google Scholar]
- Steeves, J.K.; González, E.G.; Steinbach, M.J. Vision with one eye: A review of visual function following unilateral enucleation. Spat. Vis. 2008, 21, 509–529. [Google Scholar]
- Cattaneo, Z.; Bona, S.; Monegato, M.; Pece, A.; Vecchi, T.; Herbert, A.M.; Merabet, L.B. Symmetry perception in monocular blindness. Vis. Cogn. 2014. submitted for publication. [Google Scholar]
- Cattaneo, Z.; Fantino, M.; Silvanto, J.; Tinti, C.; Pascual-Leone, A.; Vecchi, T. Symmetry perception in the blind. Acta Psychol. (Amst.) 2010, 134, 398–402. [Google Scholar]
- Cattaneo, Z.; Vecchi, T.; Fantino, M.; Herbert, A.M.; Merabet, L.B. The effect of vertical and horizontal symmetry on memory for tactile patterns in late blind individuals. Atten. Percept. Psychophys. 2013, 75, 375–382. [Google Scholar]
- Bauer, C.; Yazzolino, L.; Hirsch, G.; Cattaneo, Z.; Vecchi, T.; Merabet, L.B. Neural correlates associated with superior tactile symmetry perception in the early blind. Cortex 2014. submitted for publication. [Google Scholar]
- Ballesteros, S.; Manga, D.; Reales, J.M. Haptic discrimination of bilateral symmetry in 2-dimensional and 3-dimensional unfamiliar displays. Percept. Psychophys. 1997, 59, 37–50. [Google Scholar]
- Ballesteros, S.; Millar, S.; Reales, J.M. Symmetry in haptic and in visual shape perception. Percept. Psychophys 1998, 60, 389–404. [Google Scholar]
- Ballesteros, S.; Reales, J.M. Visual and haptic discrimination of symmetry in unfamiliar displays extended in the z-axis. Perception 2004, 33, 315–327. [Google Scholar]
- Locher, P.J.; Simmons, R.W. Influence of stimulus symmetry and complexity upon haptic scanning strategies during detection, learning, and recognition tasks. Percept. Psychophys. 1978, 23, 110–116. [Google Scholar]
- Millar, S. Short-term serial tactual recall: Effects of grouping on tactually probed recall of Braille letters and nonsense shapes by blind children. Br. J. Psychol. 1978, 69, 17–24. [Google Scholar]
- Millar, S. Understanding and Representing Space: Theory and Evidence from Studies with Blind and Sighted Children; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Simmons, R.W.; Locher, P.J. Role of extended perceptual experience upon haptic perception of nonrepresentational shapes. Percept. Mot. Skills 1979, 48, 987–991. [Google Scholar]
- Jenkins, B. Spatial limits to the detection of transpositional symmetry in dynamic dot textures. J. Exp. Psychol. Hum. Percept. Perform. 1983, 9, 258–269. [Google Scholar]
- Labonté, F.; Shapira, Y.; Cohen, P.; Faubert, J. A model for global symmetry detection in dense images. Spat. Vis. 1995, 9, 33–55. [Google Scholar]
- Thinus-Blanc, C.; Gaunet, F. Representation of space in blind persons: Vision as a spatial sense? Psychol. Bull. 1997, 121, 20–42. [Google Scholar]
- Latimer, C.R. Eye-movement indices of form zerception: Some methods and preliminary results. In From Eye to Mind: Information Acquisition in Perception, Search and Reading; Groner, R., d'Ydewalle, G., Parham, R., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1990; pp. 41–57. [Google Scholar]
- Latimer, C.R.; Joung, W.; van der Zwan, R.; Beh, H. Modelling experiential and task effects on attentional proceses in symmetry detection. In Current Oculomotor Research: Physiological and Psychological Aspects; Becker, W., Mergner, T., Eds.; Plenum: New York, NY, USA, 1999; pp. 309–312. [Google Scholar]
- Rossi-Arnaud, C.; Pieroni, L.; Baddeley, A. Symmetry and binding in visuo-spatial working memory. Neuroscience 2006, 139, 393–400. [Google Scholar]
- Rossi-Arnaud, C.; Pieroni, L.; Spataro, P.; Baddeley, A. Working memory and individual differences in the encoding of vertical, horizontal and diagonal symmetry. Acta Psychol. (Amst.) 2012, 141, 122–132. [Google Scholar]
- Gentaz, E.; Hatwell, Y. Role of gravitational cues in the haptic perception of orientation. Percept. Psychophys. 1996, 58, 1278–1292. [Google Scholar]
- Gentaz, E.; Hatwell, Y. The haptic oblique effect in the perception of rod orientation by blind adults. Percept. Psychophys. 1998, 60, 157–167. [Google Scholar]
- Goldreich, D.; Kanics, I.M. Performance of blind and sighted humans on a tactile grating detection task. Percept. Psychophys. 2006, 68, 1363–1371. [Google Scholar]
- Théoret, H.; Merabet, L.; Pascual-Leone, A. Behavioural and neuroplastic changes in the blind: Evidence from functionally relevant cross-modal interactions. J. Physiol. Paris 2004, 98, 221–233. [Google Scholar]
- Burton, H.; Snyder, A.Z.; Conturo, T.E.; Akbudak, E.; Ollinger, J.M.; Raichle, M.E. Adaptive changes in early and late blind: A fMRI study of Braille reading. J. Neurophysiol. 2002, 87, 589–607. [Google Scholar]
- Ales, J.M.; Appelbaum, L.G.; Cottereau, B.R.; Norcia, A.M. The time course of shape discrimination in the human brain. Neuroimage 2013, 67, 77–88. [Google Scholar]
- Grill-Spector, K.; Kourtzi, Z.; Kanwisher, N. The lateral occipital complex and its role in object recognition. Vis. Res. 2001, 41, 1409–1422. [Google Scholar]
- Höfel, L.; Jacobsen, T. Electrophysiological indices of processing aesthetics: Spontaneous or intentional processes? Int. J. Psychophysiol. 2007, 65, 20–31. [Google Scholar]
- Höfel, L.; Jacobsen, T. Electrophysiological indices of processing symmetry and aesthetics: A result of judgment categorization or judgment report? J. Psychophysiol. 2007, 21, 9–21. [Google Scholar]
- Jacobsen, T.; Höfel, L. Descriptive and evaluative judgment processes: Behavioral and electrophysiological indices of processing symmetry and aesthetics. Cogn. Affect. Behav. Neurosci. 2003, 3, 289–299. [Google Scholar]
- Makin, A.D.; Wilton, M.M.; Pecchinenda, A.; Bertamini, M. Symmetry perception and affective responses: A combined EEG/EMG study. Neuropsychologia 2012, 50, 3250–3261. [Google Scholar]
- Makin, A.D.; Rampone, G.; Pecchinenda, A.; Bertamini, M. Electrophysiological responses to visuospatial regularity. Psychophysiology 2013. [Google Scholar] [CrossRef]
- Robertson, E.M.; Théoret, H.; Pascual-Leone, A. Studies in cognition: The problems solved and created by transcranial magnetic stimulation. Cogn. Neurosci. 2003, 15, 948–960. [Google Scholar]
- Verma, A.; van der Haegen, L.; Brysbaert, M. Symmetry detection in typically and atypically speech lateralized individuals: A visual half-field study. Neuropsychologia 2013, 51, 2611–2619. [Google Scholar]
- Christie, J.; Ginsberg, J.P.; Steedman, J.; Fridriksson, J.; Bonilha, L.; Rorden, C. Global versus local processing: Seeing the left side of the forest and the right side of the trees. Front. Hum. Neurosci. 2012, 6. [Google Scholar] [CrossRef]
- Yovel, G.; Levy, J.; Yovel, I. Hemispheric asymmetries for global and local visual perception: Effects of stimulus and task factors. J. Exp. Psychol. Hum. Percept. Perform. 2001, 27, 1369–1385. [Google Scholar]
- Latimer, C.R.; Stevens, C.J. Some remarks on wholes, parts and their perception. Psycholoquy. 1997, 8. Available online: http://www.cogsci.ecs.soton.ac.uk/cgi/psyc/newpsy?8.13 (accessed on 26 May 2014).
- Julesz, B.; Chang, J.J. Symmetry perception and spatial-frequency channels. Perception 1979, 8, 711–718. [Google Scholar]
- Jenkins, B. Redundancy in the perception of bilateral symmetry in dot textures. Percept. Psychophys. 1982, 32, 171–177. [Google Scholar]
- Peyrin, C.; Chauvin, A.; Chokron, S.; Marendaz, C. Hemispheric specialization for spatial frequency processing in the analysis of natural scenes. Brain Cogn. 2003, 53, 278–282. [Google Scholar]
- Pietrini, P.; Furey, M.L.; Ricciardi, E.; Gobbini, M.I.; Wu, W.H.; Cohen, L.; Guazzelli, M.; Haxby, J.V. Beyond sensory images: Object-based representation in the human ventral pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 5658–5663. [Google Scholar]
- Stilla, R.; Hanna, R.; Hu, X.; Mariola, E.; Deshpande, G.; Sathian, K. Neural processing underlying tactile microspatial discrimination in the blind: A functional magnetic resonance imaging study. J. Vis. 2008, 8, 1–19. [Google Scholar]
- Merabet, L.; Thut, G.; Murray, B.; Andrews, J.; Hsiao, S.; Pascual-Leone, A. Feeling by sight or seeing by touch? Neuron 2004, 42, 173–179. [Google Scholar]


© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cattaneo, Z.; Bona, S.; Bauer, C.; Silvanto, J.; Herbert, A.M.; Vecchi, T.; Merabet, L.B. Symmetry Detection in Visual Impairment: Behavioral Evidence and Neural Correlates. Symmetry 2014, 6, 427-443. https://doi.org/10.3390/sym6020427
Cattaneo Z, Bona S, Bauer C, Silvanto J, Herbert AM, Vecchi T, Merabet LB. Symmetry Detection in Visual Impairment: Behavioral Evidence and Neural Correlates. Symmetry. 2014; 6(2):427-443. https://doi.org/10.3390/sym6020427
Chicago/Turabian StyleCattaneo, Zaira, Silvia Bona, Corinna Bauer, Juha Silvanto, Andrew M. Herbert, Tomaso Vecchi, and Lotfi B. Merabet. 2014. "Symmetry Detection in Visual Impairment: Behavioral Evidence and Neural Correlates" Symmetry 6, no. 2: 427-443. https://doi.org/10.3390/sym6020427