Using a Chemical Genetic Screen to Enhance Our Understanding of the Antibacterial Properties of Silver



Abstract
1. Introduction
2. Materials and Methods
2.1. Escherichia coli Strains and Storage
2.2. Stock Ag Solution
2.3. Determination of the Minimal Inhibitory Concentration and Controls
2.4. Screening
2.5. Normalization
2.6. Data Mining and Analyses
3. Results and Discussion
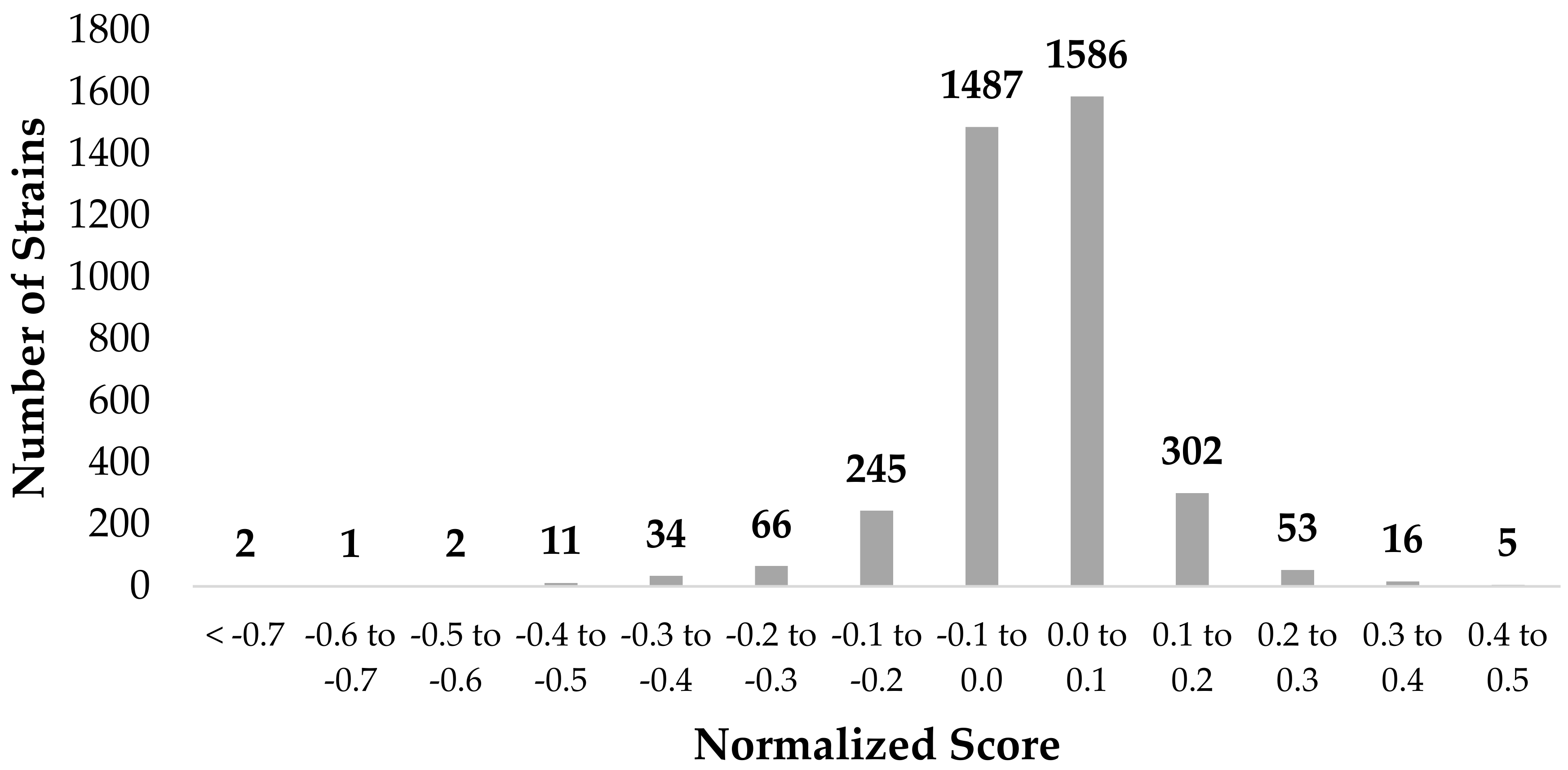
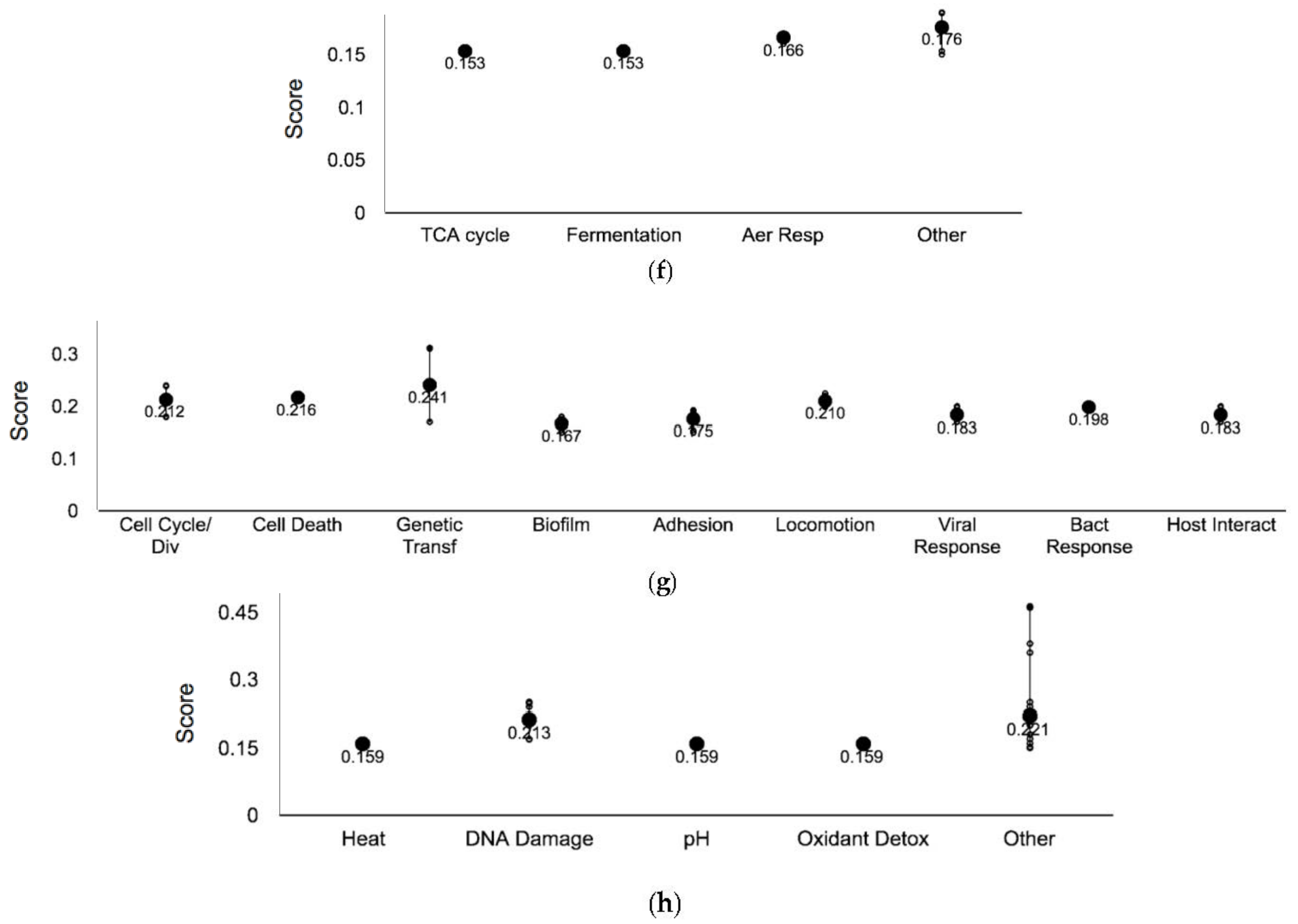
3.1. Genome-Wide Screen of Ag-Resistant and Ag-Sensitive Hits
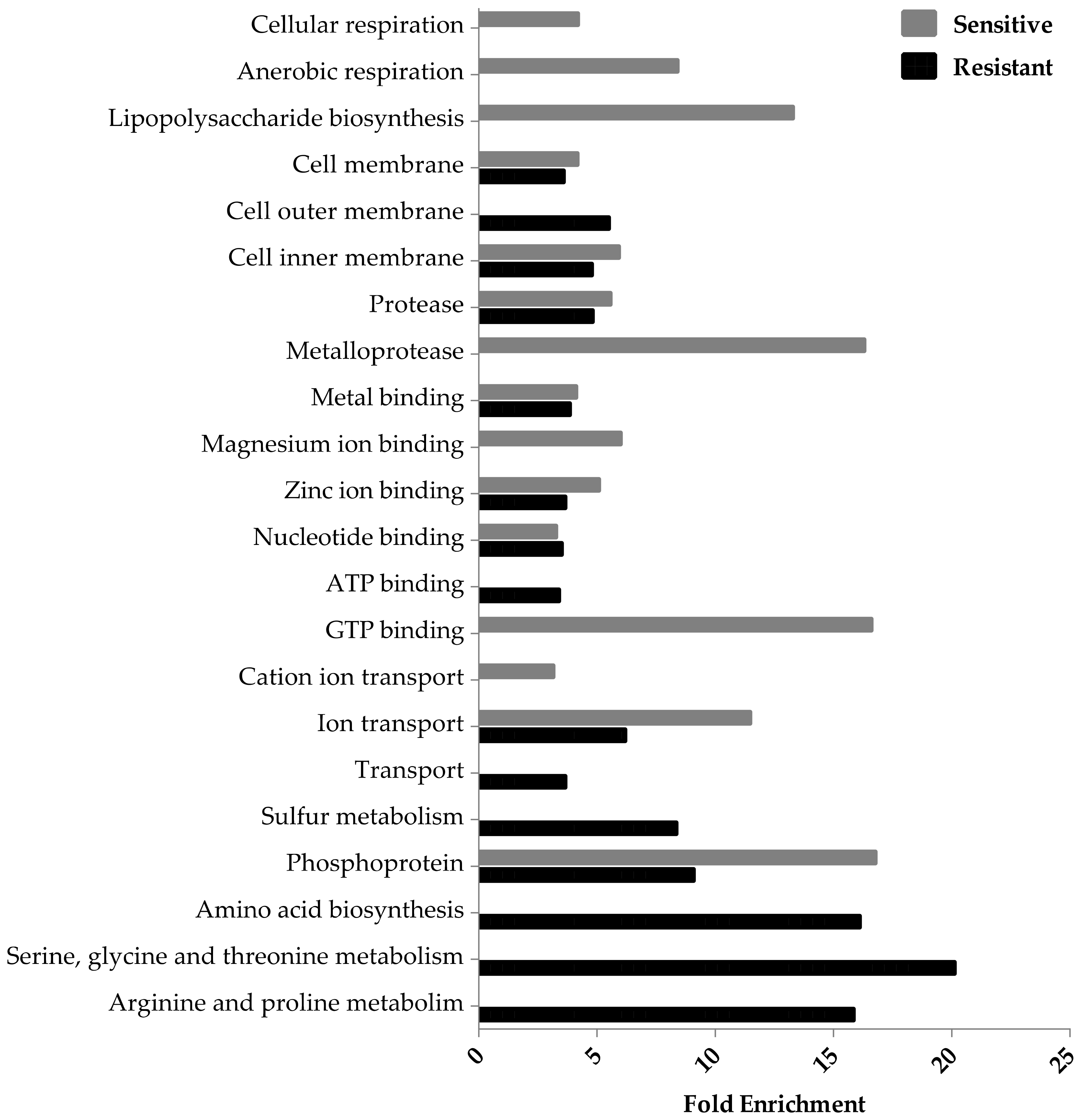
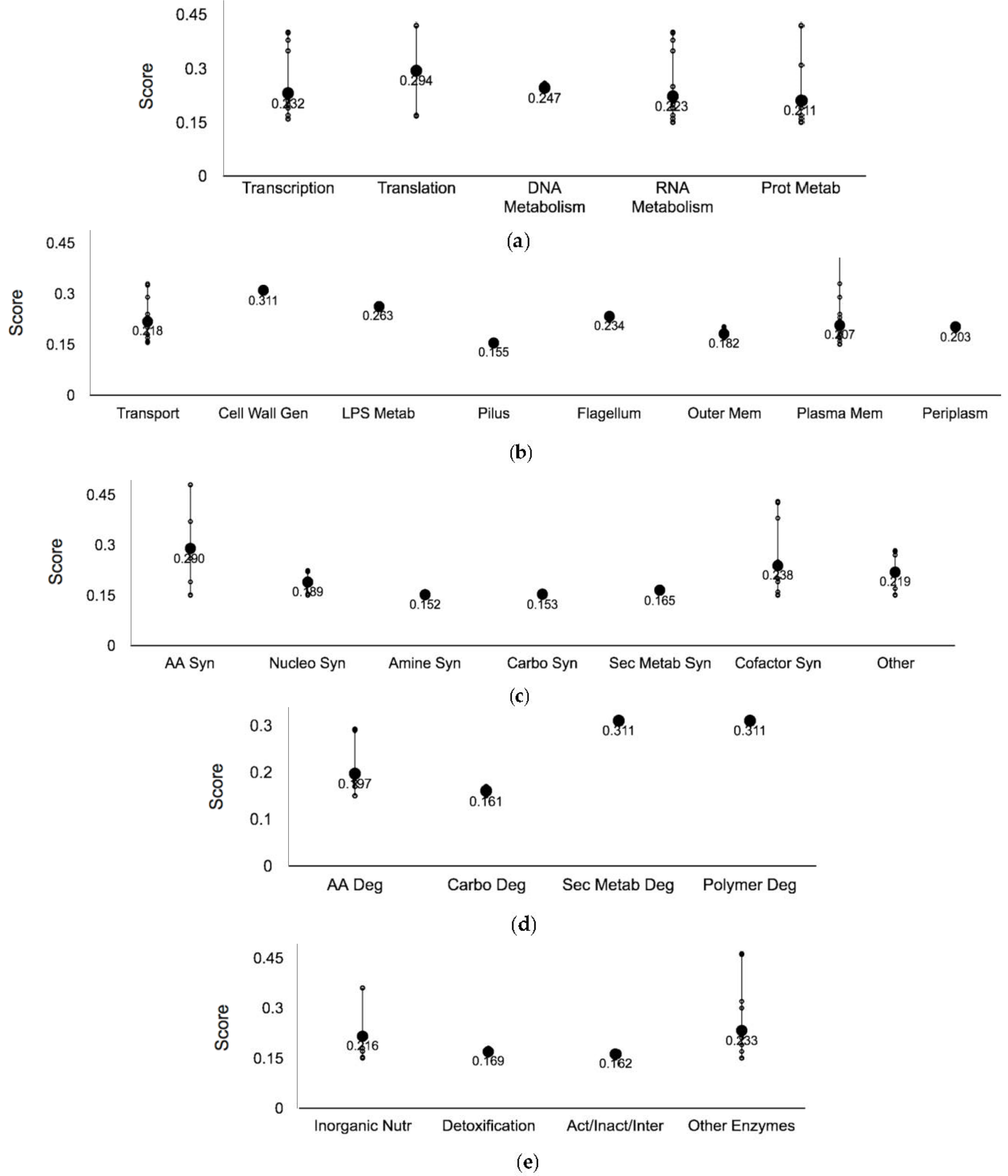
3.2. Ag-Resistant Gene Hits
3.2.1. Regulators of Gene Expression
3.2.2. Cell Membrane Proteins
3.2.3. Biosynthetic Enzymes
3.2.4. Catabolic Enzymes
3.2.5. Sulfur Metabolism Proteins
3.2.6. Biofilm Formation
3.2.7. DNA Damage and Repair
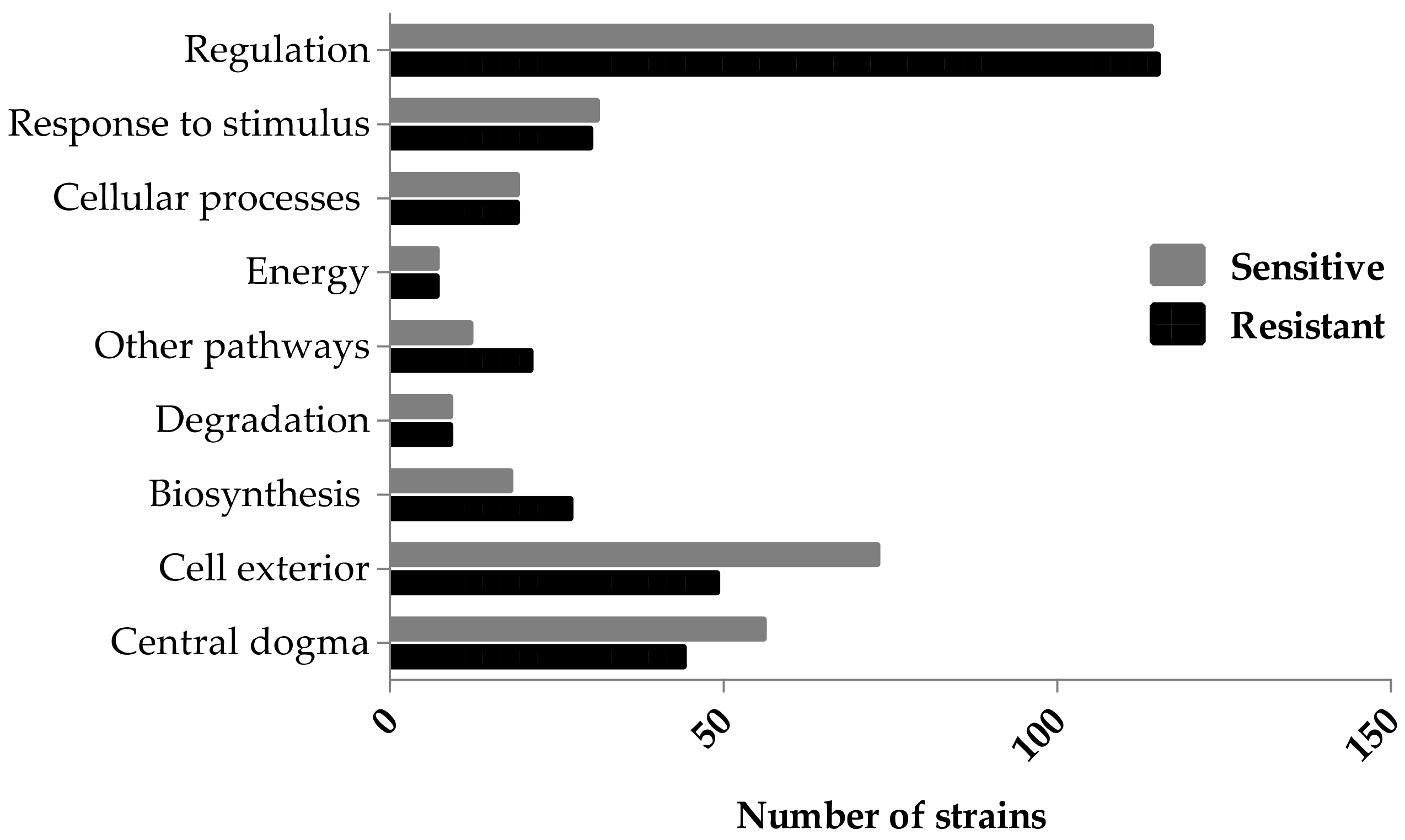
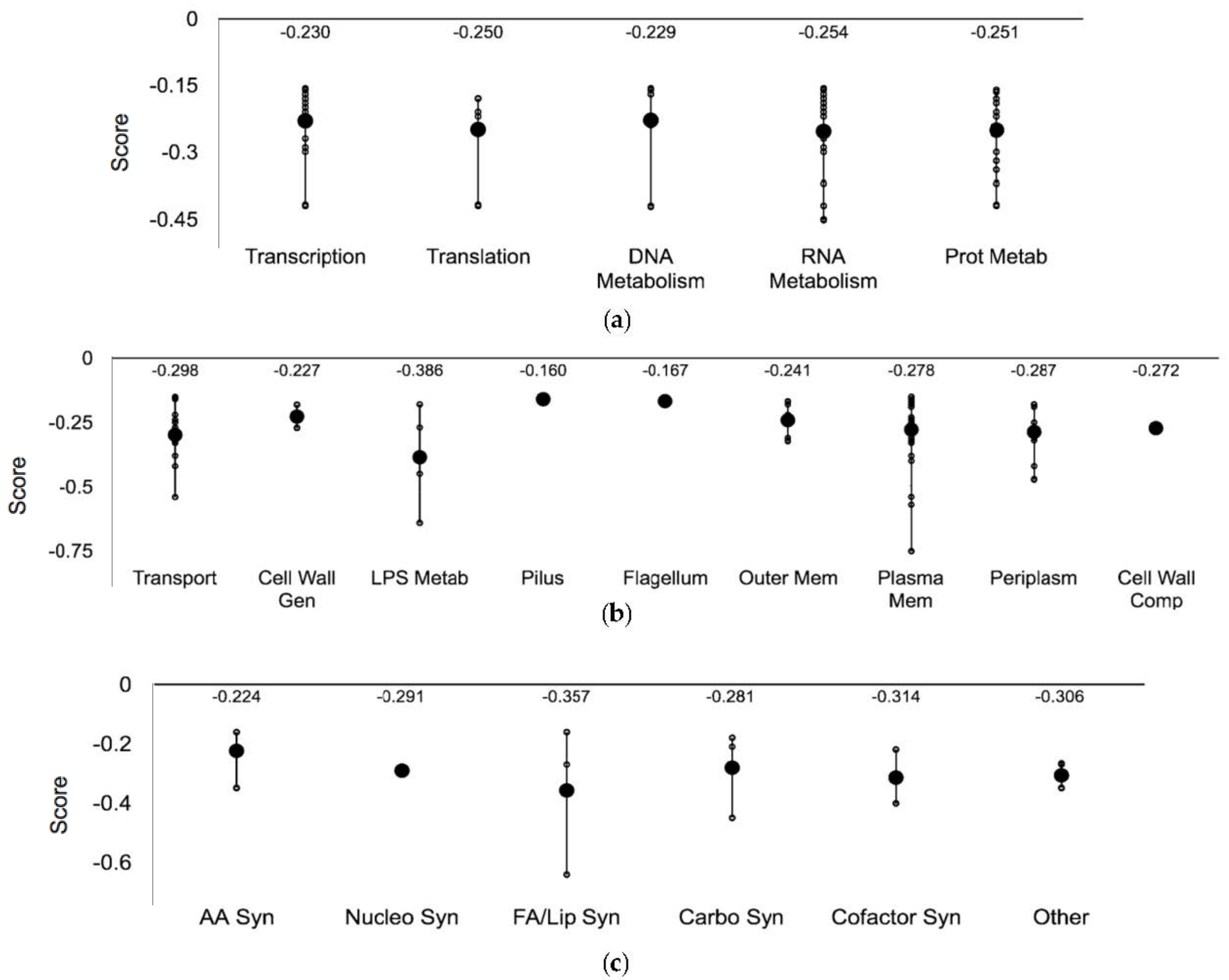
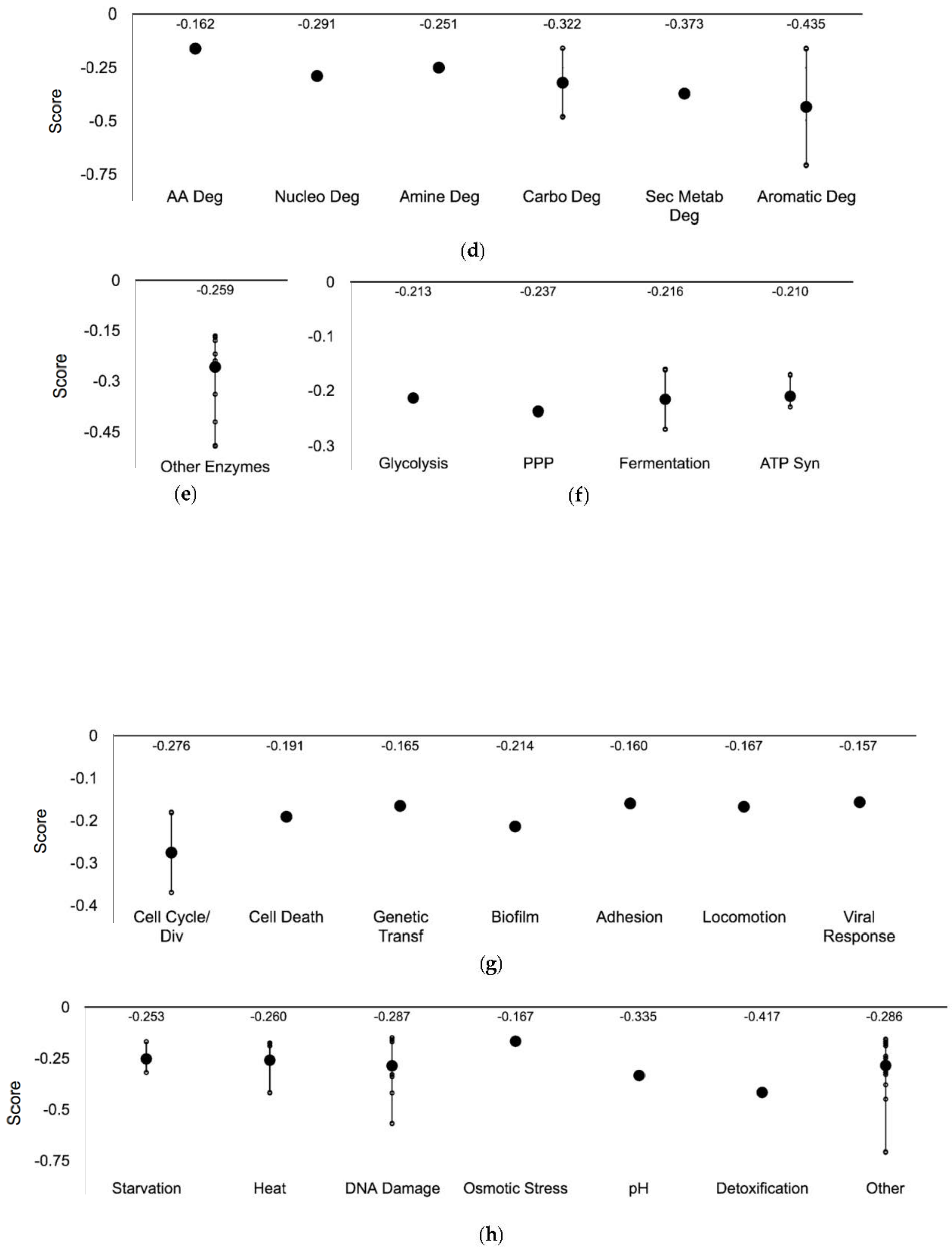
3.3. Ag-Sensitive Hits
3.3.1. Central Dogma and Cell Exterior Proteins
3.3.2. Lipopolysaccharide Biosynthetic Genes
3.3.3. Three Ag-Sensitive Hits Comprise the ATP Synthase Fo Complex
3.3.4. Oxidative Stress Response Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systems | Subsystems |
|---|---|
| Regulation | Signaling, sigma factor regulon, transcription factor, and transcription factor regulons |
| Response to Stimulus | Starvation, heat, cold, DNA damage, pH, detoxification, osmotic stress, and other |
| Cellular processes | Cell cycle and division, cell death, genetic transfer, biofilm formation, quorum sensing, adhesion, locomotion, viral response, response to bacterium, host interactions with host, other pathogenesis proteins |
| Energy | Glycolysis, the pentose phosphate pathway, the TCA cycle, fermentation, and aerobic and anaerobic respiration |
| Other pathways | Detoxification, inorganic nutrient metabolism, macromolecule modification, activation/inactivation/interconversion, and other enzymes |
| Degradation | Amino acids, nucleotide, amine, carbohydrate/carboxylate, secondary metabolite, alcohol, polymer and aromatic, the cell exterior, and regulation |
| Biosynthesis | Amino acids, nucleotides, fatty acid/lipid amines, carbohydrate/carboxylates, cofactors, secondary metabolites, and other pathways |
| Cell exterior | Transport, cell wall biogenesis and organization, lipopolysaccharide metabolism, pilus, flagellar, outer and inner membrane, periplasm, and cell wall components |
| Central Dogma | Transcription, translation, DNA metabolism, RNA metabolism, protein metabolism and protein folding and secretion |
References
- Turner, R.J. Metal-Based Antimicrobial Strategies. Microb. Biotechnol. 2017, 10, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.W. History of the Medical Use of Silver. Surg. Infect. 2009, 10, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Melaiye, A.; Youngs, W.J. Silver and Its Application as an Antimicrobial Agent. Expert Opin. Ther. Pat. 2005, 15, 125–130. [Google Scholar] [CrossRef]
- Lemire, J.A.; Kalan, L.; Bradu, A.; Turner, R.J. Silver Oxynitrate, an Unexplored Silver Compound with Antimicrobial and Antibiofilm Activity. Antimicrob. Agents Chemother. 2015, 59, 4031–4039. [Google Scholar] [CrossRef] [PubMed]
- Politano, A.D.; Campbell, K.T.; Rosenberger, L.H.; Sawyer, R.G. Use of Silver in the Prevention and Treatment of Infections: Silver Review. Surg. Infect. 2013, 14, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Gugala, N.; Lemire, J.A.; Turner, R.J. The Efficacy of Different Anti-Microbial Metals at Preventing the Formation of, and Eradicating Bacterial Biofilms of Pathogenic Indicator Strains. J. Antibiot. 2017, 70, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R.I. A Brief History of the Antibiotic Era: Lessons Learned and Challenges for the Future. Front. Microbiol. 2010, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Guidos, R.; Gilbert, D.; Bradley, J.; Boucher, H.W.; Scheld, W.M.; Bartlett, J.G.; Edwards, J. The Epidemic of Antibiotic-Resistant Infections: A Call to Action for the Medical Community from the Infectious Diseases Society of America. Clin. Infect. Dis. 2008, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- French, G.L. The Continuing Crisis in Antibiotic Resistance. Int. J. Antimicrob. Agents 2010, 36, S3–S7. [Google Scholar] [CrossRef]
- Neu, H.C. The Crisis in Antibiotic Resistance. Science 1992, 257, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Rigo, C.; Roman, M.; Munivrana, I.; Vindigni, V.; Azzena, B.; Barbante, C.; Cairns, W.R.L. Characterization and Evaluation of Silver Release from Four Different Dressings Used in Burns Care. Burns 2012, 38, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Sataev, M.S.; Koshkarbaeva, S.T.; Tleuova, A.B.; Perni, S.; Aidarova, S.B.; Prokopovich, P. Novel Process for Coating Textile Materials with Silver to Prepare Antimicrobial Fabrics. Colloids Surf. A Physicochem. Eng. Asp. 2014, 442, 146–151. [Google Scholar] [CrossRef]
- George, N.; Faoagali, J.; Muller, M. Silvazine® (Silver Sulfadiazine and Chlorhexidine) Activity against 200 Clinical Isolates. Burns 1997, 23, 493–495. [Google Scholar] [CrossRef]
- Guzman, M.; Dille, J.; Godet, S. Synthesis and Antibacterial Activity of Silver Nanoparticles against Gram-Positive and Gram-Negative Bacteria. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Paladini, F.; Pollini, M.; Talà, A.; Alifano, P.; Sannino, A. Efficacy of Silver Treated Catheters for Haemodialysis in Preventing Bacterial Adhesion. J. Mater. Sci. Mater. Med. 2012, 23, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Münstedt, H. Silver Ion Release from Antimicrobial Polyamide/silver Composites. Biomaterials 2005, 26, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Gordon, O.; Slenters, T.V.; Brunetto, P.S.; Villaruz, A.E.; Sturdevant, D.E.; Otto, M.; Landmann, R.; Fromm, K.M. Silver Coordination Polymers for Prevention of Implant Infection: Thiol Interaction, Impact on Respiratory Chain Enzymes, and Hydroxyl Radical Induction. Antimicrob. Agents Chemother. 2010, 54, 4208–4218. [Google Scholar] [CrossRef] [PubMed]
- Morones-Ramirez, J.R.; Winkler, J.A.; Spina, C.S.; Collins, J.J. Silver Enhances Antibiotic Activity against Gram-Negative Bacteria. Sci. Transl. Med. 2013, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial Activity of Metals: Mechanisms, Molecular Targets and Applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Turner, R.J. Multimetal Resistance and Tolerance in Microbial Biofilms. Nat. Rev. Microbiol. 2007, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Ivask, A.; Elbadawy, A.; Kaweeteerawat, C.; Boren, D.; Fischer, H.; Ji, Z.; Chang, C.H.; Liu, R.; Tolaymat, T.; Telesca, D.; et al. Toxicity Mechanisms in Escherichia coli Vary for Silver Nanoparticles and Differ from Ionic Silver. ACS Nano 2014, 8, 374–386. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, J.S.; Shaw, A.M. Differential Gene Regulation in the Ag Nanoparticle and Ag+ Induced Silver Stress Response in Escherichia coli: A Full Transcriptomic Profile. Nanotoxicology 2014, 8, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Saulou-Bérion, C.; Gonzalez, I.; Enjalbert, B.; Audinot, J.N.; Fourquaux, I.; Jamme, F.; Cocaign-Bousquet, M.; Mercier-Bonin, M.; Girbal, L. Escherichia coli under Ionic Silver Stress: An Integrative Approach to Explore Transcriptional, Physiological and Biochemical Responses. PLoS ONE 2015, 10, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2. [Google Scholar] [CrossRef] [PubMed]
- Franke, S.; Grass, G.; Nies, D.H. The Product of the ybdE Gene of the Escherichia coli Chromosome Is Involved in Detoxification of Silver Ions. Microbiology 2001, 147, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Wagih, O.; Usaj, M.; Baryshnikova, A.; VanderSluis, B.; Kuzmin, E.; Costanzo, M.; Myers, C.L.; Andrews, B.J.; Boone, C.M.; Parts, L. SGAtools: One-Stop Analysis and Visualization of Array-Based Genetic Interaction Screens. Nucleic Acids Res. 2013, 41, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Keseler, I.M.; Mackie, A.; Santos-Zavaleta, A.; Billington, R.; Bonavides-Martínez, C.; Caspi, R.; Fulcher, C.; Gama-Castro, S.; Kothari, A.; Krummenacker, M.; et al. The EcoCyc Database: Reflecting New Knowledge about Escherichia coli K-12. Nucleic Acids Res. 2017, 45, D543–D550. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING Database in 2017: Quality-Controlled Protein-Protein Association Networks, Made Broadly Accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Boël, G.; Smith, P.C.; Ning, W.; Englander, M.T.; Chen, B.; Hashem, Y.; Testa, A.J.; Fischer, J.J.; Wieden, H.J.; Frank, J.; et al. The ABC-F Protein EttA Gates Ribosome Entry into the Translation Elongation Cycle. Nat. Struct. Mol. Biol. 2014, 21, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Liu, Y.; Mathieu, J.; Wang, J.; Zhu, D.; Alvarez, P.J. Elucidating the Genetic Basis for Escherichia coli Defense against Silver Toxicity Using Mutant Arrays. Environ. Toxicol. Chem. 2014, 33, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Goldberg, A.L. Purification and Characterization of Protease So, a Cytoplasmic Serine Protease in Escherichia coli. J. Bacteriol. 1983, 170, 921–926. [Google Scholar]
- Sondi, I.; Salopek-Sondi, B. Silver Nanoparticles as Antimicrobial Agent: A Case Study on E. coli as a Model for Gram-Negative Bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Koo, H.C.; Kim, K.W.; Shin, S.; Kim, S.H.; Park, Y.H. Antibacterial Activity and Mechanism of Action of the Silver Ion in Staphylococcus aureus and Escherichia coli. Appl. Environ. Microbiol. 2008, 74, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Radzig, M.A.; Nadtochenko, V.A.; Koksharova, O.A.; Kiwi, J.; Lipasova, V.A.; Khmel, I.A. Antibacterial Effects of Silver Nanoparticles on Gram-Negative Bacteria: Influence on the Growth and Biofilms Formation, Mechanisms of Action. Colloids Surf. B Biointerfaces 2013, 102, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Beard, S.J.; Hashim, R.; Wu, G.; Binet, M.R.B.; Hughes, M.N.; Poole, R.K. Evidence for the Transport of Zinc(II) Ions via the Pit Inorganic Phosphate Transport System in Escherichia coli. FEMS Microbiol. Lett. 2000, 184, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Skare, J.T.; Postle, K. Evidence for a TonB-Dependent Energy Transduction Complex in Escherichia coli. Mol. Microbiol. 1991, 5, 2883–2890. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, R.; Horie, T.; Katoh, A.; Kaino, Y.; Tanabe, H.; Noda, M. Isolation and Characterization of the Heat-Responsive Genes in Escherichia coli. Biosci. Biotechnol. Biochem. 1996, 60, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Parr, R.G.; Pearson, R.G. Absolute Hardness: Companion Parameter to Absolute Electronegativity. J. Am. Chem. Soc. 1983, 105, 7512–7516. [Google Scholar] [CrossRef]
- Bhatia, R.; Calvo, K.C. The Sequencing, Expression, Purification, and Steady-State Kinetic Analysis of Quinolinate Phosphoribosyl Transferase from Escherichia coli. Arch. Biochem. Biophys. 1996, 325, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Cicchillo, R.M.; Tu, L.; Stromberg, J.A.; Hoffart, L.M.; Krebs, C.; Booker, S.J. Escherichia coli Quinolinate Synthetase Does Indeed Harbor a [4Fe-4S] Cluster. J. Am. Chem. Soc. 2015, 127, 7310–7311. [Google Scholar] [CrossRef]
- Xu, F.F.; Imlay, J.A. Silver(I), Mercury(II), Cadmium(II), and Zinc(II) Target Exposed Enzymic Iron-Sulfur Clusters When They Toxify Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 3614–3621. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, S.; Imlay, J.A. Two Sources of Endogenous H2O2 in Escherichia coli. Mol. Microbiol. 2011, 75, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Templin, M.F.; Ursinus, A.; Höltje, J.V. A Defect in Cell Wall Recycling Triggers Autolysis during the Stationary Growth Phase of Escherichia coli. EMBO J. 1999, 18, 4108–4117. [Google Scholar] [CrossRef] [PubMed]
- Goodell, E.W.; Schwarz, U. Release of Cell Wall Peptides into Culture Medium by Exponentially Growing Escherichia coli. J. Bacteriol. 1985, 162, 391–397. [Google Scholar] [PubMed]
- Raghunath, A.; Perumal, E. Metal Oxide Nanoparticles as Antimicrobial Agents: A Promise for the Future. Int. J. Antimicrob. Agents 2017, 49, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Berendt, U.; Haverkamp, T.; Prior, A.; Schwenn, J.D. Reaction Mechanism of Thioredoxin: 3′-Phospho-adenylylsulfate Reductase Investigated by Site-Directed Mutagenesis. Eur. J. Biochem. 1995, 233, 347–356. [Google Scholar] [CrossRef] [PubMed]
- White, D.C.; Frerman, F.E. Extraction, Characterization, and Cellular Localization of the Lipids of Staphylococcus aureus. J. Bacteriol. 1967, 94, 1854–1867. [Google Scholar] [PubMed]
- Siegel, L.M.; Rueger, D.C.; Barber, M.J.; Krueger, R.J.; Orme-Johnson, N.R.; Orme-Johnson, W.H. Escherichia coli Sulfite Reductase Hemoprotein Subunit. J. Biol. Chem. 1982, 257, 6348–6350. [Google Scholar]
- Stewart, P.S.; William Costerton, J. Antibiotic Resistance of Bacteria in Biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Harrison, J.J.; Turner, R.J.; Ceri, H. High-Throughput Metal Susceptibility Testing of Microbial Biofilms. BMC Microbiol. 2005, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Turner, R.J.; Ceri, H. Persister Cells, the Biofilm Matrix and Tolerance to Metal Cations in Biofilm and Planktonic Pseudomonas aeruginosa. Environ. Microbiol. 2005, 7, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Shafreen, R.B.; Seema, S.; Ahamed, A.P.; Thajuddin, N.; Ali Alharbi, S. Inhibitory Effect of Biosynthesized Silver Nanoparticles from Extract of Nitzschia Palea Against Curli-Mediated Biofilm of Escherichia coli. Appl. Biochem. Biotechnol. 2017, 183, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.W.; Rafferty, J.B.; Artymiuk, P.J.; Lloyd, R.G. Insights into the Mechanisms of Homologous Recombination from the Structure of RuvA. Curr. Opin. Struct. Biol. 1997, 7, 798–803. [Google Scholar] [CrossRef]
- Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.Y. Use of Hydrogen Peroxide as a Biocide: New Consideration of Its Mechanisms of Biocidal Action. J. Antimicrob. Chemother. 2012, 67, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Yang, F.; Long, F.; Reyon, D.; Routh, M.D.; W, D.; Mokhtari, A.K.; Ornam, J.D. Van; Rabe, K.L.; Hoy, J.A.; et al. Crystal Structure of the Membrane Fusion Protein CusB from Escherichia coli. J. Mol. Biol. 2009, 393, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Fralick, A. Evidence That TolC Is Required for Functioning of the Mar/AcrAB Efflux Pump of Escherichia coli. Am. Soc. Microbiol. 1996, 178, 5803–5805. [Google Scholar] [CrossRef]
- Holt, K.B.; Bard, A.J. Interaction of Silver(I) Ions with the Respiratory Chain of Escherichia coli: An Electrochemical and Scanning Electrochemical Microscopy Study of the Antimicrobial Mechanism of Micromolar Ag. Biochemistry 2005, 44, 13214–13223. [Google Scholar] [CrossRef] [PubMed]
- DiRienzo, J.M.; Nakamura, K.; Inouye, M. The Outer Membrane Porteins of Gram-Negative Bacteria: Biosynthesis, Assembly, and Functions. Proteins 1978, 47, 481–532. [Google Scholar] [CrossRef]
- Helbig, K.; Bleuel, C.; Krauss, G.J.; Nies, D.H. Glutathione and Transition-Metal Homeostasis in Escherichia coli. J. Bacteriol. 2008, 190, 5431–5438. [Google Scholar] [CrossRef] [PubMed]
- Pomposiello, P.J.; Koutsolioutsou, A.; Carrasco, D.; Demple, B. SoxRS-Regulated Expression and Genetic Analysis of the yggX Gene of Escherichia coli. Society 2003, 185, 6624–6632. [Google Scholar] [CrossRef]
- Link, A.J.; Robison, K.; Church, G.M. Comparing the Predicted and Observed Properties of Proteins Encoded in the Genome of Escherichia coli K-12. Electrophoresis 1997, 18, 1259–1313. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, N.; Cooper, C.E.; Poole, R.K. Spectroscopic Studies on an Oxygen-Binding Haemoglobin-like Flavohaemoprotein from Escherichia Coli. Biochem. J. 1992, 288, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A Review of the Scientific Evidence for Biofilms in Wounds. Wound Repair Regen. 2012, 20, 647–657. [Google Scholar] [CrossRef] [PubMed]







| System | Subsystem | Gene 3 |
|---|---|---|
| Central Dogma | Transcription | alaS crp dicC gadE gcvR lysR putA yciT yhjB yiif yjiR |
| Translation | alaS ettA | |
| DNA Metabolism | cffC dam recT | |
| RNA Metabolism | rluF alaS gluQ trmL crp dicC gadE gcvR lysR ogrK putA yciT yhjB yiif yjiR yjtD | |
| Protein Metabolism | argE envZ lipB sdhE ldcA pepB prc rhsB rzpD | |
| Cell Exterior | Transport | malE nhaB exbB btuB dppF glcA ompG lptB mngA yejF |
| Cell wall biogenesis/organization | idcA | |
| Lipopolysaccharide Metabolism | wcaI | |
| Pilus | yraK | |
| Flagellum | fliL fliR | |
| Outer membrane | bbtuB csgF nlpE ompA ompG rhsB | |
| Plasma membrane | agaD cyoC cysQ damX dppF envZ ettA exbB fliL fliR glcA IptB malE mngA nhaB ppx prc putA yaiP yccF yejF ygdD yifK yojI yqfA | |
| Periplasm | malE nlpE prc | |
| Biosynthesis | Amino acid biosynthesis | argE cysk serC proC serA serC metL trpB trpD |
| Nucleotide biosynthesis | dcd pyrF | |
| Amine biosynthesis | gss | |
| Carbohydrate biosynthesis | mdh | |
| Secondary metabolite biosynthesis | fldB | |
| Cofactor biosynthesis | bioC bioF nudB lipB nadA nadB nadC gss thiS serC | |
| Other | aroC metL argE alaS | |
| Degradation | Amino acid degradation | astA cysK gadA putA |
| Carbohydrate degradation | galM yigL glcE | |
| Secondary metabolite degradation | idcA | |
| Polymer degradation | idcA | |
| Other pathways | Inorganic nutrient metabolism | cysC cysD cysH cysI |
| Detoxification | gadA sodA | |
| Activation/inactivation/interconversion | cysC cysD | |
| Other | ahpF bglB cysQ dam gluQ pepB ppx prc purU rluF trmL yfaU yjhG | |
| Energy | TCA cycle | mdh |
| Fermentation | mdh | |
| Aerobic respiration | cyoC putA | |
| Other | bioC bioF mdh | |
| Cellular process | Cell cycle/Division | dam damX dicC |
| Cell death | ldcA | |
| Genetic transfer | ompA ygcO | |
| Biofilm formation | csgF | |
| Adhesion | yraK | |
| Locomotion | fliL malE rzpD | |
| Viral response | ompA rzpD | |
| Bacterial response | rzpD | |
| Host interaction | ompA rzpD | |
| Response to stimulus | Heat | sodA |
| DNA damage | dam malE ompA recT yaiP yciT | |
| pH | sodA | |
| Oxidant detoxification | sodA | |
| Other | ahpF btuB crp cysC cysD cysH cysI dcd dppF envZ exbB fliL nhaB prc putA recT rzpD ybaM yejF yigL yojI |
| System | Subsystem | Gene 3 |
|---|---|---|
| Central Dogma | Transcription | arcB exuR fis galR glnL higB hupB rapA rfaH sspA rhoL ybeY yfjR |
| Translation | higB prfC rhaH rplI tufB ybeY | |
| DNA Metabolism | fis hsdS hofM ruvA mutL | |
| RNA Metabolism | arcB exuR fis galR glnL higB hupB rapA rfah rhoL rsmE rraB sspA ybeY yfjR ygfZ | |
| Protein Metabolism | arcB glnL higB hybD iadA mobA pflA prfC pqqL rfaH rplI tufB ybeY ygeY yicR | |
| Cell Exterior | Transport | chbB clcA cusB cysA cysP dtpB fepA feoB tdcC tolC trkH tyrP yiaN |
| Cell wall biogenesis/organization | amiB rfe | |
| Lipopolysaccharide metabolism | kdsD rfaD rfe waaG | |
| Pilus | yfcQ | |
| Flagellum | flgH | |
| Outer membrane | fepA flgH lpp tolC yraP | |
| Plasma membrane | arcB atpB atpE atpF bcsF clcA clcB cstA cysA dtpB feoB glnL glvB hokD hycB ppdB rfe sanA tdcC tolC trkH tufB tyrP ydcV ydjZ ygeY ygiZ yhaH yhjD yiaB yiaN yibN yjiG yqiJ | |
| Periplasm | amiB cusB cysP hmp lpp sanA tolC yfdX yjfY yraP ytfJ | |
| Cell wall components | rfe | |
| Biosynthesis | Amino acid biosynthesis | hisA ilvG lysC |
| Nucleotide biosynthesis | add | |
| Fatty acid and lipid biosynthesis | fabF wag clsB | |
| Carbohydrate biosynthesis | yggF rfaD kdsD | |
| Cofactor biosynthesis | mobA ubiE gshB | |
| Other | aroL lysC | |
| Degradation | Amino acid degradation | ilvG pflB |
| Nucleotide degradation | add | |
| Amine degradation | caiC | |
| Carbohydrate degradation | yidA ulaG | |
| Secondary metabolite degradation | lsrF | |
| Aromatic degradation | hcaD mhpC | |
| Other pathways | Other | amiB higB hmp hsdS iadA mutL nfsB nudF pflA qorB rsmE ruvA |
| Energy | Glycolysis | yggF |
| Pentose phosphate pathway | rpiA | |
| Fermentation | hycB pflB | |
| ATP synthesis | atpB atpE atpF | |
| Cellular processes | Cell cycle and division | amiB minC |
| Cell death | hokD | |
| Genetic transfer | ydcV | |
| Biofilm formation | yfjR | |
| Adhesion | yfcQ | |
| Locomotion | flgH | |
| Viral Response | fis | |
| Response to Stimulus | Starvation | cstA sanA sspA |
| Heat | Nudf ybeY yobF | |
| DNA damage | add feoB hisA mutL pflA ruvA ybiX yiaB yqiJ | |
| Osmotic stress | flgH | |
| pH | clcA | |
| Detoxification | cusB | |
| Other | arcB cstA dtpB fis glnL hcaD hmp hsdS mhpC sanA sspA tolC tufB yfdS yggX |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugala, N.; Lemire, J.; Chatfield-Reed, K.; Yan, Y.; Chua, G.; Turner, R.J. Using a Chemical Genetic Screen to Enhance Our Understanding of the Antibacterial Properties of Silver. Genes 2018, 9, 344. https://doi.org/10.3390/genes9070344
Gugala N, Lemire J, Chatfield-Reed K, Yan Y, Chua G, Turner RJ. Using a Chemical Genetic Screen to Enhance Our Understanding of the Antibacterial Properties of Silver. Genes. 2018; 9(7):344. https://doi.org/10.3390/genes9070344
Chicago/Turabian StyleGugala, Natalie, Joe Lemire, Kate Chatfield-Reed, Ying Yan, Gordon Chua, and Raymond J. Turner. 2018. "Using a Chemical Genetic Screen to Enhance Our Understanding of the Antibacterial Properties of Silver" Genes 9, no. 7: 344. https://doi.org/10.3390/genes9070344
APA StyleGugala, N., Lemire, J., Chatfield-Reed, K., Yan, Y., Chua, G., & Turner, R. J. (2018). Using a Chemical Genetic Screen to Enhance Our Understanding of the Antibacterial Properties of Silver. Genes, 9(7), 344. https://doi.org/10.3390/genes9070344