Size and Content of the Sex-Determining Region of the Y Chromosome in Dioecious Mercurialis annua, a Plant with Homomorphic Sex Chromosomes

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
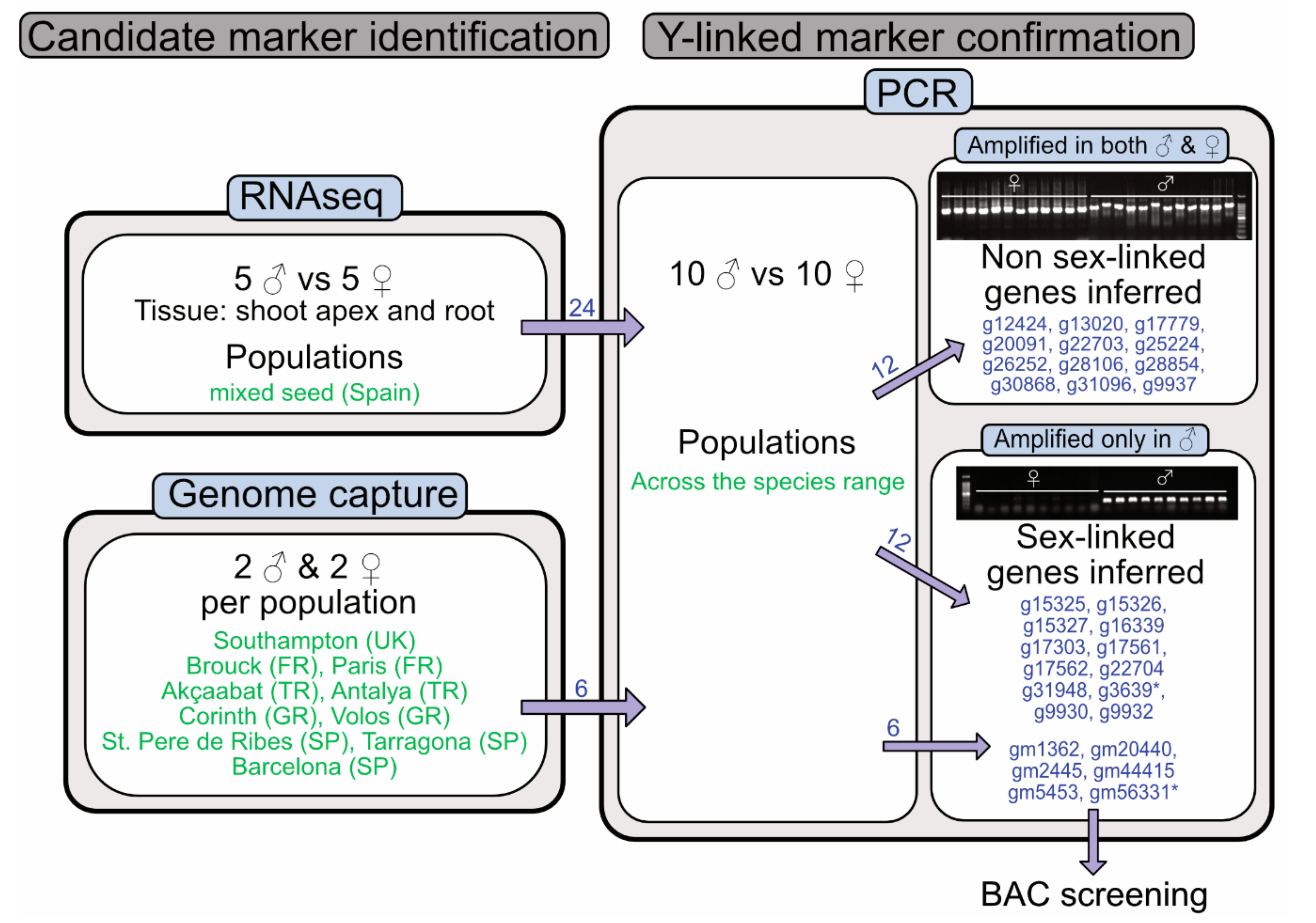
2.1. Overview
2.2. Identification and Confirmation of Y-Linked Markers
2.3. Bacterial Artifical Chromosome Library Construction, Sequencing and Assembly
2.4. Transcript and Transposable Element Annotation
3. Results
3.1. Identification of Male-Specific Genes Based on Expression and Genome Capture Data
3.2. Bacterial Artifical Chromosome Assembly
3.3. Functional Annotation of Genes Located on the Bacterial Artifical Chromosomes
3.4. Comparison of Transposable Element Density and Type Between the Bactertial Artificial Chromosomes and the Full Genome
4. Discussion
4.1. Identification of Y-Linked Markers
4.2. Size of the Sex-Determining Region
4.3. Content of the Sex-Determining Region
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Renner, S.S. The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 2014, 101, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlates in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef]
- Beukeboom, L.W.; Perrin, N. The Evolution of Sex Determination; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Policansky, D. Sex change in plants and animals. Annu. Rev. Ecol. Syst. 1982, 13, 471–495. [Google Scholar] [CrossRef]
- Zimmerman, J.K. Ecological correlates of labile sex expression in the orchid Catasetum viridiflavum. Ecology 1991, 72, 597–608. [Google Scholar] [CrossRef]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex chromosomes in land plants. Annu. Rev. Plant Boil. 2011, 62, 485–514. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D. Plant sex chromosomes. Annu. Rev. Plant Biol. 2016, 67, 397–420. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Fuchs, J.; Renner, S.S. Cytogenetic comparison of heteromorphic and homomorphic sex chromosomes in Coccinia (Cucurbitaceae) points to sex chromosome turnover. Chromosome Res. 2017, 25, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D. Plant contributions to our understanding of sex chromosome evolution. New Phytol. 2015, 208, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Fuchs, J.; Renner, S.S. Molecular cytogenetics (FISH, GISH) of Coccinia grandis: A ca. 3 myr-old species of Cucurbitaceae with the largest Y/autosome divergence in flowering plants. Cytogenet. Genome Res. 2013, 139, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Cherif, E.; Zehdi-Azouzi, S.; Crabos, A.; Castillo, K.; Chabrillange, N.; Pintaud, J.C.; Salhi-Hannachi, A.; Glemin, S.; Aberlenc-Bertossi, F. Evolution of sex chromosomes prior to speciation in the dioecious Phoenix species. J. Evol. Biol. 2016, 29, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- AbdAlla, M.M.; Abd El-Kawy, M. Cytological studies for date palm (Phoenix dactylifera L.) tissue culture derived plants. Rep. Opin. 2010, 2, 17–21. [Google Scholar]
- Hagelamin, O.; Alzahrani, M.M.A. Characterization and analysis of date palm (Phoenix dactylifera L.) karyotypes. Asia Life Sci. 2015, 24, 809–822. [Google Scholar]
- Bergero, R.; Charlesworth, D. The evolution of restricted recombination in sex chromosomes. Trends Ecol. Evol. 2009, 24, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Hobza, R.; Cegan, R.; Jesionek, W.; Kejnovsky, E.; Vyskot, B.; Kubat, Z. Impact of repetitive elements on the Y chromosome formation in plants. Genes 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Bergero, R.; Guirao-Rico, S.; Campos, J.L.; Cezard, T.; Gharbi, K.; Charlesworth, D. RAD mapping reveals an evolving, polymorphic and fuzzy boundary of a plant pseudoautosomal region. Mol. Ecol. 2016, 25, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.G.; Tsang, G.K.; Datson, P.M.; De Silva, H.N.; Harvey, C.F.; Gill, G.P.; Crowhurst, R.N.; McNeilage, M.A. A gene-rich linkage map in the dioecious species Actinidia chinensis (kiwifruit) reveals putative X/Y sex-determining chromosomes. BMC Genom. 2009, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.S.; Spannagl, M.; Al-Malki, A.; George, B.; Torres, M.F.; Al-Dous, E.K.; Al-Azwani, E.K.; Hussein, E.; Mathew, S.; Mayer, K.F.X.; et al. A first genetic map of date palm (Phoenix dactylifera) reveals long-range genome structure conservation in the palms. BMC Genom. 2014, 15, 285. [Google Scholar] [CrossRef] [PubMed]
- Hobza, R.; Lengerova, M.; Svoboda, J.; Kubekova, H.; Kejnovsky, E.; Vyskot, B. An accumulation of tandem DNA repeats on the Y chromosome in Silene latifolia during early stages of sex chromosome evolution. Chromosoma 2006, 115, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Na, J.K.; Wang, J.P.; Murray, J.E.; Gschwend, A.R.; Zhang, W.L.; Yu, Q.Y.; Navajas-Perez, R.; Feltus, F.A.; Chen, C.X.; Kubat, Z.; et al. Construction of physical maps for the sex-specific regions of papaya sex chromosomes. BMC Genom. 2012, 13, 176. [Google Scholar] [CrossRef] [PubMed]
- Telgmann-Rauber, A.; Jamsari, A.; Kinney, M.S.; Pires, J.C.; Jung, C. Genetic and physical maps around the sex-determining M-locus of the dioecious plant Asparagus. Mol. Genet. Genom. 2007, 278, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Pannell, J.R.; Dorken, M.E.; Pujol, B.; Berjano, R. Gender variation and transitions between sexual systems in Mercurialis annua (Euphorbiaceae). Int. J. Plant Sci. 2008, 169, 129–139. [Google Scholar] [CrossRef]
- Russell, J.R.W.; Pannell, J.R. Sex determination in dioecious Mercurialis annua and its close diploid and polyploid relatives. Heredity 2015, 114, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Durand, B. Le complèxe Mercurialis annua L. s.l.: Une étude biosystématique. Ann. Sci. Nat. Bot. Paris 1963, 12, 579–736. [Google Scholar]
- Durand, R.; Durand, B. Dioecy, monoecy, polyploidy and speciation in the annual Mercuries. Bull. Soc. Bot. France Lett. Bot. 1992, 139, 377–399. [Google Scholar]
- Obbard, D.J.; Harris, S.A.; Pannell, J. Sexual systems and population genetic structure in an annual plant: Testing the metapopulation model. Am. Nat. 2006, 167, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Eppley, S.M.; Pannell, J.R. Sexual systems and measures of occupancy and abundance in an annual plant: Testing the metapopulation model. Am. Nat. 2007, 169, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Pannell, J.R.; Eppley, S.M.; Dorken, M.E.; Berjano, R. Regional variation in sex ratios and sex allocation in androdioecious Mercurialis annua. J. Evol. Biol. 2014, 27, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.; Veltsos, P.; Muyle, A.; Emery, O.; Rastas, P.; Marais, G.; Filatov, D.; Pannell, J.R. Hallmarks of early sex-chromosome evolution in the dioecious plant Mercurialis annua revealed by de novo genome assembly, genetic mapping and transcriptome analysis. bioRxiv 2017. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, S.C.; Ridout, K.; Pannell, J.R. Range expansion compromises adaptive evolution in an outcrossing plant. Curr. Biol. 2017, 27, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Cossard, G.; Pannell, J.R. Sexual dimorphism and rapid turnover in gene expression in pre-reproductive seedlings of a dioecious herb. 2018, submitted.
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.F.A.; Hubley, R.; Green, P. RepeatMasker Open-4.0. Available online: http://www.repeatmasker.org/.
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Schoffmann, O.; Morgenstern, B.; Waack, S. Gene prediction in eukaryotes with a generalized hidden Markov model that uses hints from external sources. BMC Bioinform. 2006, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Adamczyk, B.J.; Fernandez, D.E. MIKC* MADS Domain Heterodimers Are Required for Pollen Maturation and Tube Growth in Arabidopsis. Plant Physiol. 2009, 149, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.N.; Zambryski, P.C. ORGAN BOUNDARY1 defines a gene expressed at the junction between the shoot apical meristem and lateral organs. Proc. Natl. Acad. Sci. USA 2011, 108, 2154–2159. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Matsushika, A.; Ashikari, M.; Yamashino, T.; Mizuno, T. Circadian-associated rice pseudo response regulators (OsPRRs): Insight into the control of flowering time. Biosci. Biotechnol. Biochem. 2005, 69, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Cherif, E.; Zehdi, S.; Castillo, K.; Chabrillange, N.; Abdoulkader, S.; Pintaud, J.C.; Santoni, S.; Salhi-Hannachi, A.; Glémin, S.; Aberlenc-Bertossi, F. Male-specific DNA markers provide genetic evidence of an XY chromosome system, a recombination arrest and allow the tracing of paternal lineages in date palm. New Phytol. 2013, 197, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Muyle, A.; Zemp, N.; Deschamps, C.; Mousset, S.; Widmer, A.; Marais, G.A.B. Rapid de novo evolution of X chromosome dosage compensation in Silene latifolia, a plant with young sex chromosomes. PLoS Biol. 2012, 10, e1001308. [Google Scholar] [CrossRef] [PubMed]
- Khadka, D.K.; Nejidat, A.; Tal, M.; Golan-Goldhirsh, A. DNA markers for sex: Molecular evidence for gender dimorphism in dioecious Mercurialis annua L. Mol. Breed. 2002, 9, 251–257. [Google Scholar] [CrossRef]
- Fechter, I.; Hausmann, L.; Daum, M.; Sorensen, T.R.; Viehover, P.; Weisshaar, B.; Topfer, R. Candidate genes within a 143 kb region of the flower sex locus in Vitis. Mol. Genet. Genom. 2012, 287, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Picq, S.; Santoni, S.; Lacombe, T.; Latreille, M.; Weber, A.; Ardisson, M.; Ivorra, S.; Maghradze, D.; Arroyo-Garcia, R.; Chatelet, P.; et al. A small XY chromosomal region explains sex determination in wild dioecious V. vinifera and the reversal to hermaphroditism in domesticated grapevines. BMC Plant Biol. 2014, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, I.; Gaudet, M.; Jorge, V.; Beritognolo, I.; Terzoli, S.; Kuzminsky, E.; Muleo, R.; Mugnozza, G.S.; Sabatti, M. Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species. Tree Genet. Genomes 2010, 6, 863–875. [Google Scholar] [CrossRef]
- Geraldes, A.; Hefer, C.A.; Capron, A.; Kolosova, N.; Martinez-Nunez, F.; Soolanayakanahally, R.Y.; Stanton, B.; Guy, R.D.; Mansfield, S.D.; Douglas, C.J.; et al. Recent Y chromosome divergence despite ancient origin of dioecy in poplars (Populus). Mol. Ecol. 2015, 24, 3243–3256. [Google Scholar] [CrossRef] [PubMed]
- Tennessen, J.A.; Govindarajulu, R.; Liston, A.; Ashman, T.L. Homomorphic ZW chromosomes in a wild strawberry show distinctive recombination heterogeneity but a small sex-determining region. New Phytol. 2016, 211, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, E. Selbstbestäubungen subdiöcischer Blütenpflanzen, ein neuer Beweis für die genetische Theorie der Geschlechtsbestimmung. Planta 1939, 30, 457–470. [Google Scholar] [CrossRef]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Na, J.K.; Yu, Q.Y.; Gschwend, A.R.; Han, J.; Zeng, F.C.; Aryal, R.; VanBuren, R.; Murray, J.E.; Zhang, W.L.; et al. Sequencing papaya X and Y-h chromosomes reveals molecular basis of incipient sex chromosome evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13710–13715. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Perrin, N. Sex reversal: A fountain of youth for sex chromosomes? Evolution 2009, 63, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Akiyama, Y.; Fukui, K.; Kamada, H.; Satoh, S. Characterization, genome sizes and morphology of sex chromosomes in hemp (Cannabis sativa L.). Cytologia 1998, 63, 459–464. [Google Scholar] [CrossRef]
- Van Bakel, H.; Stout, J.M.; Cote, A.G.; Tallon, C.M.; Sharpe, A.G.; Hughes, T.R.; Page, J.E. The draft genome and transcriptome of Cannabis sativa. Genome Biol. 2011, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Grabowska-Joachimiak, A.; Sliwinska, E.; Pigula, M.; Skomra, U.; Joachimiak, A.J. Genome size in Humulus lupulus L. and H. japonicus Siebold & Zucc. (Cannabaceae). Acta Soc. Bot. Pol. 2006, 75, 207–214. [Google Scholar]
- Divashuk, M.G.; Alexandrov, O.S.; Kroupin, P.Y.; Karlov, G.I. Molecular cytogenetic mapping of Humulus lupulus sex chromosomes. Cytogenet. Genome Res. 2011, 134, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.J.; Filatov, D.A. A cytogenetic view of sex chromosome evolution in plants. Cytogenet. Genome Res. 2008, 120, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Błocka-Wandas, M.; Sliwinska, E.; Grabowska-Joachimiak, A.; Musial, K.; Joachimiak, A.J. Male gametophyte development and two different DNA classes of pollen grains in Rumex acetosa L., a plant with an XX/XY1Y2 sex chromosome system and a female-biased sex ratio. Sex. Plant Reprod. 2007, 20, 171–180. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Plant DNA C-values Database. Available online: http://data.kew.org/cvalues/ (accessed on 16 April 2018).
- Shibata, F.; Hizume, M.; Kuroki, Y. Differentiation and the polymorphic nature of the Y chromosomes revealed by repetitive sequences in the dioecious plant, Rumex acetosa. Chromosome Res. 2000, 8, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Hough, J.; Hollister, J.D.; Wang, W.; Barrett, S.C.H.; Wright, S.I. Genetic degeneration of old and young Y chromosomes in the flowering plant Rumex hastatulus. Proc. Natl. Acad. Sci. USA 2014, 111, 7713–7718. [Google Scholar] [CrossRef] [PubMed]
- Grabowska-Joachimiak, A.; Kula, A.; Ksiazczyk, T.; Chojnicka, J.; Sliwinska, E.; Joachimiak, A.J. Chromosome landmarks and autosome-sex chromosome translocations in Rumex hastatulus, a plant with XX/XY1Y2 sex chromosome system. Chromosome Res. 2015, 23, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.X.; Ding, J.; Deng, D.J.; Tang, W.; Sun, H.H.; Liu, D.Y.; Zhang, L.; Niu, X.L.; Zhang, X.; Meng, M.; et al. Draft genome of the kiwifruit Actinidia chinensis. Nat. Commun. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, C.Y.; Liu, Y.F.; VanBuren, R.; Yao, X.H.; Zhong, C.H.; Huang, H.W. High-density interspecific genetic maps of kiwifruit and the identification of sex-specific markers. DNA Res. 2015, 22, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Al-Dous, E.K.; George, B.; Al-Mahmoud, M.E.; Al-Jaber, M.Y.; Wang, H.; Salameh, Y.M.; Al-Azwani, E.K.; Chaluvadi, S.; Pontaroli, A.C.; DeBarry, J.; et al. De novo genome sequencing and comparative genomics of date palm (Phoenix dactylifera). Nat. Biotechnol. 2011, 29, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Al-Mssallem, I.S.; Hu, S.; Zhang, X.; Lin, Q.; Liu, W.; Tan, J.; Yu, X.; Liu, J.; Pan, L.; Zhang, T.; et al. Genome sequence of the date palm Phoenix dactylifera L. Nat. Commun. 2013, 4, 2274. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, S.; Khodaeiaminjan, M.; Guney, M.; Kafkas, E. Identification of sex-linked SNP markers using RAD sequencing suggests ZW/ZZ sex determination in Pistacia vera L. BMC Genom. 2015, 16, 98. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Hou, S.B.; Feng, Y.; Yu, Q.Y.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991. [Google Scholar] [CrossRef] [PubMed]
- Spigler, R.B.; Lewers, K.S.; Main, D.S.; Ashman, T.L. Genetic mapping of sex determination in a wild strawberry, Fragaria virginiana, reveals earliest form of sex chromosome. Heredity 2008, 101, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Tao, R.; Yonemori, K.; Utsunomiya, N.; Sugiura, A. Ploidy level and genome size of several Diospyros species. J. Jpn. Soc. Hortic. Sci. 1998, 67, 306–312. [Google Scholar] [CrossRef]
- Akagi, T.; Henry, I.M.; Tao, R.; Comai, L. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 2014, 346, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; DiFazio, S.P.; Gunter, L.E.; Zhang, X.; Sewell, M.M.; Woolbright, S.A.; Allan, G.J.; Kelleher, C.T.; Douglas, C.J.; Wang, M.; et al. Genome structure and emerging evidence of an incipient sex chromosome in Populus. Genome Res. 2008, 18, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Pakull, B.; Groppe, K.; Meyer, M.; Markussen, T.; Fladung, M. Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.). Tree Genet. Genomes 2009, 5, 505–515. [Google Scholar] [CrossRef]
- Pucholt, P.; Ronnberg-Wastljung, A.C.; Berlin, S. Single locus sex determination and female heterogamety in the basket willow (Salix viminalis L.). Heredity 2015, 114, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463. [Google Scholar] [PubMed]
- Obbard, D.J.; Harris, S.A.; Buggs, R.J.A.; Pannell, J.R. Hybridization, polyploidy, and the evolution of sexual systems in Mercurialis (Euphorbiaceae). Evolution 2006, 60, 1801–1815. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.F.; Skaletsky, H.; Pyntikova, T.; Graves, T.A.; van Daalen, S.K.M.; Minx, P.J.; Fulton, R.S.; McGrath, S.D.; Locke, D.P.; Friedman, C.; et al. Chimpanzee and human Y chromosomes are remarkably divergent in structure and gene content. Nature 2010, 463, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Evolution of the avian sex chromosomes and their role in sex determination. Trends Ecol. Evol. 2000, 15, 188–192. [Google Scholar] [CrossRef]
- Ayers, K.L.; Davidson, N.M.; Demiyah, D.; Roeszler, K.N.; Grutzner, F.; Sinclair, A.H.; Oshlack, A.; Smith, C.A. RNA sequencing reveals sexually dimorphic gene expression before gonadal differentiation in chicken and allows comprehensive annotation of the W-chromosome. Genome Biol. 2013, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, A.; Murata, K.; Mizuno, S. The location of Z- and W-linked marker genes and sequence on the homomorphic sex chromosomes of the ostrich and the emu. Proc. Natl. Acad. Sci. USA 1998, 95, 4415–4418. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, J.L.; Bachtrog, D.; An, N.; Huang, Q.F.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G.J. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1246338. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.C.; Grabherr, M.G.; Chan, Y.F.; Russell, P.; Mauceli, E.; Johnson, J.; Swofford, R.; Pirun, M.; Zody, M.C.; White, S.; et al. The genomic basis of adaptive evolution in threespine sticklebacks. Nature 2012, 484, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Naruse, K.; Sasaki, S.; Nakatani, Y.; Qu, W.; Ahsan, B.; Yamada, T.; Nagayasu, Y.; Doi, K.; Kasai, Y.; et al. The medaka draft genome and insights into vertebrate genome evolution. Nature 2007, 447, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhu, H.M.; Huang, Q.F.; Zhao, L.; Zhang, G.J.; Roy, S.W.; Vicoso, B.; Xuan, Z.L.; Ruan, J.; Zhang, Y.; et al. Deciphering neo-sex and B chromosome evolution by the draft genome of Drosophila albomicans. BMC Genom. 2012, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Liu, Y.; Bettencourt, B.R.; Hradecky, P.; Letovsky, S.; Nielsen, R.; Thornton, K.; Hubisz, M.J.; Chen, R.; Meisel, R.P.; et al. Comparative genome sequencing of Drosophila pseudoobscura: Chromosomal, gene, and cis-element evolution. Genome Res. 2005, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Steinemann, M.; Steinemann, S. Enigma of Y chromosome degeneration: Neo-Y and Neo-X chromosomes of Drosophila miranda a model for sex chromosome evolution. Genetica 1998, 102, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, V.B.; Charlesworth, B. Muller’s ratchet and the degeneration of the Drosophila miranda neo-Y chromosome. Genetics 2010, 185, 339. [Google Scholar] [CrossRef] [PubMed]
- Criscione, C.D.; Valentim, C.L.L.; Hirai, H.; LoVerde, P.T.; Anderson, T.J.C. Genomic linkage map of the human blood fluke Schistosoma mansoni. Genome Biol. 2009, 10, R71. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| BAC Contig | Length (Kb) | Male-Specific PCR Hit | ORFs | Additional Predicted Genes | Potential Candidate Genes for Sex Determination |
|---|---|---|---|---|---|
| 1, 2, 9 | 170 | g16339 | 3 | 3 | |
| 3 | 110 | g17561 | 1 | 3 | |
| 4, 21 | 143 | g22704 | 4 | 0 | |
| 5, 20 | 200 | g9930, g9932 | 3 | 0 | |
| 6 | 103 | g31948 | 3 | 1 | g18/g22596: XP_021600590.1 Agamous-like MADS-box protein AGL66 |
| 8 | 146 | NA | 13 | 3 | g21/g20779: XP_002519813.1 Auxin response factor |
| 10 | 127 | g2445 | 1 | 0 | |
| 11 | 209 | g5453 | 2 | 0 | |
| 12 | 123 | gm20440 | 2 | 2 | g41/g29576: XP_002267312.1 Light-dependent short hypocotyls 6 |
| 13 | 99 | g3639 | 2 | 0 | g44/g27235: XP_021670876.1 Two-component response regulator-like PRR73 |
| 7, 14, 23, 24 | 215 | g15326, g15327 | 5 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veltsos, P.; Cossard, G.; Beaudoing, E.; Beydon, G.; Savova Bianchi, D.; Roux, C.; C. González-Martínez, S.; R. Pannell, J. Size and Content of the Sex-Determining Region of the Y Chromosome in Dioecious Mercurialis annua, a Plant with Homomorphic Sex Chromosomes. Genes 2018, 9, 277. https://doi.org/10.3390/genes9060277
Veltsos P, Cossard G, Beaudoing E, Beydon G, Savova Bianchi D, Roux C, C. González-Martínez S, R. Pannell J. Size and Content of the Sex-Determining Region of the Y Chromosome in Dioecious Mercurialis annua, a Plant with Homomorphic Sex Chromosomes. Genes. 2018; 9(6):277. https://doi.org/10.3390/genes9060277
Chicago/Turabian StyleVeltsos, Paris, Guillaume Cossard, Emmanuel Beaudoing, Genséric Beydon, Dessislava Savova Bianchi, Camille Roux, Santiago C. González-Martínez, and John R. Pannell. 2018. "Size and Content of the Sex-Determining Region of the Y Chromosome in Dioecious Mercurialis annua, a Plant with Homomorphic Sex Chromosomes" Genes 9, no. 6: 277. https://doi.org/10.3390/genes9060277
APA StyleVeltsos, P., Cossard, G., Beaudoing, E., Beydon, G., Savova Bianchi, D., Roux, C., C. González-Martínez, S., & R. Pannell, J. (2018). Size and Content of the Sex-Determining Region of the Y Chromosome in Dioecious Mercurialis annua, a Plant with Homomorphic Sex Chromosomes. Genes, 9(6), 277. https://doi.org/10.3390/genes9060277