A Toxin-Antitoxin System VapBC15 from Synechocystis sp. PCC 6803 Shows Distinct Regulatory Features

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Culture Conditions, and Biochemicals
2.2. Plasmid Construction
2.3. Analyses of Toxicity, Anti-Toxicity, and Growth Rescue
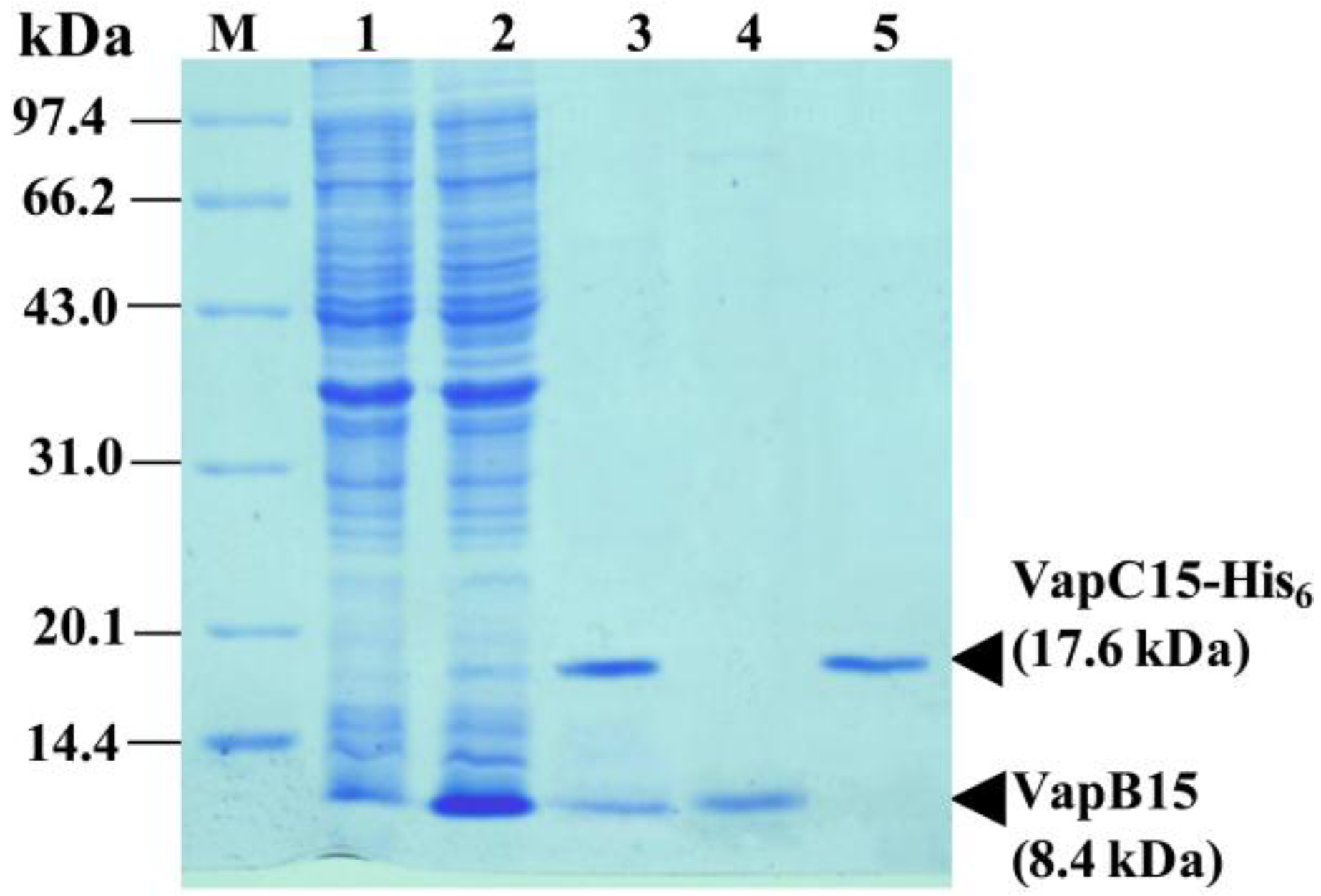
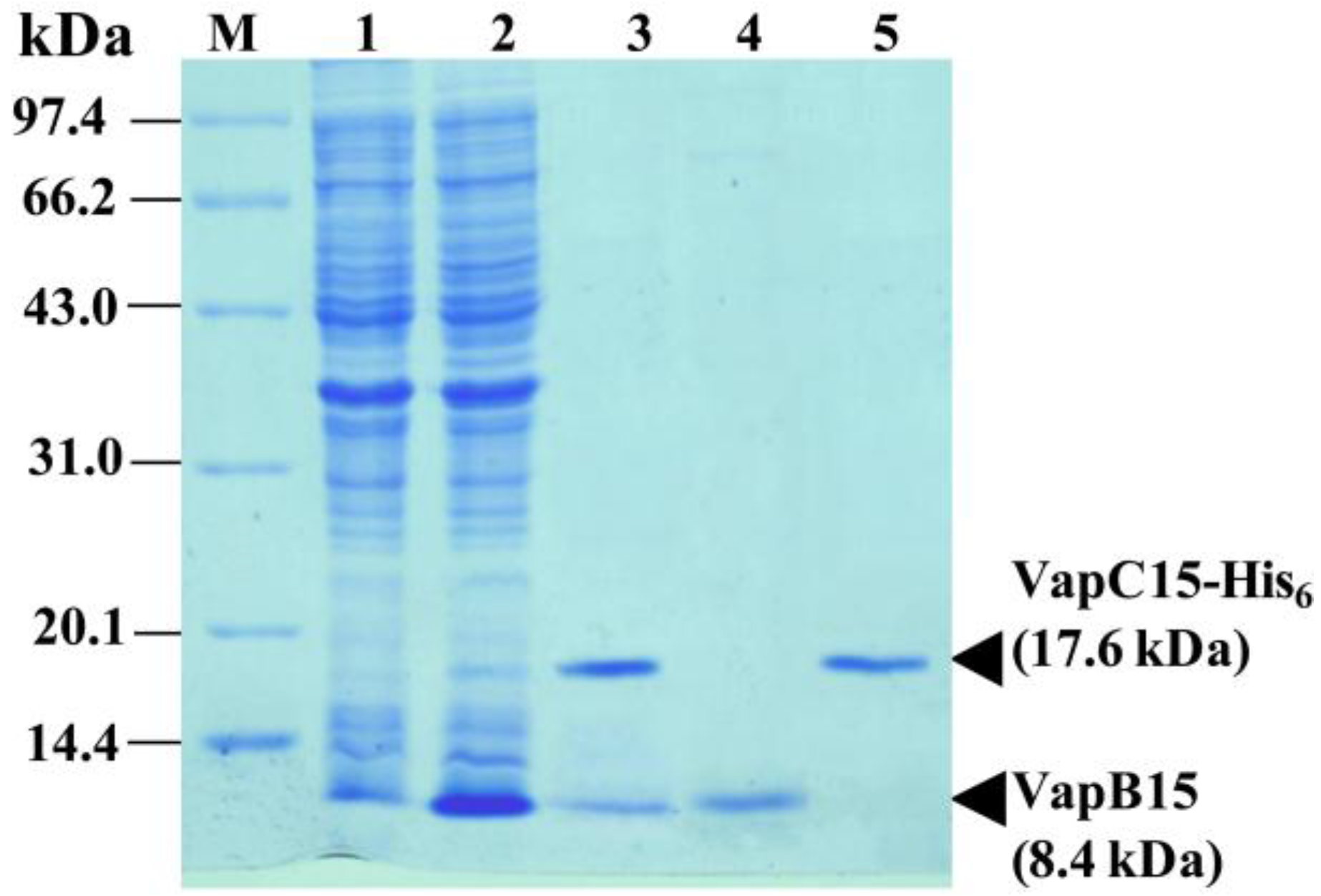
2.4. Co-Expression, Purification and Antibody Preparation of VapB15 and VapC15
2.5. Assay of β-Galactosidase (LacZ) Activity
2.6. Electrophoretic Mobility Shift Assay (EMSA)
2.7. Western Blot Analysis
3. Results
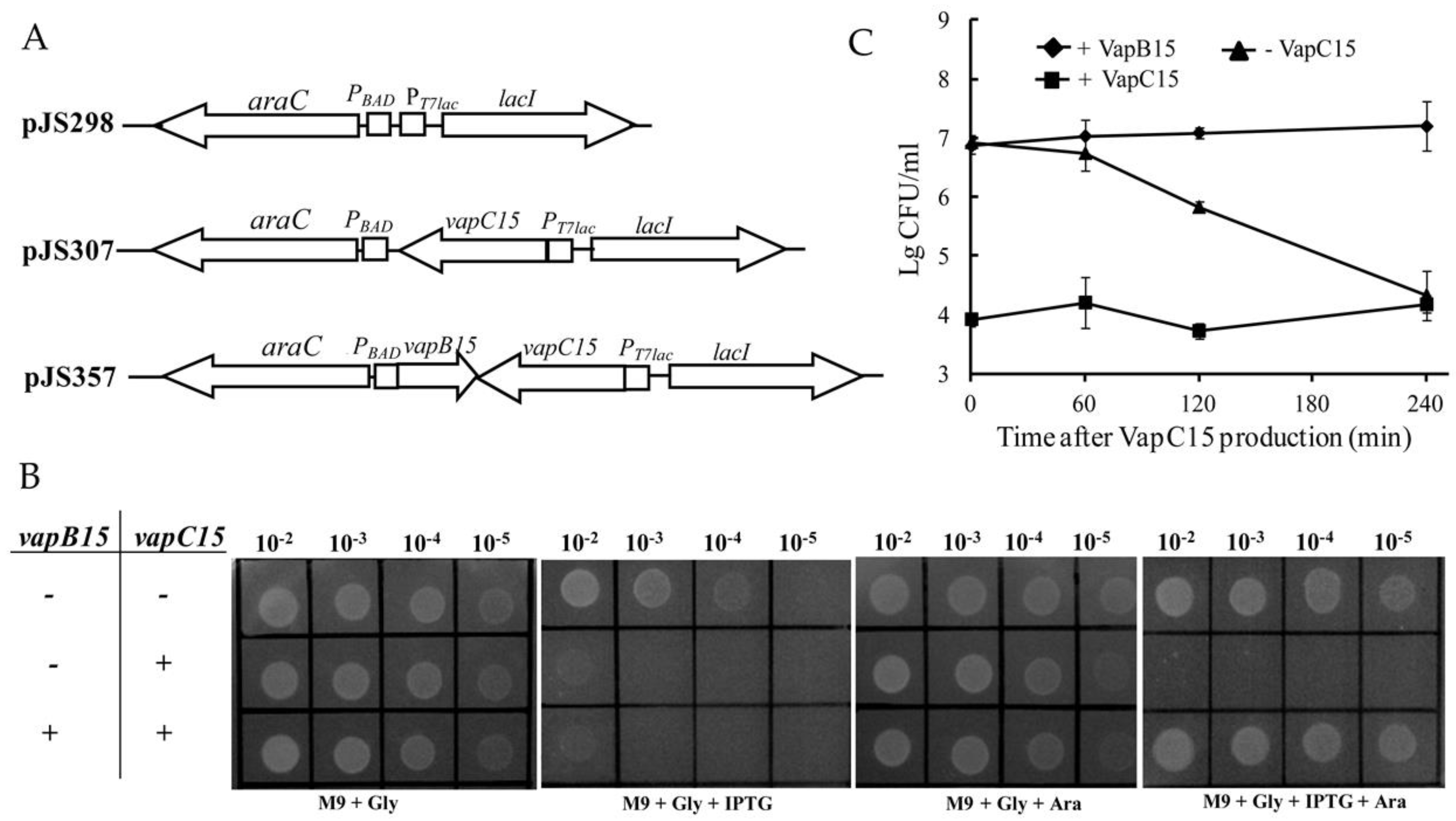
3.1. Growth-Arrest Effect of VapC15 and Toxicity-Neutralization Effect of VapB15
3.2. VapB15 Rescues the Growth Arrest Induced by VapC15
3.3. Physical Interaction between VapB15 and VapC15
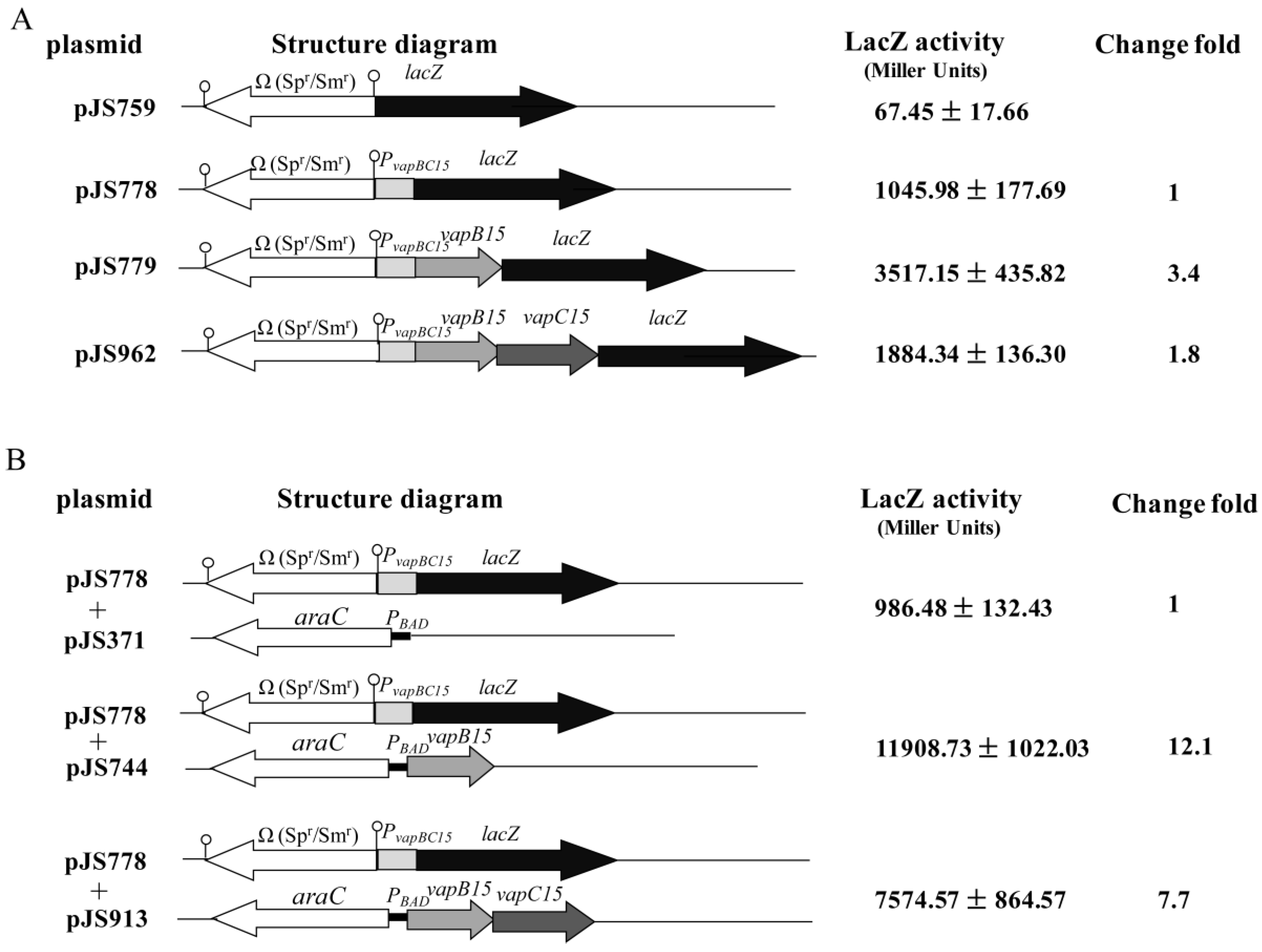
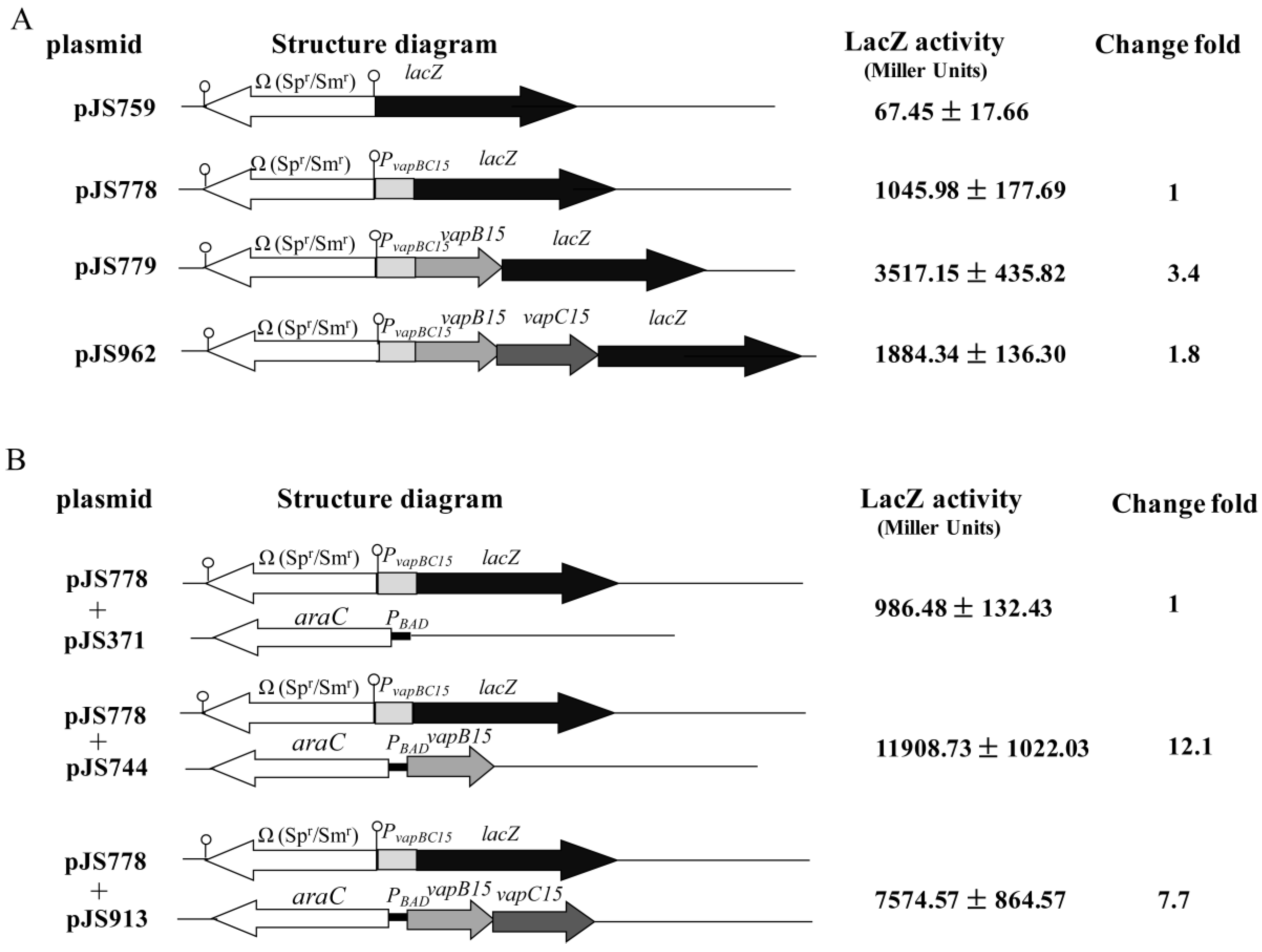
3.4. Up-Regulation Effect of VapB15 and Neutralization Effect of VapC15
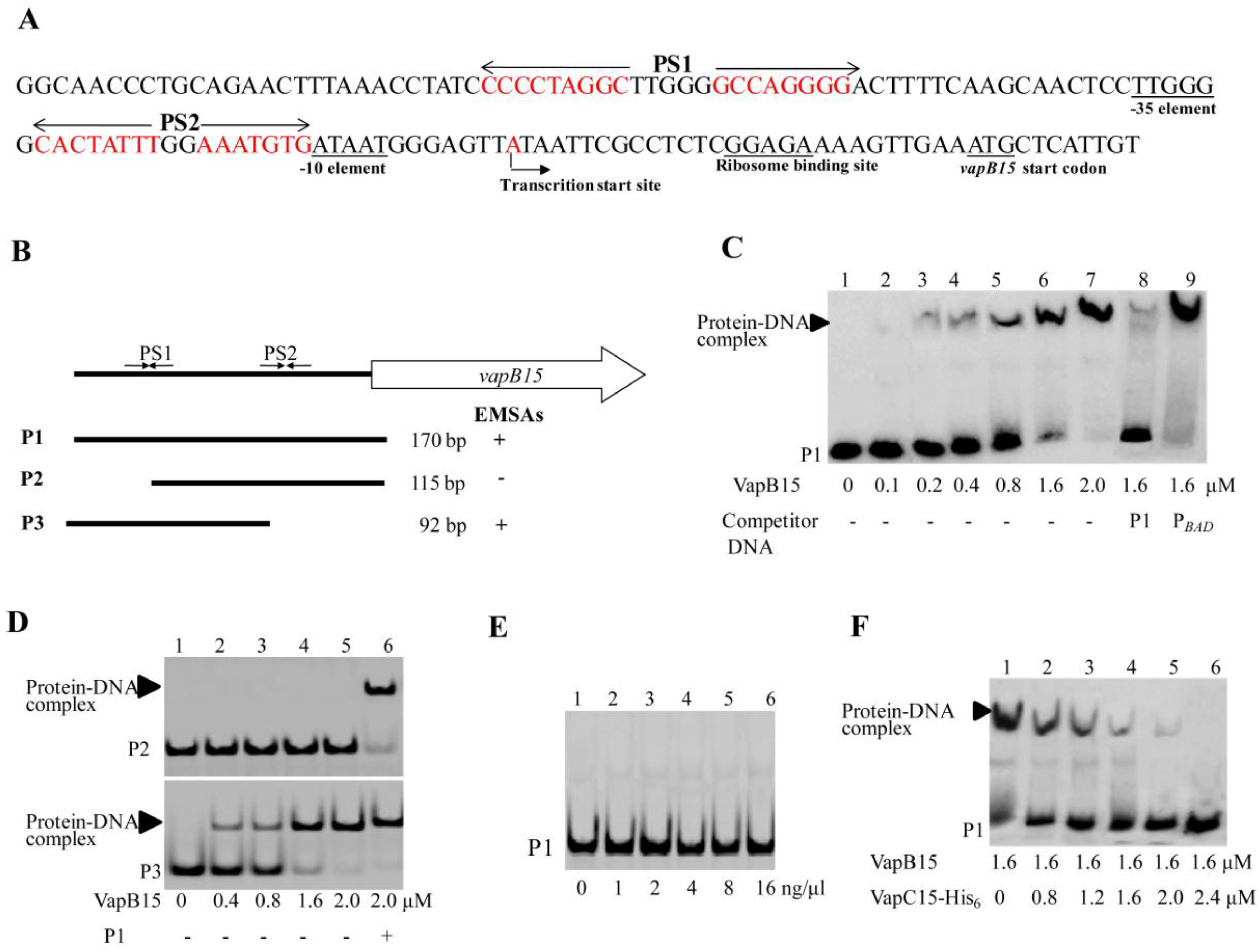
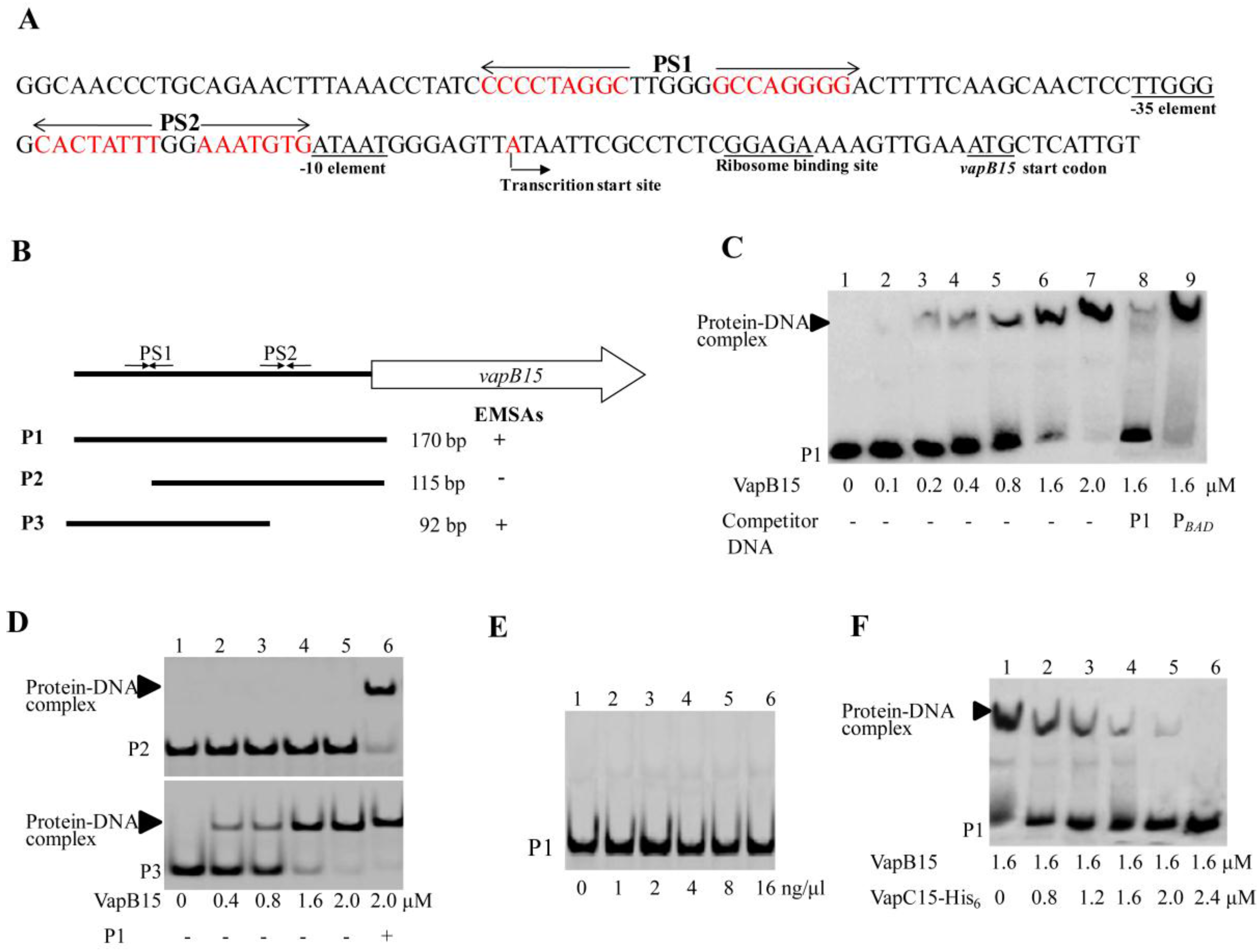
3.5. VapB15 Specifically Binds to the PvapBC15 Promoter
3.6. VapC15 Destabilizes the Binding of VapB15 to the PvapBC15 Promoter
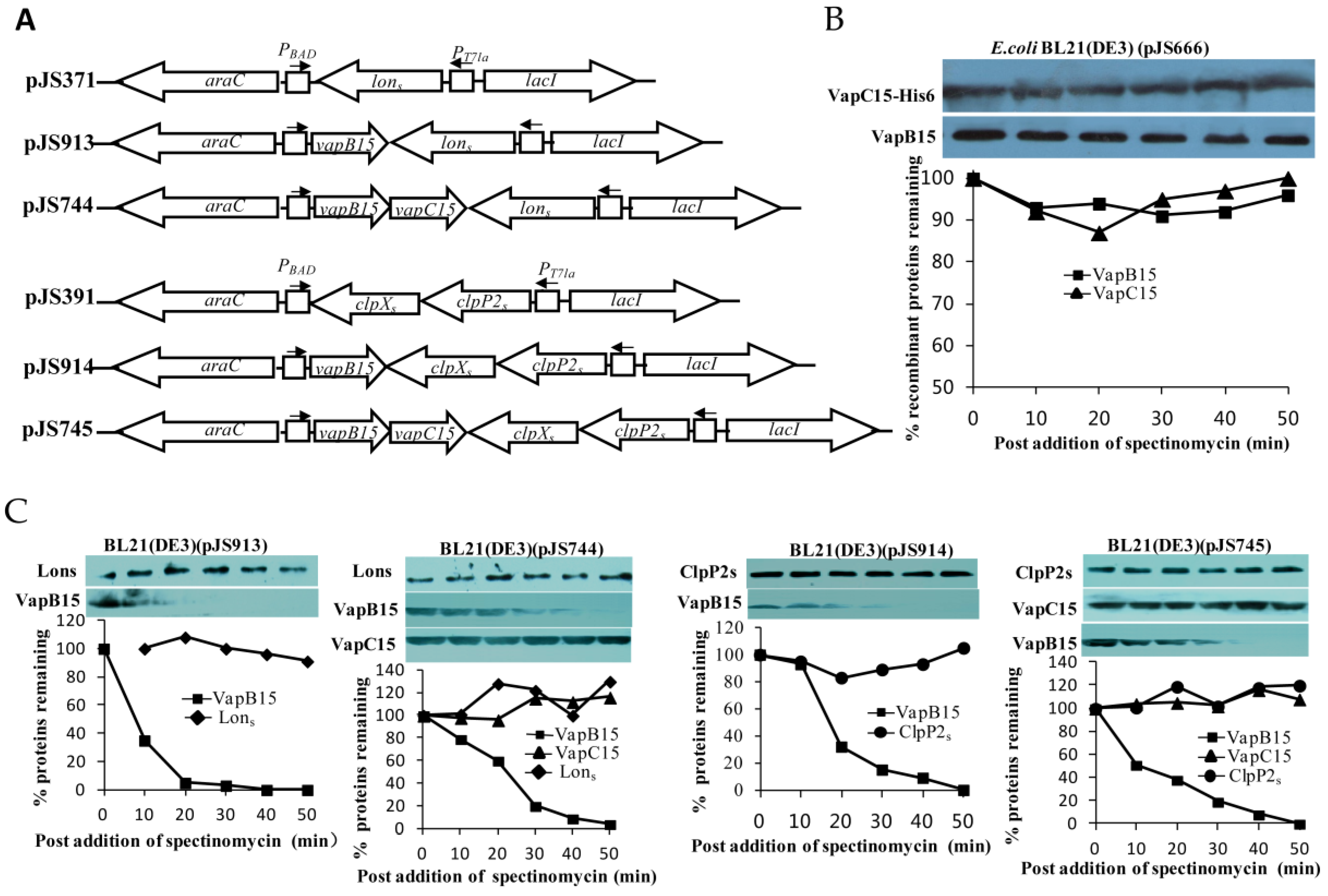
3.7. VapB15 Is Susceptble to the Proteases ClpXP2s and Lons
3.8. Proteolysis of VapB15 Activates the Potential Toxicity of VapBC15
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ogura, T.; Hiraga, S. Mini-F plasmid genes that couple host cell division to plasmid proliferation. Proc. Natl. Acad. Sci. USA 1983, 80, 4784–4788. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.Y.; Rajakumar, K.; Deng, Z. TADB: A web-based resource for Type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic Acids Res. 2011, 39, D606–D611. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.; Wolf, Y.; Koonin, E. Comprehensive comparative-genomic analysis of Type 2 toxin-antitoxin systems and related mobile stress response systems in prokaryotes. Biol. Direct 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, D.; Natarajan, J. Network analysis of S. aureus response to ramoplanin reveals modules for virulence factors and resistance mechanisms and characteristic novel genes. Gene 2015, 574, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Van Melderen, L.; Saavedra De Bast, M. Bacterial toxin-antitoxin systems: More than selfish entities? PLoS Genet. 2009, 5, e1000437. [Google Scholar] [CrossRef] [PubMed]
- Coussens, N.P.; Daines, D.A. Wake me when it’s over-Bacterial toxin-antitoxin proteins and induced dormancy. Exp. Biol. Med. 2016, 241, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Blower, T.R.; Short, F.L.; Rao, F.; Mizuguchi, K.; Pei, X.Y.; Fineran, P.C.; Luisi, B.F.; Salmond, G.P. Identification and classification of bacterial Type III toxin-antitoxin systems encoded in chromosomal and plasmid genomes. Nucleic Acids Res. 2012, 40, 6158–6173. [Google Scholar] [CrossRef] [PubMed]
- Fozo, E.M.; Makarova, K.S.; Shabalina, S.A.; Yutin, N.; Koonin, E.V.; Storz, G. Abundance of Type I toxin-antitoxin systems in bacteria: Searches for new candidates and discovery of novel families. Nucleic Acids Res. 2010, 38, 3743–3759. [Google Scholar] [CrossRef] [PubMed]
- Brantl, S.; Jahn, N. sRNAs in bacterial type I and type III toxin-antitoxin systems. FEMS Microbiol. Rev. 2015, 39, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Lee, B.J. Structure, biology, and therapeutic application of toxin-antitoxin systems in pathogenic bacteria. Toxins 2016, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Schuster, C.F.; Bertram, R. Toxin-antitoxin systems are ubiquitous and versatile modulators of prokaryotic cell fate. FEMS Microbiol. Lett. 2013, 340, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Aakre, C.D.; Phung, T.N.; Huang, D.; Laub, M.T. A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp. Mol. Cell 2013, 52, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Lord, D.M.; Cheng, H.Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Christensen, S.K.; Lobner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Inouye, M. Regulation of growth and death in Escherichia coli by toxin-antitoxin systems. Nat. Rev. Microbiol. 2011, 9, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Leplae, R.; Geeraerts, D.; Hallez, R.; Guglielmini, J.; Dreze, P.; Van Melderen, L. Diversity of bacterial type II toxin-antitoxin systems: A comprehensive search and functional analysis of novel families. Nucleic Acids Res. 2011, 39, 5513–5525. [Google Scholar] [CrossRef] [PubMed]
- Winther, K.S.; Gerdes, K. Enteric virulence associated protein VapC inhibits translation by cleavage of initiator tRNA. Proc. Natl. Acad. Sci. USA 2011, 108, 7403–7407. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, Y. Prokaryotic toxin-antitoxin systems: novel regulations of the toxins. Curr. Genet. 2016, 62, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pino, A.; Balasubramanian, S.; Wyns, L.; Gazit, E.; De Greve, H.; Magnuson, R.D. Allostery and intrinsic disorder mediate transcription regulation by conditional cooperativity. Cell 2010, 142, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, M.; Borch, J.; Jorgensen, M.G.; Gerdes, K. Messenger RNA interferase RelE controls relBE transcription by conditional cooperativity. Mol. Microbiol. 2008, 69, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Winther, K.S.; Gerdes, K. Regulation of enteric vapBC transcription: Induction by VapC toxin dimer-breaking. Nucleic Acids Res. 2012, 40, 4347–4357. [Google Scholar] [CrossRef] [PubMed]
- Buts, L.; Lah, J.; Dao-Thi, M.H.; Wyns, L.; Loris, R. Toxin-antitoxin modules as bacterial metabolic stress managers. Trends Biochem. Sci. 2005, 30, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Behiels, E.; Devreese, B. Toxin-Antitoxin systems: Their role in persistence, biofilm formation, and pathogenicity. Pathog. Dis. 2014, 70, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Kedzierska, B. Regulating toxin-antitoxin expression: Controlled detonation of intracellular molecular timebombs. Toxins 2014, 6, 337–358. [Google Scholar] [CrossRef] [PubMed]
- Kopfmann, S.; Roesch, S.K.; Hess, W.R. Type II toxin-Antitoxin systems in the unicellular cyanobacterium Synechocystis sp. PCC 6803. Toxins 2016, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.G.; Ye, S.; Liu, B.; Chang, J.N. The proteolytic activation of the relNEs (ssr1114/slr0664) toxin–antitoxin system by both proteases Lons and ClpP2s/Xs of Synechocystis sp. PCC 6803. Curr. Microbiol. 2011, 63, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.G.; Liu, S.B.; Xu, W.D.; Zhuang, Q.; Wen, C.W.; Tang, X.X. Transcriptional and proteolytic regulation of the toxin-antitoxin locus vapBC10 (ssr2962/slr1767) on the chromosome of Synechocystis sp. PCC 6803. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Srikumar, A.; Krishna, P.S.; Sivaramakrishna, D.; Kopfmann, S.; Hess, W.R.; Swamy, M.J.; Lin-Chao, S.; Prakash, J.S. The Ssl2245-Sll1130 toxin-antitoxin system mediates heat-induced programmed cell death in Synechocystis sp. PCC6803. J. Biol. Chem. 2017, 292, 4222–4234. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Klahn, S.; Scholz, I.; Matthiessen, J.K.; Hess, W.R.; Voss, B. Comparative analysis of the primary transcriptome of Synechocystis sp. PCC 6803. DNA Res. 2004, 21, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.T.; Balsa, D.; Espinosal, M. One cannot rule them all: Are bacterial toxins-antitoxins druggable? FEMS. Microbiol. Rev. 2015, 39, 522–540. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.J.; Hergenrother, P.J. Artificial activation of toxin-antitoxin systems as an antibacterial strategy. Trends Microbiol. 2012, 20, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Christensen, S.; Gerdes, K. Rapid induction and reversal of a bacteriostatic condition by controlled expression of toxins and antitoxins. Mol. Microbiol. 2002, 45, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.G.; Jiang, Y.; Liu, Z.Y.; Xu, Q.G. Characterization of a chromosomal type II toxin-antitoxin system mazEaFa in the cyanobacterium Anabaena sp. PCC 7120. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Shah, D.; Spoering, A.; Kaldalu, N.; Lewis, K. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 2004, 186, 8172–8180. [Google Scholar] [CrossRef] [PubMed]
- Korch, S.B.; Contreras, H.; Clark-Curtiss, J.E. Three Mycobacterium tuberculosis Rel toxin-antitoxin modules inhibit mycobacterial growth and are expressed in infected human macrophages. J. Bacteriol. 2009, 191, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Robson, J.; McKenzie, J.L.; Cursons, R.; Cook, G.M.; Arcus, V.L. The vapBC operon from Mycobacterium smegmatis is an autoregulated toxin-antitoxin module that controls growth via inhibition of translation. J. Mol. Biol. 2009, 390, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Amitai, S.; Yassin, Y.; Engelberg-Kulka, H. MazF-mediated cell death in Escherichia coli: A point of no return. J. Bacteriol. 2004, 186, 8295–8300. [Google Scholar] [CrossRef] [PubMed]
- Prentki, P.; Krisch, H.M. In vitro insertional mutagenesis with a selectable DNA fragment. Gene 1984, 29, 303–313. [Google Scholar] [CrossRef]
- Bates, P.A.; Kelley, L.A.; MacCallum, R.M.; Sternberg, M.J.E. Enhancement of protein modelling by human intervention in applying the automatic programs 3D-JIGSAW and 3D-PSSM. Proteins 2001, 45 (Suppl. S5), 39–46. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Rosenstrom, P. Dali server: Conservation mapping in 3D. Nucleic Acids Res. 2010, 38, W545–W549. [Google Scholar] [CrossRef] [PubMed]
- Schuster-Bockler, B.; Bateman, A. Visualizing profile-profile alignment: Pairwise HMM logos. Bioinformatics 2005, 21, 2912–2913. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, J.L.; Duyvestyn, J.M.; Smith, T.; Bendak, K.; Mackay, J.; Cursons, R.; Cook, G.M.; Arcus, V.L. Determination of ribonuclease sequence-specificity using Pentaprobes and mass spectrometry. RNA 2012, 18, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Brzozowska, I.; Zielenkiewicz, U. Regulation of toxin-antitoxin systems by proteolysis. Plasmid 2013, 70, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Sato, S.; Kotani, H.; Tanaka, A.; Asamizu, E.; Nakamura, Y.; Miyajima, N.; Hirosawa, M.; Sugiura, M.; Sasamoto, S.; et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions (supplement). DNA Res. 1996, 3, 185–209. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Genotype/Plasmid Characteristics * | Source or Reference |
|---|---|---|
| Strains | ||
| E. coli DH5α | F−, φ80d lacZΔM15, Δ(lacZYA-argF) U169, deoR, recA1, endA1, hsdR17(rk−, mk+), phoA, supE44, λ−, thi-1, gyrA96, relA1 | Stratagene (San Diego, CA, USA) |
| E. coli BL21(DE3) | F−, ompT, hsdSB(rB− mB−), gal(λ cI857, ind1, Sam7, nin5, lacUV5-T7gene1) dcm(DE3), lon | Stratagene |
| Synechocystis sp. PCC 6803 | wild type (WT) | ATCC 27184 (Manassas, VA, USA) |
| Plasmids | ||
| pMD-18T | pUC replicon, Apr | Takara Biotech (Dalian, China) |
| pET30a | pBR replicon, lacIq, PT7lac, Kmr | Novagen (Madison, WI, USA) |
| pJS298 | pBR replicon, PT7lac, PBAD, Kmr | [27] |
| pJS307 | pJS298, PT7lac-vapC15, Kmr | This study |
| pJS357 | pJS298, PT7lac-vapC15, PBAD-vapB15, Kmr | This study |
| pJS371 | pJS298, PT7lac-lons, Kmr | [27] |
| pJS391 | pJS298, PT7lac-clpxp2s, Kmr | [27] |
| pJS666 | pET30a, PT7lac-vapBC15, Kmr | This study |
| pJS694 | pMD-18T, PvapBC15, Apr | This study |
| pJS744 | pJS298, PBAD-vapBC15, PT7lac-lons, Kmr | This study |
| pJS745 | pJS298, PBAD-vapBC15, PT7lac-clpxp2s, Kmr | This study |
| pJS759 | p15 replicon, promoter-less lacZ, Spr | [28] |
| pJS766 | pMD-18T, PvapBC15-vapB15, Apr | This study |
| pJS778 | pJS759, PvapBC15-lacZ, Spr | This study |
| pJS779 | pJS759, PvapBC15-vapB15-lacZ, Spr | This study |
| pJS882 | pJS298, PBAD-vapB15, PT7lac-lons, Kmr | This study |
| pJS883 | pJS298, PBAD-vapB15, PT7lac-clpxp2s, Kmr | This study |
| pJS956 | pMD-18T, PvapBC15-vapBC15, Apr | This study |
| pJS962 | pJS759, PvapBC15-vapBC15-lacZ, Spr | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, Q.; Gao, E.-B.; Liu, B.; Wei, Y.; Ning, D. A Toxin-Antitoxin System VapBC15 from Synechocystis sp. PCC 6803 Shows Distinct Regulatory Features. Genes 2018, 9, 173. https://doi.org/10.3390/genes9040173
Fei Q, Gao E-B, Liu B, Wei Y, Ning D. A Toxin-Antitoxin System VapBC15 from Synechocystis sp. PCC 6803 Shows Distinct Regulatory Features. Genes. 2018; 9(4):173. https://doi.org/10.3390/genes9040173
Chicago/Turabian StyleFei, Qian, E-Bin Gao, Biao Liu, Yao Wei, and Degang Ning. 2018. "A Toxin-Antitoxin System VapBC15 from Synechocystis sp. PCC 6803 Shows Distinct Regulatory Features" Genes 9, no. 4: 173. https://doi.org/10.3390/genes9040173