Genome-Wide Identification of Hsp70 Genes in the Large Yellow Croaker (Larimichthys crocea) and Their Regulated Expression Under Cold and Heat Stress

1
Key Laboratory of Sustainable Utilization of Technology Research for Fisheries Resources of Zhejiang Province, Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds, Ministry of Agriculture, Marine Fishery Research Institute of Zhejiang Province, Zhoushan 316002, China
2
Fishery College, Zhejiang Ocean University, Zhoushan 316022, China
*
Author to whom correspondence should be addressed.
Genes 2018, 9(12), 590; https://doi.org/10.3390/genes9120590
Submission received: 13 October 2018
/
Revised: 26 November 2018
/
Accepted: 27 November 2018
/
Published: 29 November 2018
(This article belongs to the Section Animal Genetics and Genomics)
Abstract
:Heat shock proteins 70 (Hsp70) are required for key cellular processes and responses to environmental changes, however, there are an unknown number of hsp70 gene family members in the large yellow croaker (Larimichthys crocea). In the present study, 17 hsp70 genes were identified through the genome of the large yellow croaker. These genes are divided into seven evolutionarily distinct groups according to a phylogenetic tree. The orthologs of these hsp70 genes were found in humans and zebrafish. The expression patterns of the hsp70 gene family in the large yellow croaker under cold and heat stress were studied by examining transcriptome data. Six out of 17 genes were significantly unregulated or downregulated after cold or heat stress. There were two genes significantly upregulated and two genes downregulated in the liver after cold treatment, while after heat treatment, five genes were significantly upregulated, and no genes were significantly downregulated. Three expression patterns were detected: strictly heat-inducible hsp70, constitutively expressed and moderately heat-inducible hsp70, and constitutively expressed and less stress-dependent hsp70 genes. All the findings will contribute to a better understanding of the biological function of hsp70s in defending against thermal challenges.
1. Introduction
Heat shock proteins (Hsps) are a large group of molecular chaperones. They are produced by cells in response to high temperature, hypoxia, infection, toxins, and a number of other types of stress. Some Hsps genes are named housekeeping proteins with constitutively expressed in non-stressed cells [1]. Heat shock proteins play crucial roles in restoring damaged proteins to their functional three-dimensional structure under external environmental stress [2]. Seven different major families, Hsp110, Hsp90, Hsp70, Hsp60, Hsp40, Hsp10, and small Hsps, are classified by their molecular weight [3]. Of these, the hsp70 gene family is an important member of the Hsps superfamily, as it is widely distributed in all organisms from bacteria to mammals, except for some archaea [4,5,6].
Since they were first identified in the early 1960s [7], hsp70 genes have been of great interest as they play critical roles in a series of biochemical processes, including the assembly of multimeric protein complexes, facilitating the intracellular folding of proteins, protein transportation across membranes, and the regulation of the heat-shock response [8]. They are expressed rapidly to protect organisms from environmental stress [9]. The heat shock proteins 70 gene family members provide a potent buffering system for adaption of cellular stress, either from extrinsic (pathogenic and environmental) or intrinsic stimuli (physiological, replicative, or oncogenic) [10].
The heat shock proteins 70 gene family members have different regulatory patterns at transcriptional level. The inducible members of hsp70 genes (strictly inducible) exhibit a low basal level of expression and are only expressed under stress conditions, which typically lack introns. The stress includes, but is not limited to, heat stress [11]. The proteins of the heat-shock genes serve critical functions in protecting cellular proteins from damage under stressful conditions [12]. In contrast, the constitutively expressed Hsps (heat-shock cognates; hsc70s), which assist in the folding of newly translated proteins, are present in cells under all conditions. Heat-shock cognate 70 (hsc70s) typically contain several introns in the coding sequence [2].
The large yellow croaker, Larimichthys crocea, belongs to the family Sciaenidae, order Perciformes, and it is a temperate-water migratory fish [13]. It is mainly distributed from the southern Yellow Sea to the northern South China Sea [14]. As once one of the top four marine species in China, the catch production of large yellow croaker reached about 200,000 tons in the mid-1970s [15]. However, due to over-fishing spawn and overwinter stocks, fishery resources in terms of the large yellow croaker had suffered a severe decline since 1975 [15]. With the advance of artificial cultures, L. crocea has been gradually cultured in the coastal waters of the Zhejiang and Fujian provinces. The expansion of the artificial breeding scale and high density from the 2000s has led to the obvious decline of its immunity and caused lack of resistance to disease for survival in the aquatic environment [16]. High-density farming and temperature changes exacerbate the occurrence of diseases. The heat and cold stress have a serious impact on the survival rate of L. crocea [13]. The optimum water temperature for the growth of large yellow croaker is 18–25 °C. The large yellow croaker cannot tolerate high temperatures above 34 °C and low temperatures below 7 °C; the fish will reduce food intake when seawater temperatures are above 28 °C or below 14 °C [17,18].
Although many genetic studies have reported the expression pattern of some hsp70 genes, the nomenclature for hsp70 is exceptionally complex, generating a confusing array of data and naming conventions in databases and published literature. In some cases, it is impossible to know exactly which gene or protein in the family is being studied, since only the term hsp70 is used without further clarification for the gene or protein [4].
Analyses of hsp70 genes at the genome level to successfully identify new members have been carried out for a few species, such as humans, the channel catfish (Ictalurus punctatus), and the scallop (Patinopecten yessoensis) [1,4,10]. In humans, 17 identified genes were divided into seven evolutionarily distinct groups with distinguishable characteristic according to phylogenetic relationship [4]. In the channel catfish genome, 16 hsp70 genes were detected and 10 out of 16 genes were significantly upregulated or downregulated after bacterial challenges [1]. However, in the large yellow croaker, only one member of the hsp70 gene family has been identified and characterized [16]. There is still no genome-wide characterization of the hsp70 gene family in the large yellow croaker to date. The large yellow croaker genomic resources and transcriptome sequences have been well developed in recent years [13,19]. These resources have made it feasible to conduct a systematic analysis of the genes of interest in the large yellow croaker genome.
A coherent, unifying informational framework encompassing all hsp70 members that would facilitate an understanding of Hsps individually is lacking in teleost. Despite increasing demonstration of the role of hsp70 genes in response to environmental stress in the large yellow croaker and other fish, some issues remain unclear. These include, for example, the constitution and evolutionary features of the hsp70 gene family, the association between gene structure and function, and a systematic analysis of hsp70 gene family involvement in environmental stress. In the present study, we conducted a genome-wide identification of a full set of hsp70 genes and investigated their gene expression in the large yellow croaker under cold and heat stress. Seventeen hsp70 genes were identified in the genome of the large yellow croaker, and the expression patterns in response to cold and heat were examined. Our observations highlight the molecular evolutionary properties and the response mechanism to temperature stress of hsp70 genes in the large yellow croaker. Our findings will provide useful genetic resources to unravel the expression patterns and functions of hsp70 genes in future studies and will contribute to understanding the evolutionary history of hsp70 genes in teleosts.
2. Materials and Methods
The genome sequence, amino acid sequences, and transcriptome data of the large yellow croaker were downloaded from National Center for Biotechnology Information (NCBI) databases (accession number JRPU00000000.2) [20]. In order to identify the full set of hsp70 genes, we used two strategies to search the large yellow croaker genome. First, BlastP (standard protein basic local alignment search tool (BLAST)) searches were performed against amino acid sequences of the large yellow croaker using hsp70s identified from humans and zebrafish as query sequences. Second, the hidden Markov model (HMM) profile of the Hsp70 domain (PF00012) was downloaded from the Pfam protein families database [21], and the HMM profiles of Hsp12a and Hsp12b (PTHR14187:SF46 and PTHR14187:SF3) were obtained from the PANTHER classification system [22]. These were exploited for the identification of hsp70 genes from the yellow croaker genome using the software HMMER 3.0 [23]. The conserved domains of the candidate proteins were survey by online program SMART [24] and the National Center for Biotechnology Information (NCBI) conserved domain database [25]. Furthermore, the obtained full conserved domain sequences (CDS) of proteins from the large yellow croaker genome were used as queries to search against this species in RNA-seq datasets.
The protein sequences of hsp70 genes identified from the large yellow croaker and other selected species were used to construct phylogenetic trees. The representative species for clustering and phylogenetic analysis were human (Homo sapiens), mouse (Mus musculus), platypus (Ornithorhynchus anatinus), chicken (Gallus gallus), Chinese softshell turtle (Pelodiscus sinensis), African clawed frog (Xenopus laevis), zebrafish (Danio rerio), medaka (Oryzias latipes), Nile tilapia (Oreochromis niloticus), torafugu (Takifugu rubripes), stickleback (Gasterosteus aculeatus), and anole lizard (Anolis carolinensis). The hsp70 sequences from these species were retrieved from the NCBI (http://www.ncbi.nlm.nih.gov), Ensembl (http://asia.ensembl.org/index.html), and UniProt (http://www.uniprot.org) databases as listed by Song et al., 2016 [1] (Table S1). Multiple protein sequence alignments were conducted using the Multiple Sequence Comparison By Log-expectation (MUSCLE) method in the program MEGA 7.0 [26]. A maximum likelihood phylogenetic tree was constructed using MEGA 7.0 with a bootstrap test of 1000 replicates based on the LG + G model (amino acid replacement matrices incorporated the gamma distribution for modeling rate heterogeneity G) [27]. The LG + G model was selected as the best-fit model by the ProtTest program (version 3.4) [28].
The Multiple Expectation Maximization for Motif Elicitation software (MEME, version 4.8.1) was used to determine the conserved DNA sequence motifs in the hsp70s of large yellow croaker [29]. The parameters of MEME for detecting motif were the following: site distribution was set at 0 or 1 occurrence per sequence, the maximum number of motifs to be found was 15, and the motif width varied from 10 to 50. The diverse exon–intron organization of the hsp70 genes was identified by comparing predicted coding sequences with their corresponding genomic sequences using the Gene Structure Display Server (version 2.0) [30].
Expression Analysis of Heat Shock Proteins 70 Genes Under Thermal and Hypoxia Stress
To determine the expression profiles of hsp70 genes after cold and heat treatment, the RNA-seq reads were retrieved from cold and heat challenge experiments in the large yellow croaker (SRR1964642-SRR1964649) [17]. A total of 160 individual large yellow croakers were divided in three groups: cold stress group (n = 60), heat stress group (n = 60), and control group (natural temperature, NT, n = 40). The temperatures for the three groups were 9 °C, 31 °C, and 22 °C, respectively. Liver tissues for transcriptome analysis were harvested from six fish 12 h after heat stress and cold stress.
After quality control, RNA sequencing reads were aligned independently with the large yellow croaker genome (GenBank accession no. JRPU00000000.1) [13] using TopHat (v2.0.3) software [31]. The mapped reads were then assembled into transcripts by Trinity (version 2.5.1), guided by a reference annotation of the large yellow croaker genome. Differential gene expressions were analyzed by RNA-seq by expectation-maximization (RSEM). The expression estimations of hsp70 genes were normalized and represented in the form of RPKM (reads per kilobase per million mapped reads), and fold change (log2) values were calculated through the ratio of gene expression from the treated group compared to the control group. Genes with absolute log2 values > 2 and t test values (p < 0.05) after Bonferroni correction served as significantly differential expressed genes. Based on the log2 values, a heat map was generated by the Multiple Experiment Viewer 4.90 (MeV 4.90) [32].
Six significantly differential expressed genes were selected to validate RNA sequencing data by quantitative real-time polymerase chain reaction (qPCR) with gene specific primers designed using Primer v5.0 software (Premier Biosoft International). Primers are listed in Supplement Table S2. The ideal dilution times of cDNA samples were designed by the standard curves. The cDNA samples of template for PCR were diluted 20-fold with nuclease-free water in the procedure of qPCR analysis, following the manufacturer’s instructions for the SYBR® Premix Ex TaqTM (Tli RNaseH Plus, Dalian, China). Using the ABI PRISM 7300 real-time PCR system (Applied Biosystems, Carlsbad, CA, USA) a reaction system of 25 µL was amplified. In order to increase the veracity of the result, three parallel experiments of every cDNA template were conducted The data were analyzed with ABI7300 SDS software (version 1.8, Applied Biosystems). The relative expression levels of all target unigenes or contigs were calculated by the 2−ΔΔCT analysis method to analyze the expression level of genes. We chose the β-actin gene as a reference gene for internal standardization.
3. Results
3.1. Genome-Wide Identification of the Heat Shock Proteins 70 Gene Family in the Large Yellow Croaker
Basic local alignment search tool (BLAST) and HMM searches were conducted to extensively search hsp70 genes in the genome of the large yellow croaker using human and zebrafish protein sequences as queries. A total of 26 putative hsp70 genes were initially identified. Based on the valuation of Pfam and SMART scans, we discarded nine candidate sequences without the hsp70 domain. Eventually, we obtained a total of 17 members representing the unique hsp70 gene family in the large yellow croaker and created robust nomenclature following the Zebrafish Nomenclature Guidelines (https://wiki.zfin.org/display/general/ZFIN+Zebrafish+Nomenclature+Conventions).
The full length of coding sequences among these genes were identified in both transcriptome and genome databases. Detailed information about each hsp70 gene is shown in Table 1. The predicted molecular weights varied from 46.4 kDa (hspa12a) to 110.2 kDa (hyou1). The hspa12a gene had the shortest conserved domain with 399 amino acids, and the longest domain (694 amino acids) was found in the gene hspa4l.1.
3.2. Phylogenetic Relationships of the Heat Shock Proteins 70 Genes Among Species
Based on the amino acid sequences of Hsp70s, an unrooted phylogenetic tree for the hsp70 family was generated to investigate the evolutionary relationships and elucidate classification in this gene family (Figure 1). Some gene duplications were revealed. Two copies of large yellow croaker hsp70 genes were found highly orthologous to zebrafish hspa5 and hspa4l, respectively. These were named hspa5.1, hspa5.2, hspa4l.1, and hspa4l.2 after the zebrafish orthologies. Two related human HSPA8A genes were detected in the teleost fish genome, which were named hspa8a and hspa8b based on the cluster. The teleost-specific genes indicated that hspa8a and hspa8b were present after the divergence between teleost and Tetrapoda. The hsp70s gene names in the large yellow croaker were standardized following the zebrafish and human orthologs. The hsp70s from different species were more closely related to those in the same gene than to others in the same species. All members of the large yellow croaker hsp70 gene family were well distributed into distinct groups and first clustered with corresponding genes of other fish species with strong bootstrap values (Figure 1).
The phylogenetic tree of hsp70 family members from model species revealed seven major evolutionarily groups (Figure 1), with bootstrap support values over 87%. Group I, the largest group, included the hsp70, hspa1a, hspa1b, and hspa8 genes. Group II, Group III and Group IV contained the hspa5, hspa9, and hspa13 genes, respectively. Group V contained the closely related hyou1, hspa4, hspa4L, and hspha1 genes. Group VI was composed of one gene, hspa14. Group VII was composed of the most diverged genes, hspa12a and hspa12b, which were quite dissimilar from the other hsp70 genes.
The large yellow croaker has 12 homologous genes of hsp70 that are also present in humans, with the exception of hspa2, hspa6, and hspa7. The hsph1 gene was not found in any teleosts in the present study except in zebrafish. Most pairs of orthologs from the large yellow croaker and all other fish species were identified, indicating that the common ancestral genes of the hsp70 family might have existed before the genetic divergence of fish species. In addition, two pairs of duplicated hsp70 genes were found in the large yellow croaker, suggesting that hsp70 genes underwent some duplication events after speciation. In Group I, three fish genes (hspa8a, hspa8b, and hsc70) showed a close relationship to the human HSPA8, showing possible paralogs (hsc70 vs. hspa8a and hspa8b).
3.3. Gene Structure and Motif Analysis of the Heat Shock Proteins 70 in the Large Yellow Croaker
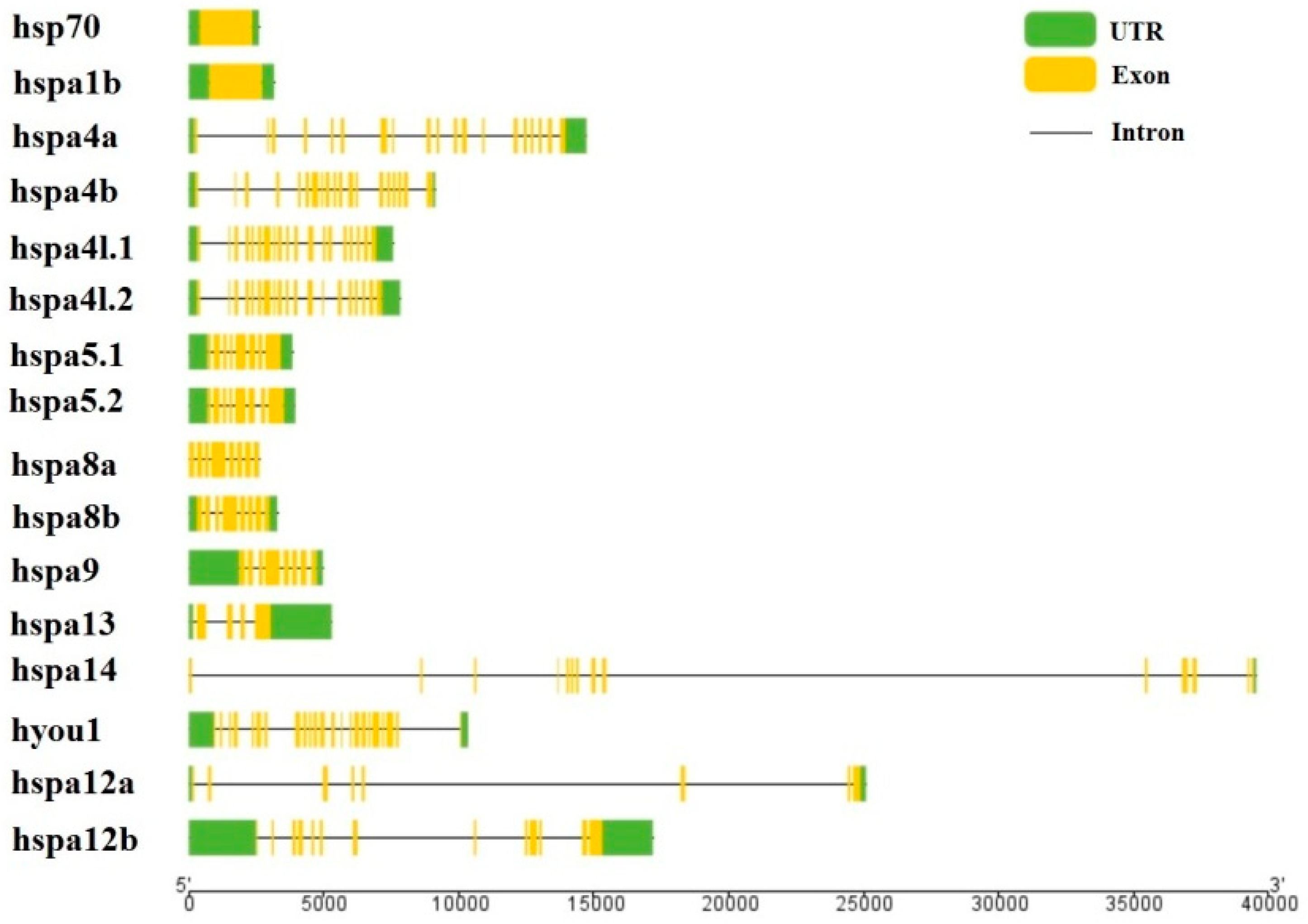
We used the genome annotation file to analyze the gene structure of the large yellow croaker hsp70 genes for better understanding the evolutionary conservation of this gene family. The exon-intron structure of each hsp70 gene was showed in Figure 2. The intron–exon numbers vary greatly in these hsp70 genes, indicating potential biological function diversities among those genes. However, the paralogous gene pairs derived from phylogenetic analysis shared a similar gene structure. However, the number of introns in these genes ranged from 0–13 (Figure 2). Based on the number of introns, genes could be divided into two patterns: pattern 1 with no introns and pattern 2 containing more than one intron. Only hsp70 and hspa1b were assigned to pattern 1. Pattern 2 was common and contained 15 other genes.
To understand the functional diversification of the hsp70 gene family, a conserved motif analysis was performed (Figure 3). We searched for 15 putative motifs in each gene as shown in Figure 3. In general, the Hsp70s from the same groups shared the similar motifs. The motifs were highly conserved within closely related hsp70 members. Moreover, some hsp70 from sister branches even had common motif compositions. Such phenomena were correlated with the gene structures and phylogenetic relationships. In the present study, the recently duplicated hsp70 homologs exhibited similar motif arrangement architectures in their protein structures (Figure 3). Motifs 1, 2, 5, 6, 7, 11, and 13 were present in most hsp70 genes. Only two motifs were detected in hspa12a and hspa12b.
3.4. Expression Regulation of the Heat Shock Proteins 70Gene Under Thermal Stress Treatment
In order to assess the responses of the hsp70 genes to thermal stress at the transcriptome level, RNA data from the livers of large yellow croakers after cold stress treatment, heat stress treatment, and normal temperature were analyzed. Based on these transcriptome data, the involvement of hsp70 genes under cold and heat treatment was studied. Among the 17 hsp70 genes, 12 members were expressed in liver tissue (Figure 4), which indicated a tissue expression pattern. As a result, six of the 17 hsp70 genes were significantly differentially expressed by more than four-fold change (log2 fold change > 2) under cold treatment, including two upregulated genes (hsp70, hspa4a; log2 fold change: 2.03 to 2.17) and two downregulated genes (hspa5.1 and hspa5.2; fold change: −4.49 to −4.85; Table 2, Figure 4 and Figure 5). Five significantly upregulated genes (log2 fold change: 2.01 to 7.32) were found under heat treatment, and no significantly downregulated genes were detected in this treatment (Table 2, Figure 4 and Figure 5). When comparing the cold and heat groups, five significantly upregulated genes (hs70, hspa5.1, hspa5.2, hspa8b and hyou1) were detected, while no significantly downregulated genes were found. The expression of six hsp70 genes (hspa4b, hspa8a, hspa9, hsapa13, hspa14, and hyou1) showed little change in expression levels (Figure 4 and Figure 5) after thermal stress.
3.5. Validation of RNA-Seq Results by Quantitative Polymerase Chain Reaction
To validate the RNA-Seq results, six significant expressed genes were used for qPCR analysis (Figure 6). The results revealed that the expression trends of qPCR findings were consistent with the RNA-Seq results. Generally, the reliability and accuracy of the RNA-Seq analysis were confirmed by the qPCR results.
4. Discussion
Although regulated expression of one hsp70 gene under thermal stress has been reported in the large yellow croaker [16], a systematic analysis of hsp70 involvement in response to thermal stress had not been conducted in this species. Therefore, we performed an overall analysis of the hsp70 gene family in the large yellow croaker, including an analysis of their gene structure, conserved motifs, phylogeny, and expression patterns under cold and heat stress.
A total of 17 hsp70 genes were identified and annotated in the large yellow croaker. Compared to humans, most members of the hsp70 gene family were found in the large yellow croaker, except hspa2, hspa6, hspa7, and hsph1 found in humans Although exhaustive searches were conducted with genomic resources of the large yellow croaker, hsph1, hspa6, and hspa7 were not found in the large yellow croaker genome. The hsph1 gene was only found in zebrafish and was absent in other fish species, including the large yellow croaker, medaka, Nile tilapia, torafugu, and stickleback. The hspa6 and hspa7 genes were not found in any of the six fish species.
Early analyses based on sequences of typical Hsp70s in fish revealed two clusters, named “fish Hsp70-1” and “fish Hsp70-2” [9]. Our phylogenetic analysis of Hsp70s from humans, mice, chickens, zebrafish, the large yellow croaker, and other fish species can be sorted into seven distinct groups, which were supported by other sequence features, such as the exon–intron gene structure. The phylogenetic tree unveiled two major evolutionary events in fish Hsp70s: first, the duplication of hspa4 into two separate genes (corresponding to hspa4a and hspa4b) in teleosts seems to have occurred after the divergence between teleosts and tetrapods (amphibians, reptiles, birds, and mammals). This was a newer duplication than the duplication of hspa4 and hspa4l, which appeared to have occurred early in the evolution of the vertebrate lineage, before the appearance of tetrapods) [4]. Second, the close relation between the hspa8a and hspa8b genes in the large yellow croaker suggests that they duplicated after the divergence of the large yellow croaker and zebrafish, with the hspa8a gene in the zebrafish as a separate subcluster. Third, in the large yellow croaker, we found two recently diverged copies in hspa4l and hspa5 (no sequence variation in CDS). The present results indicate that the teleost tended to have more duplications than mammals, likely as a consequence of whole genome duplication. In the channel catfish, three tandem repeats of hspa8 were detected [1].
Six out of 17 hsp70 genes were significantly upregulated or downregulated under thermal stress, indicating their involvement in thermal adaption. However, different expressed patterns were observed under cold and heat stress. There were two genes significantly upregulated and two genes downregulated in the liver after cold treatment, while after heat treatment, five genes were significantly upregulated and no genes were significantly downregulated. The different patterns may be attributed to the different influence of cold and heat on physiology. In general, cold stress usually reduces rates of enzymatic reactions, diffusion, and membrane transport (whereas heat stress would tend to accelerate these processes) [33]. Under cold stress, only two genes (hspa5.1 and hspa5.2) were significantly downregulated. As an endoplasmic reticulum (ER) chaperone protein, the hspa5 protein plays a key role in protein folding and quality control of protein synthesis in the ER lumen [34]. With decreasing temperature, the synthesis of proteins in the ER is restrained. Therefore, the expression of the hspa5 genes participated in the correct folding of proteins and the degradation of misfolded proteins was limited by cold stress. The expression of hspa5.1 was significantly upregulated under heat stress. The expression of the hspa5 genes under cold and heat treatments was temperature induced. A similar result was reported in rainbow trout (Oncorhynchus mykiss). The hspa5 mRNA expression of rainbow trout was significantly higher at 25 and 26 °C than that at 18 °C (p < 0.05) [35]. Based on the transcriptomic data for rainbow trout, hspa5 was a significantly upregulated gene under heat stress [36]. Moreover, five genes (hsp70, hspa4a, hspa5.1, hsc70, and hspa8b) were significantly upregulated under heat stress. Under cold stress, hspa4 and hsp70 were also significantly upregulated. In the present study, there is a new finding for the Hspa13 gene, which is induced by heat and cold stress in the large yellow croaker. However, the human HSPA13 gene is induced by increases in cytoplasmic calcium but not by heat shock [37]. This finding indicated the different expression of hspa13 in different species. Some studies have demonstrated the higher expression of the hsp70 gene in zebrafish, rohu (Labeo rohita), and sea bass (Dicentrarchus labrax) after heat or cold stress [8,38,39]. The hsp70 gene family can be distinguished by three expression patterns: strictly heat-inducible hsp70, constitutively expressed moderately heat-inducible hsp70, and constitutively expressed and less stress-dependent hsp70 genes. Genes hsp70 and hspa8b are the most inducible gene response to heat stress and belong to the group of strictly heat-inducible hsp70 genes. In the large yellow croacker, hspa4a, hsap5.1, hsap5.2, hsap8a, and hsc70 were constitutive and moderately induced by heat shock, which puts them in the group of moderately heat-inducible hsp70 genes. The hspa4b, hspa9, hspa14, and hyou1 genes, which belong to the group of constitutively expressed and less stress-dependent hsp70 genes, were relatively highly expressed in the cold, control, and heat groups, but showed no significant expression between groups. Previous studies on the hsp70 gene family mainly focused on strictly heat-inducible Hsp70s and cell-cycle-dependent heat-inducible hsp70 genes. The previously identified two distinct groups of the Hsp70 gene family (fish Hsp70–1 and fish Hsp70–2) in reference [9] refer to the heat-inducible Hsp70 and cell-cycle-dependent heat-inducible hsp70 genes.
Despite the lack of biological replicates in the transcriptome data [19], the expression results are still an indication. The pooling sample strategy in thermal treatments could have eliminated individual variation. The purpose of the present study was to provide a preliminary classification of all the members of the hsp70 gene family in the genome of large yellow croaker. This is an essential step towards their unambiguous functional characterization. Our study will provide the basic foundation for further biological study to demonstrate the role of each large yellow croaker hsp70 in thermal stress.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/9/12/590/s1, Table S1: The species accession numbers of Hsp70 cited in the present study; Table S2: Primers used for quantitative real-time PCR (qPCR) validations.
Author Contributions
Conceptualization, K.X. and Z.H.; Methodology, K.X.; Investigation, H.X.; Data Curation, H.X.; Writing-Original Draft Preparation, K.X.; Writing-Review & Editing, Z.H.; Supervision, H.X.; Project Administration, Z.H.; Funding Acquisition, Z.H.
Funding
This research was funded by the National Natural Science Foundation of China (31472281) and the National Key Research and Development Program of China (2017YFA0604904).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Song, L.; Li, C.; Xie, Y.J.; Liu, S.K.; Zhang, J.R.; Yao, J.; Jiang, C.; Li, Y.; Liu, Z.J. Genome-wide identification of Hsp70 genes in channel catfish and their regulated expression after bacterial infection. Fish Shellfish Immun. 2016, 49, 154–162. [Google Scholar] [Green Version]
- Metzger, D.C.; Hemmerhansen, J.; Schulte, P.M. Conserved structure and expression of hsp70 paralogs in teleost fishes. Comp. Biochem. Phys. D 2016, 18, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, D.; Chen, Q.; Ma, L. Genome-wide survey and characterization of the small heat shock protein gene family in Bursaphelenchus xylophilus. Gene 2016, 579, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Brocchieri, L.; Conway, D.M.E.; Macario, A.J. hsp70 genes in the human genome: Conservation and differentiation patterns predict a wide array of overlapping and specialized functions. BMC Evol. Biol. 2008, 8, 19. [Google Scholar]
- Liu, J.; Wang, R.; Liu, W.; Zhang, H.; Guo, Y.; Wen, R. Genome-wide characterization of heat-shock protein 70s from Chenopodium quinoa and expression analyses of Cqhsp70s in response to drought stress. Genes 2018, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Karlin, S.; Brocchieri, L. Heat shock protein 70 family: Multiple sequence comparisons, function, and evolution. J. Mol. Evol. 1998, 47, 565–577. [Google Scholar] [CrossRef]
- Ritossa, F. Discovery of the heat shock response. Cell Stress Chaperon 1996, 1, 97–98. [Google Scholar] [CrossRef]
- Giri, S.S.; Sen, S.S.; Sukumaran, V. Role of HSP70 in cytoplasm protection against thermal stress in rohu, Labeo rohita. Fish Shellfish Immun. 2014, 41, 294–299. [Google Scholar] [CrossRef]
- Yamashita, M.; Yabu, T.; Ojima, N. Stress protein HSP70 in fish. ABSM 2010, 3, 111–141. [Google Scholar] [CrossRef]
- Cheng, J.; Xun, X.G.; Kong, Y.F.; Wang, S.Y.; Yang, Z.H.; Li, Y.J.; Kong, D.X.; Wang, S.; Zhang, L.L.; Hu, X.L.; et al. Hsp70 gene expansions in the scallop (Patinopecten yessoensis) genome and their expression regulation after exposure to the toxic dinoflagellate Alexandrium catenella. Fish Shellfish Immun. 2016, 58, 266–273. [Google Scholar]
- Thanaphum, S.; Haymer, D.S. A member of the hsp70 gene family from the Mediterranean fruit fly, Ceratitis capitata. Insect Mol. Biol. 1998, 7, 63–72. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The function of heat-shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 27, 437–496. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Mu, Y.; Xiang, L.X.; Fan, D.D.; Feng, M.J.; Zhang, S.; Shi, Q.; Zhu, L.Y.; Li, T.; Ding, Y.; et al. Genome sequencing of the perciform fish Larimichthys crocea provides insights into molecular and genetic mechanisms of stress adaptation. PLoS Genet. 2015, 11, e1005118. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.Q.; Xu, H.X.; Shui, B.N.; Zhou, Y.D.; Gao, T.X. Lack of genetic structure in endangered large yellow croaker Larimichthys crocea, from China inferred from mitochondrial control region sequence data. Biochem. Syst. Ecol. 2015, 61, 1–7. [Google Scholar] [CrossRef]
- Liu, M.; Mitcheson, S. Profile of a fishery collapse: Why mariculture failed to save the large yellow croaker. Fish Fish. 2008, 9, 219–242. [Google Scholar] [CrossRef]
- He, J.Y.; Wang, J.R.; Xu, M.S.; Wu, C.W.; Liu, H.H. The cooperative expression of heat shock protein 70 KD and 90 KD gene in juvenile Larimichthys crocea under Vibrio alginolyticus stress. Fish Shellfish Immun. 2016, 58, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Y.; Xue, L.Y.; Huang, H.L. Liver transcriptome analysis of the large yellow croaker (Larimichthys crocea) during fasting by using RNA-seq. PLoS ONE 2016, 11, 0150240. [Google Scholar] [CrossRef]
- Zhang, C.L.; Liu, J.F.; Li, Y.C.; Chen, Z. Analyzing the present condition and countermeasure of cultured large yellow croaker Pseudosciaena crocea in Fujian Province. Shanghai Fish. Univ. 2002, 11, 77–83. [Google Scholar]
- Qian, B.Y.; Xue, L.Y. Liver transcriptome sequencing and de novo annotation of the large yellow croaker (Larimichthys crocea) under heat and cold stress. Mar. Genom. 2016, 25, 95–102. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information Home Page. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 29 November 2018).
- Pfam Home Page. Available online: https://pfam.xfam.org/ (accessed on 15 November 2018).
- PANTHER Classification System Home Page. Available online: http://www.pantherdb.org/ (accessed on 5 October 2018).
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef]
- Simple Modular Architecture Research Tool Home Page. Available online: http://smart.embl-heidelberg.de/ (accessed on 2 November 2018).
- National Center for Biotechnology Information conserved domain database. Available online: http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi/ (accessed on 12 November 2018).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the Second International Conference on Intelligent Systems for Molecular Biology, Stanford, CA, USA, 14–17 August 1994; AAAI Press: Menlo Park, CA, USA, 1994; pp. 28–36. [Google Scholar]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free; open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef]
- Sonna, L.A.; Fujita, J.; Gaffin, S.L.; Lilly, C.M. Molecular biology of thermoregulation invited review: Effects of heat and cold stress on mammalian gene expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef]
- Dana, R.C.; Welch, W.J.; Deftos, L.J. Heat shock proteins bind calcitonin. Endocrinology 1990, 126, 672–674. [Google Scholar] [CrossRef]
- Xia, B.P.; Liu, Z.; Zhou, Y.J.; Wang, Y.J.; Huang, J.Q.; Li, Y.J.; Kang, Y.J.; Wang, J.F.; Liu, X.X. Effects of heat stress on biochemical parameters and heat shock protein family a (Hsp70) member 5 (HSPA5) mRNA expression in rainbow trout (Oncorhynchus mykiss). Mar. Freshw. Res. 2018. [Google Scholar] [CrossRef]
- Huang, J.Q.; Li, Y.J.; Liu, Z.; Kang, Y.J.; Wang, J.F. Transcriptomic responses to heat stress in rainbow trout Oncorhynchus mykiss head kidney. Fish Shellfish Immun. 2018, 82, 32–40. [Google Scholar] [CrossRef]
- Otterson, G.A.; Gregory, C.F.; Kratzke, R.A.; Coxon, A.; Johnston, P.G.; Kaye, J.F. Stch encodes the ‘ATPase core’ of a microsomal stress 70 protein. EMBO J. 1994, 13, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Airaksinen, S.; Jokilehto, T.; Rabergh, C.M.; Nikinmaa, M. Heat-and-cold inducible regulation of HSP70 expression in zebrafish ZF4 cells. Comp. Biochem. Physiol. B 2003, 136, 275–282. [Google Scholar] [CrossRef]
- Bertotto, D.; Poltronieri, C.; Negrato, E.; Richrad, J.; Pascoli, F.; Simontacchi, C.; Radaelli, G. Whole body cortisol and expression of HSP70, IGF-I and MSTN in early development of sea bass subjected to heat shock. Gen. Comp. Endocrinol. 2011, 174, 44–50. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree of the heat shock protein 70 (Hsp70) family based on the maximum likelihood method. The bootstrap consensus tree inferred from 1000 replicates is taken.
Figure 1.
Phylogenetic tree of the heat shock protein 70 (Hsp70) family based on the maximum likelihood method. The bootstrap consensus tree inferred from 1000 replicates is taken.

Figure 2.
Exon-intron structure of large yellow croaker hsp70 genes. Green boxes indicate untranslated 5′- and 3′-regions; yellow boxes indicate exons; black lines indicate introns.
Figure 2.
Exon-intron structure of large yellow croaker hsp70 genes. Green boxes indicate untranslated 5′- and 3′-regions; yellow boxes indicate exons; black lines indicate introns.

Figure 3.
Distribution of conserved motifs in hsp70 of the large yellow croaker. Each colored box represents a putative motif detected in the protein sequence.
Figure 3.
Distribution of conserved motifs in hsp70 of the large yellow croaker. Each colored box represents a putative motif detected in the protein sequence.

Figure 4.
The hsp70 expression of the large yellow croaker based on reads per kilobase per million mapped reads (RPKM) values.
Figure 4.
The hsp70 expression of the large yellow croaker based on reads per kilobase per million mapped reads (RPKM) values.

Figure 5.
Heat map of hsp70 expression under cold and heat stress treatments based on the fold change (log2) in RPKM values.
Figure 5.
Heat map of hsp70 expression under cold and heat stress treatments based on the fold change (log2) in RPKM values.

Figure 6.
Validation of six significantly differential expressed genes by quantitative polymerase chain reaction (qPCR).
Figure 6.
Validation of six significantly differential expressed genes by quantitative polymerase chain reaction (qPCR).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of 17 hsp70 genes identified in the large yellow croaker genome.
| Gene Name | Gene Accession Number | Protein Accession Number | CDS Length (bp) | Protein Length (aa) | Hsp70 Domain Location (aa) | Domain Feature | WT (kDa) | |
|---|---|---|---|---|---|---|---|---|
| 1 | hsp70 | XM_010755062.2 | XP_010753364.1 | 1920 | 639 | 8–614 | HSPA1-2-6-8-like_NBD | 70.3 |
| 2 | hspa1b | XM_010740692.2 | XP_010738994.1 | 1917 | 638 | 7–613 | HSPA1-2-6-8-like_NBD | 70.3 |
| 3 | hspa4a | XM_010729327.2 | XP_010727629.1 | 2514 | 837 | 3–586 | HSPA4_NBD | 93.6 |
| 4 | hspa4b | XM_010754752.2 | XP_010753054.2 | 2535 | 844 | 3–598 | HSPA4_NBD | 94.6 |
| 5 | hspa4l.1 | XM_019277085.1 | XP_019132630.1 | 2523 | 840 | 3–696 | HSPA4_NBD | 94.4 |
| 6 | hspa4l.2 | XM_019279519.1 | XP_019135064.1 | 2493 | 830 | 3–557 | HSPA4_NBD | 93.5 |
| 7 | hspa5.1 | XM_010738795.2 | XP_010737097.1 | 1965 | 654 | 28–634 | HSPA5-like_NBD | 72.2 |
| 8 | hspa5.2 | XM_019274093.1 | XP_019129638.1 | 1965 | 654 | 28–634 | HSPA5-like_NBD | 72.2 |
| 9 | hspa8a | XM_019279481.1 | XP_019135026.1 | 1953 | 650 | 6–612 | HSPA1-2-6-8-like_NBD | 71.1 |
| 10 | hspa8b | XM_019270840 | XP_019126385.1 | 1941 | 646 | 6–612 | HSPA1-2-6-8-like_NBD | 70.9 |
| 11 | hsc70 | XM_010747566.2 | XP_010745868.1 | 1950 | 649 | 6–612 | HSPA1-2-6-8-like_NBD | 71.07 |
| 12 | hspa9 | XM_010734059.2 | XP_010732361.2 | 2043 | 680 | 55–653 | HSPA9-like_NBD | 73.4 |
| 13 | hspa12a | XM_019257914.1 | XP_019113459.1 | 1236 | 411 | 40–389 | HSPA12A-like_NBD | 46.4 |
| 14 | hspa12b | XM_019255647.1 | XP_019111192.1 | 2064 | 687 | 51–584 | HSPA12B-like_NBD | 76.5 |
| 15 | hspa13 | XM_010757137.2 | XP_010755439.1 | 1329 | 442 | 33–431 | HSPA13-like_NBD | 48.3 |
| 16 | hspa14 | XM_019266323.1 | XP_019121868.1 | 1566 | 521 | 18–521 | HSPA14-like_NBD | 56.4 |
| 17 | hyou1 | XM_019279502.1 | XP_019135047.1 | 2961 | 986 | 28–690 | HYOU1-like_NBD | 110.2 |
CDS, conserved domain sequences; Hsp70, heat shock proteins 70; WT, molecular weight.
Table 2.
Log2 fold change of large yellow croaker hsp70 gene expression in liver under thermal stress. The significant genes (p value < 0.05, reads number >10, absolute log2 fold change >2) and their fold changes are denoted with *
Table 2.
Log2 fold change of large yellow croaker hsp70 gene expression in liver under thermal stress. The significant genes (p value < 0.05, reads number >10, absolute log2 fold change >2) and their fold changes are denoted with *
| Gene Name | hsp70 | hspa4a | hspa4b | hspa5.1 | hspa5.2 | hspa8a | hspa8b | hsc70 | hspa9 | hspa13 | hspa14 | hyou1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cold | 2.17 * | 2.03 * | 0.64 | −4.49 * | −4.85 * | 0.33 | −0.92 | 0.68 | 0.28 | 1.66 | 0.90 | −1.27 |
| Heat | 7.32 * | 2.01 * | −0.44 | 2.24 * | 0.71 | 0.88 | 5.33 * | 2.13 * | 0.44 | 1.82 | −0.25 | 0.95 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, K.; Xu, H.; Han, Z. Genome-Wide Identification of Hsp70 Genes in the Large Yellow Croaker (Larimichthys crocea) and Their Regulated Expression Under Cold and Heat Stress. Genes 2018, 9, 590. https://doi.org/10.3390/genes9120590
AMA Style
Xu K, Xu H, Han Z. Genome-Wide Identification of Hsp70 Genes in the Large Yellow Croaker (Larimichthys crocea) and Their Regulated Expression Under Cold and Heat Stress. Genes. 2018; 9(12):590. https://doi.org/10.3390/genes9120590
Chicago/Turabian StyleXu, Kaida, Hanxiang Xu, and Zhiqiang Han. 2018. "Genome-Wide Identification of Hsp70 Genes in the Large Yellow Croaker (Larimichthys crocea) and Their Regulated Expression Under Cold and Heat Stress" Genes 9, no. 12: 590. https://doi.org/10.3390/genes9120590
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.