Antifragility and Tinkering in Biology (and in Business) Flexibility Provides an Efficient Epigenetic Way to Manage Risk

Abstract
: The notion of antifragility, an attribute of systems that makes them thrive under variable conditions, has recently been proposed by Nassim Taleb in a business context. This idea requires the ability of such systems to ‘tinker’, i.e., to creatively respond to changes in their environment. A fairly obvious example of this is natural selection-driven evolution. In this ubiquitous process, an original entity, challenged by an ever-changing environment, creates variants that evolve into novel entities. Analyzing functions that are essential during stationary-state life yield examples of entities that may be antifragile. One such example is proteins with flexible regions that can undergo functional alteration of their side residues or backbone and thus implement the tinkering that leads to antifragility. This in-built property of the cell chassis must be taken into account when considering construction of cell factories driven by engineering principles.1. Introduction
The recent dread of radioactive leaks near nuclear plants reminds us vividly of the vulnerability of life. “Fragile” is an epithet often associated with living organisms, perceived as delicate and easily damaged. But does this reflect reality? What would be the opposite of fragile? In a recent piece for Edge magazine [1] Nassim Taleb discussed another familiar attribute of life, tinkering. There he asserted that, contrary to the favored emphasis on intelligent engineering, “tinkering outperforms design”. This property enables finding unexpected solutions, especially when challenged by “normal accidents” [2], rare deleterious events (in terms of individual probability of occurrence) for which he himself coined the phrase “Black Swan” events [3].
A wealth of studies compared economics and biology [4-6]. For the lay observer of economics, a point shared by both disciplines is the pervasive presence of tinkering. However, this is usually hidden behind the secrets of mathematics and computation (see an illuminating view of “Hollywood Economics” that shows how apparently unrelated individual behaviors may aggregate into coherent outcomes [7]). Tinkering is indeed at the core of life [8]. The biological evolution of unusual structures and processes in response to extreme environments or other types of challenges is ubiquitous. We explore various facets of this notion and see whether it might help us better understand the behavior and evolution of living organisms. In this context, tinkering derives from unexpected (undesigned) properties that are put in action under stressful or difficult circumstances.
Opposing fragility, properties such as “robustness” or “resilience” help make a possibly dynamic entity hard to inactivate or destroy. The expected fate of the entity is sealed: either it hardens for some time, stays the same or it breaks down. Should we extend this view to define a possibly new concept, antifragility? To define antifragility, Taleb opposes the fragility of the Damocles' Sword not to the robustness of the Phoenix (who rises from the ashes), but to the inventiveness of the Hydra (who gets two heads each time one is cut off). Is it possible to think of entities that would not just resist the ravages of time but, through the creation and recombination of novel components, actually become able to cope with an unpredictable future? More generally, can we identify processes that make an entity stronger (sturdier) through the effect of ageing, shock or stress?
In many of its features, life indeed appears to share this property with the Hydra: challenged, it responds by creating new forms of life. This property is at the root of our models of evolution. We use dichotomies to construct the Tree of Life. Of course, many (most) branches are cut off forever—and the Greek Hydra is not an immortal animal—but in the succession of generations, antifragile life tends to expand its exploration of the world by generating a systematic creation of novelty. This is illustrated by the huge number of new life forms that flourish following mass extinctions. On shorter time scales, we know well that it takes only a few months before anticancer drugs are defeated by cells trying to become immortal. In the same way, resistance to an antibiotic follows shortly after its introduction. Moreover, resistance does not simply result from horizontal gene transfers, using some hidden form of pre-existing resistance, but, quite often, from the emergence of a completely novel activity. For example, while the precursor of Nylon® never existed before the creation of this man-made polymer, it did not take long to find bacteria that used it as a nitrogen and carbon source [9]. The enzymes involved have been created de novo from apparently random pieces of DNA [10]. In the same way, emerging diseases are often the progeny of pre-existing diseases. The SARS coronavirus evolved a novel surface antigen [11], and the Escherichia coli O104:H4 strain assembled traits that were distributed in the environment with a completely different pattern [12]. Evolution, then, should be reconsidered in this new light. Living systems are antifragile in that they can do much more than simply respond to the “pressure” of the environment by random mutations followed by selection; they have an in-built property that allows them to find solutions in the face of adversity. Antifragility is one such property. It unfolds not only in individual organisms, which age much slower than what could be expected from a fragile entity, but also in the way they generate a progeny. All these processes have in common the ability to create some novel information: antifragility cannot be separated from management of information [13].
The present reflection is organized in three parts. Rather than study multicellular organisms, where a variety of cells with complicated hierarchies display the functions of life, possibly hiding important underlying principles, we restrict our views to unicellular organisms, bacteria. We first recount the core functions, emphasizing their relative position in the process of ageing. We focus on “steady state” or “stationary” life, which we see as differing from senescence, the continuous degradation of the cell's components as time elapses (in human beings the analogous ageing-before-senescence stage would be roughly between ages 25 and 50). Subsequently we explore the process of antifragility [14] (without bothering to carry over likely challenges about the novelty of the concept) using a specific molecular example. Choosing among a variety of antifragile processes, we discuss the existence of (literally) flexible proteins and the processes in which they participate. We show how this could help delay the senescence process, providing an example of how tinkering and antifragility work together during ageing. The final discussion recapitulates the possible roles of these processes in biology at the cellular level, focusing on the stationary regime.
2. Functions Required to Sustain Life
The status of living organisms which do not reproduce such as the bee or ant workers stands out. Their life is limited in time, and does not perpetuate itself. In the absence of reproduction there would be no life unless it could be created spontaneously at a high rate. Were all organisms suddenly sterilized, life would continue until the last of them dies, and would then disappear. Reproduction must therefore be introduced as a core feature of what life is, beyond functions associated to stationary life. This creates a conflict between reproduction and stationary life, implying some trade-off between reproduction and survival of the individual [15]. Yet, because, by far the longest time spent by most organisms during their lifetime does not involve reproduction, we limit our exploration to life under stationary state. Indeed, if reproduction were allowed to proceed unchecked, the Earth would have been covered by one species that would have subsequently been halted in its tracks by exhaustion of nutrient supplies.
To go further, we use the Synthetic Biology engineering mode of reasoning: What would we need if we were to construct a long-lived organism? This question is essential for scaling up Synthetic Biology processes. It requires us to try and make a thorough inventory of functions, taking care not to forget unobtrusive but essential ones. To this aim, we separate cells into two independent components [16], the cell (the chassis) and its program. As reproduction is not essential during the stationary phase, we restrict our analysis to the functions of the chassis and the associated turnover of its components [17], reserving some thoughts about the constraints at the onset of reproduction to the end of this article. Note that we do not limit ourselves to structures (the term “chassis” is misleading in this respect), but explore processes as well. We further note that during the stationary phase the ever-ageing cell must either entirely freeze its metabolism and lay dormant (as a spore, for example; some Viable But Non-Culturable Cells (VBNC) might also represent this state [18]), or maintain a minimum core metabolism and energy supply, to be ready to start again multiplying when the conditions get better. The organism is in a state of flux, with specific functions involved [17]. Let us try to make a list of these functions.
2.1. The Cell Senses Its Physical State and Exchanges Atoms and Molecules
The cell's chassis combines management of compartmentalisation (the cell's envelope, appendages, but also its nanomachines such as the ribosome, ATP synthase, the degradosome and many others [19]) and metabolism (flux of interconversion of molecules for the building up, storage, salvage of the cell's building blocks, catalytic centers and energy management). Nutrients are imported and waste products are exported. In parallel, the cell monitors its sensors. Two types of sensing are used: sensing the physical state of the environment (temperature, light, pressure, etc) and sensing chemicals. The physical constraints cannot be neglected. Except in homeothermic organisms, changes of environmental temperature are immediately passed on to the organism. Because the effects of temperature on macromolecules are often similar to those of ageing (misfolding is the consequence of both changes), there may be considerable overlap between sensing ageing and sensing temperature variations. Indeed, molecular chaperones have both the role of sensors and of maintenance systems and they belong to the heat shock response [20]. Hence, because these molecules must act on a large number of substrates, we may assume that functions fulfilled by molecular chaperones are to be executed in priority in stationary life.
At the same time, the cell must get a large variety of chemicals from the environment. Fortunately, turnover of metabolites and salvage processes minimize the need to resort to the environment for their supply. Salvage is not a heavy-duty process. Although necessary, it should not belong to the first-line protection because few functional molecules of the corresponding enzymes are needed in the salvage process (this is akin to the marginal cost of a process in the financial world). Similarly, the time-dependent decay of most functions involved in transport of chemicals into the cell can be compensated for through considerable redundancy. The cell only needs to transport a few categories of products: amino acids, simple carbohydrates or dicarboxylates, polyamines, vitamins and ions. This is readily observed in the organisms that can live with a very low number of genes such as Mycoplasma genitalium [21]. Redundancy in the function of transporters, either pre-existing or resulting from the process of ageing, will limit the number needed to be conserved in the long term. Overall, in contrast with the front-line requirement for molecular chaperones, there is no heavy-duty requirement for these functions that may slowly deteriorate over time without dramatic consequences.
2.2. RNA is Degraded and Re-Synthesized
In contrast with the situation just described, many components have to be continuously replaced, possibly with a low turnover. This requires the continued use of the global biosynthetic capacity of the cell. Unfortunately, the identification of essential ubiquitous functions is somewhat difficult, as they cannot be solely identified using comparative genomics. While functions may be ubiquitous, enzyme structures are not [22]. In the present work we used sets of persistent genes [23] to identify the essential functions for stationary life. Consider a bacterium after it has entered stationary phase. RNAs are expected to turn over first, as they are seldom repaired (see however [24]). This process is managed by the degradosome and related activities [25]. The degradosome structure (exosome in Archaea and Eukarya) is highly diverse, being different in gamma-Proteobacteria, alpha-Proteobacteria and Firmicutes, for example [26]. It always contains polynucleotide phosphorylase (PNPase) at its core: This enzyme uses phosphorolysis rather than hydrolysis, generating nucleoside diphosphates instead of nucleoside monophosphates as basic building blocks for nucleic acids, followed by the recovery of one quantum of energy [27]. Associated to PNPase we find a variety of ribonucleases of different descent, such as RNase E in gamma-Proteobacteria or RNase J and RNase Y in Firmicutes [28], energy-dependent helicases, as well as core enzymes of the glycolysis/gluconeogenesis pathway [25]. We also find co-evolving proteins meant to degrade leftovers, such as nanoRNases of various families, depending on the phylogenetic clade [29].
A puzzling observation stems out from the composition of the degradosome: Many of its functions are carried out by proteins that are significantly longer than average bacterial proteins (300 residues). This is counter-intuitive because the synthesis of long proteins is expected to be fragile. The probability of transcription/translation errors, especially the premature termination of translation, is proportional to the length of the messenger RNA. Moreover, it is well known that truncated proteins in complexes display a negative-dominant phenotype [30]. However, the significance of this bias is supported by further observations. RNA turnover implies that the first anabolic function that must be preserved is transcription, because any requirement for re-synthesized macromolecules will first go through this step. RNA polymerase is its core component. Surprisingly, it involves two very large subunits (more than 1,300 residues), RpoB and RpoC. This might have just been a statistical coincidence, but an experiment shows that length is significant. Bacteria such as Helicobacter pylori, which thrive in difficult environments, fuse RpoB and RpoC into a single gigantic protein. Splitting it back into two components makes the cell sensitive to the denaturing agent urea, showing that premature termination followed by re-initiation is not an option for recovery [31]. Other important components of the machinery are also large, e.g., NusA [32] or the transcription-repair factor Mfd [33]. Is there a special property, associated with increased length that could compensate for the fragility of synthesis of long molecules?
2.3. Proteins also Need to be Replaced
Non-functional proteins can be either repaired [34-36] or refolded, and this costs energy. When neither is possible they are degraded. Degradation enzymes, often hydrolytic enzymes, are essential at this stage. Remarkably, while hydrolysis is a highly exothermic process, many degradation enzymes consume energy-an apparent waste. This reminds us of the remark made by Hopfield, that in order to identify important and unexpected functions, we should be exploring “known reactions which otherwise appear to be useless or deleterious complications [yet] are seen to be essential to the [studied] function” [37]. Antifragility implies improving while ageing, before inevitable senescence. This in turn requires a process making room to retain some sort of information.
We expect that energy-dependent degradation is the hallmark of information gathering and utilizing systems [13]. Many energy-dependent proteases have been characterized, including Clp, Lon, and FtsH (HflB) in E. coli. Clp proteases are degradation machines composed of a sensor required for both substrate recognition and ATP-dependent selection for unfolding, and of a peptidase made of multiple subunits, required for substrate hydrolysis (see e.g., [38]). Clp proteases (ClpP) use the ATPase subunits to choose between folded and unfolded substrates. The importance of choosing (i.e., collecting information on) the substrate to be degraded is illustrated by the unexpected role of a novel class of antibiotics, acyldepsipeptides, that are active against ClpP. The acyldepsipeptide-activated ClpP core initiates proteolytic degradation without the control exerted by Clp-ATPases. This unchecked activity, which demonstrates that the energy of ATP is not used in the very process of protein hydrolysis, leads to the inhibition of bacterial cell division and eventually cell death [39]. Again, these proteins, essential during stationary life, are long.
Proteolysis makes room in the cell, and relevant feedbacks resynthesise degraded proteins that are important. To this aim, translation requires utilization of at least part of the population of ribosomes. Some ribosomes are clustered together in “hibernating” 100S complexes combining two 70S ribosomes and Rmf [40]. The available ones must have at least some functional ribosomal proteins (several are somewhat dispensable [41]). tRNA synthetases, initiation and elongation factors are also required in the process. In contrast to the proteins described previously, ribosomal proteins are short or very short proteins, a feature that could have been selected to minimize errors when they are synthesized. Why such a contrast between the translation nanomachine, and the transcription nanomachine? We shall see below that they may have features in common.
2.4. Even Metabolites may be Repaired
Energy is central to all the processes listed above. During stationary life the core functions of energy management (adenylate kinase and nucleoside diphosphokinase in particular) are essential to replenish the energy level of the cell; central intermediary metabolism (with synthesis of phosphoenolpyruvate and acetyl-Coenzyme A) must be present for proper functioning of the cell. Structural features of the cell's envelope must also be important. Also, the murein sacculus (the rigid peptidoglycan structure that gives the cell its shape and protects it against deleterious changes in the environment) is likely to be quite stable and will not require heavy-duty maintenance. The situation may be somewhat different for membrane phospholipids [42]. Finally, in line with the fate of macromolecules, altered metabolites are either degraded, recycled or exported to the environment. Some are repaired (AdoMet via SamT [43], D-amino acids) but this may be optional. Establishing the list of repaired metabolites should tell us something about their relative importance in the functional hierarchy of the cell under stationary conditions (Table 1).

{kind=link}
{kind=link}
{kind=link}
| process | nanomachine | Escherichia coli | Bacillus subtilis | ||
|---|---|---|---|---|---|
| maintenance | |||||
| RNA turnover | degradosome (exosome) | Rne PnpA Eno TpiA Orn PcnB | Rny RnjA PnpA Eno TpiA NrnA NrnB | ||
| protein turnover | proteasome | ClpAXP Lon HslVU FtsH… | ClpXP LonAB ClpCE ClpQ ClpY FtsH | ||
| repair | refolding | Spy DnaJK GrpE GroSL… | DnaJKGrpE GroSL | ||
| restoring | Pcm FrlDB FrlC MsrAB | FrlDB MsrAB | |||
| transcription | RNA polymerase | RpoABC NusA NusG Mfd Sigmas | RpoABC NusAG Mfd Sigmas | ||
| translation | Ribosome and tRNAs | Rps[A-U] Rpl[A-Y] Rpm[A-J] 20 tRNA synthetases Rmf EFTu EFTs EFG Modifications… | Rps[B-U] Rpl[A-Y] Rpm[A-J] 19 tRNA synthetases 1 amidotransferase EFTu EFTs EFG | ||
| folding | chaperones | Tig Ppi DnaJKGrpEGroSL | Tig DnaJKGrpE GroSL | ||
| metabolism | carbon | Eno PykA Pps AceEF Lip Ppa … | Eno Tpi PykA PdhABC PpaC… | ||
| nitrogen | Aminotransferases | Aminotransferases | |||
| phosphorus | Adk Ndk Ppk… | Adk Ndk PpnKA PpnKB | |||
| compartmen-talisation | sensing transporting | Amino acids; nucleosides or bases; vitamins; carbohydrates or dicarboxylates; polyamines; ions | Amino acids; nucleosides or bases; vitamins; carbohydrates or dicarboxylates; polyamines; ions | ||
| replication | repair | chemical alterations, single and double strand breaks and recombination | chemical alterations, single and double strand breaks and recombination | ||
| initiation | primase | control of restart | control of restart |
2.5. Long and Short Proteins Make up the Stationary Essential Proteome, but Proteins of Average Length do not
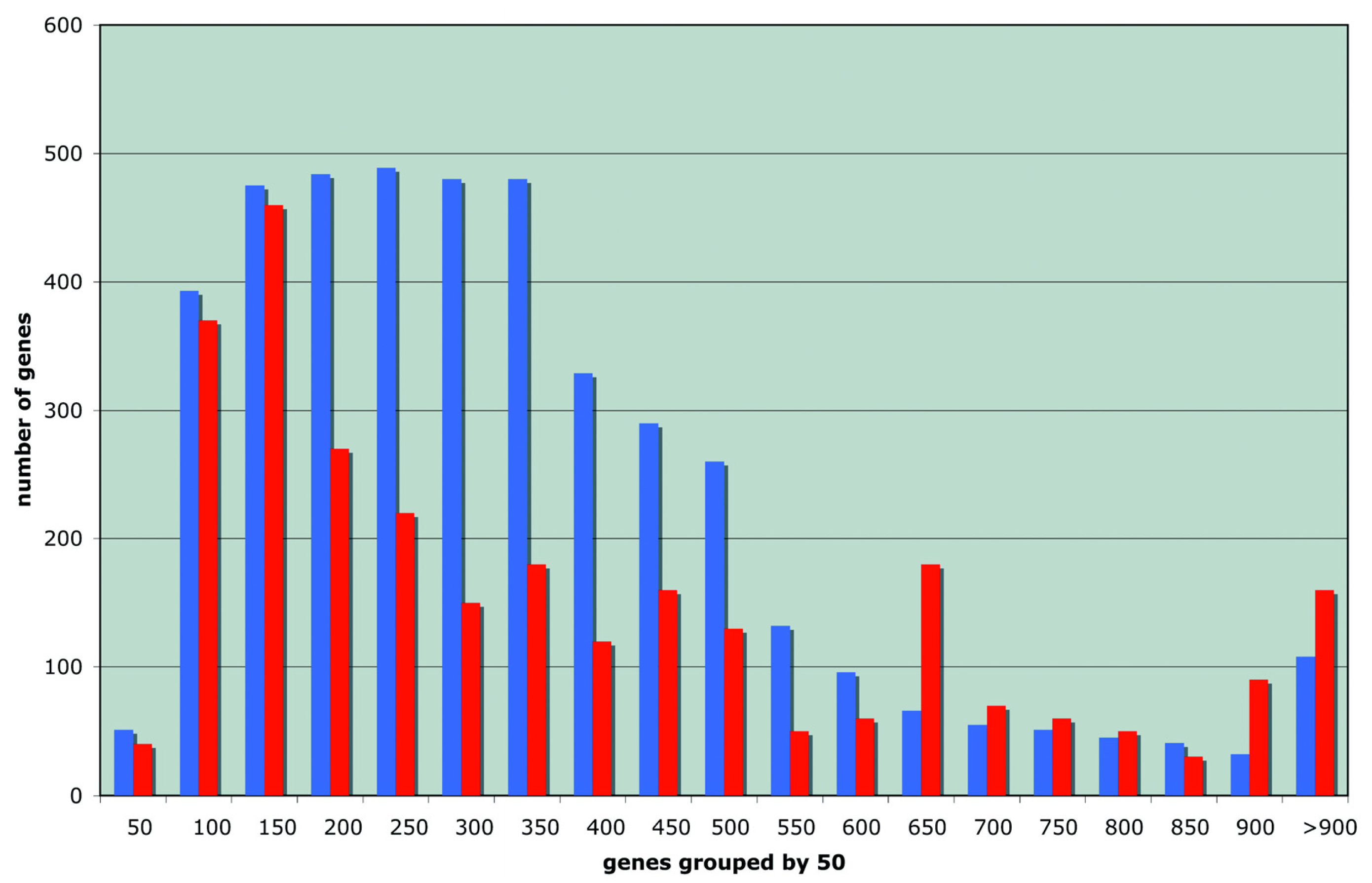
The distribution of the length of the proteins important for stationary survival is compared in Figure 1 to that of all proteins in the E. coli proteome: it displays a remarkable bias both for short and long proteins.
While the presence of short proteins is easy to understand (there is inherent robustness in being short as discussed by Galileo Galilei in his Dialogo sopra i due massimi sistemi del mondo) we need to account for the long ones. We will therefore be looking for unusual properties that could be a signature of antifragility rather than robustness. We explore now whether this unusual length distribution might suggest a direction to follow.
3. Processes Permitting Stress Management
Deleterious effects are often perceived as proportional to the amplitude of their causes. However, most biological systems possess some kind of buffering capacity. Provided a negative effect, a stress for example, is not too pronounced, the system reacts by preserving its homeostasis, displaying no negative change. Some effects, however, are cumulative: exceeding the buffering capacity of the system beyond a certain threshold results in accelerated weathering. In this challenging situation the cell has two choices: either its components will steadily accumulate malfunction, resulting in the error catastrophe [44], or the cell will find a solution where, provided that there are enough representatives of a given particular entity, variants will emerge, some of them ready to provide a relief to the stress induced over time.
This parallels antifragility, as proposed by Taleb to create specific detailed scenarios of financial risk management [14]. The idea behind this reflection is that when a collection of entities is exposed to serious challenges, it may be possible to obtain a positive overall outcome. Within the collection, one of the entities might fare extremely well, compensating for the collapse of some of the others and even do much better than the bulk, had it remained unchallenged. For living organisms this could act at the level of the population of organisms, the population of cells, or the population of intracellular molecules. We explore here how antifragility could operate at the molecular level, noting that its implementation has features highly reminiscent of what we call natural selection. Antifragility gathers and utilizes information [13] in the process of allowing some individual entities to stand out from the bulk and thus improving the fate of a population under a challenging situation. The number of obvious candidates for antifragile behavior among biological processes is enormous. We choose here to illustrate one of them, protein ageing, using a few specific structural features.
4. Flexible Molecules as Support for Antifragile Learning
Emil Fisher's lock and key model is still the paradigm of most biochemical studies. The standard view—often presented on colored images displayed on the cover of popular magazines—is that stable folding is essential not only for protein function but also for resistance to degradation. Our knowledge of proteins is significantly limited by the fact that they can be crystallized. Enzymes are perceived as rigid structures that select their substrates and synthesize products with minimal movement. The discovery of allostery changed that picture to some extent, but it still rested on the idea that activity was linked to stable and fairly rigid conformations. Yet, even crystalline proteins have “disordered” regions that do not show up in the final 3D structure, as they are invisible to X-ray diffraction due to their random distribution. Furthemore, a significant number of the proteins in the list of Table 1 are flexible or have significant flexible sections (e.g., RNA polymerase [45,46], and unexpectedly, the short ribosomal proteins [47]).
Flexibility has long been assumed in the processes of “induced fit” [48]. It reflects the selective stabilization of a conformation belonging to the spontaneous thermal fluctuations of the region of interest, leading to a final form after recruitment of proper short-range interactions (hydrogen bonds, dipolar and van der Waals interactions). Dunker and co-workers shifted the paradigm, showing that as much as one fourth of all proteins displayed a disordered structure [49]. Rather than use this negative qualifier, we here prefer “flexible”, which suggests a specific dynamic order typically relevant to processes of epistasis [50]. Despite some controversy about the role of disordered proteins, it has now been established that flexibility is part of the function of the protein, which must not interact rigidly with its targets and should be degraded once its role has been completed [51]. An example is the recent discovery of the structure and role of RNase Y in B. subtilis [28]. In short, function may require the selection of structures that can only appear after the fact, via proper multimolecular interactions.
4.1. Long Proteins have Biased Physico-Chemical Properties
Long proteins are usually multi-domain proteins connected by flexible links [52]. However, in addition to being more flexible, long proteins display remarkable structural features. Longer proteins can fulfill more easily than short ones the requirements for unfolding and misfolding stability, because they have a higher number of native interactions per residue. They are also relatively enriched in small amino acid residues. Furthermore, there is a correlation between the tendency to misfold and protein hydrophobicity, with long proteins being less hydrophobic than short ones [53]. Interestingly, when hydrophobicity is low, long proteins are more resistant to unfolding and misfolding than short proteins [54]. The collapse of unfolded polypeptides, generally believed to be driven by hydrophobic forces, is an early event in the folding of a protein. Whether hydrogen bonds and side chains play a significant role in the process has been an open question. This has recently been addressed by demonstrating that the backbone displays a considerable organizing role [55]. It seems important therefore that flexibility be a core feature of proteins involved in stationary conditions. Before discussing in more depth the way antifragility may exploit flexibility, we explore some features of flexible regions, which might be a trait selected in proteins that must solve new functional questions while ageing.
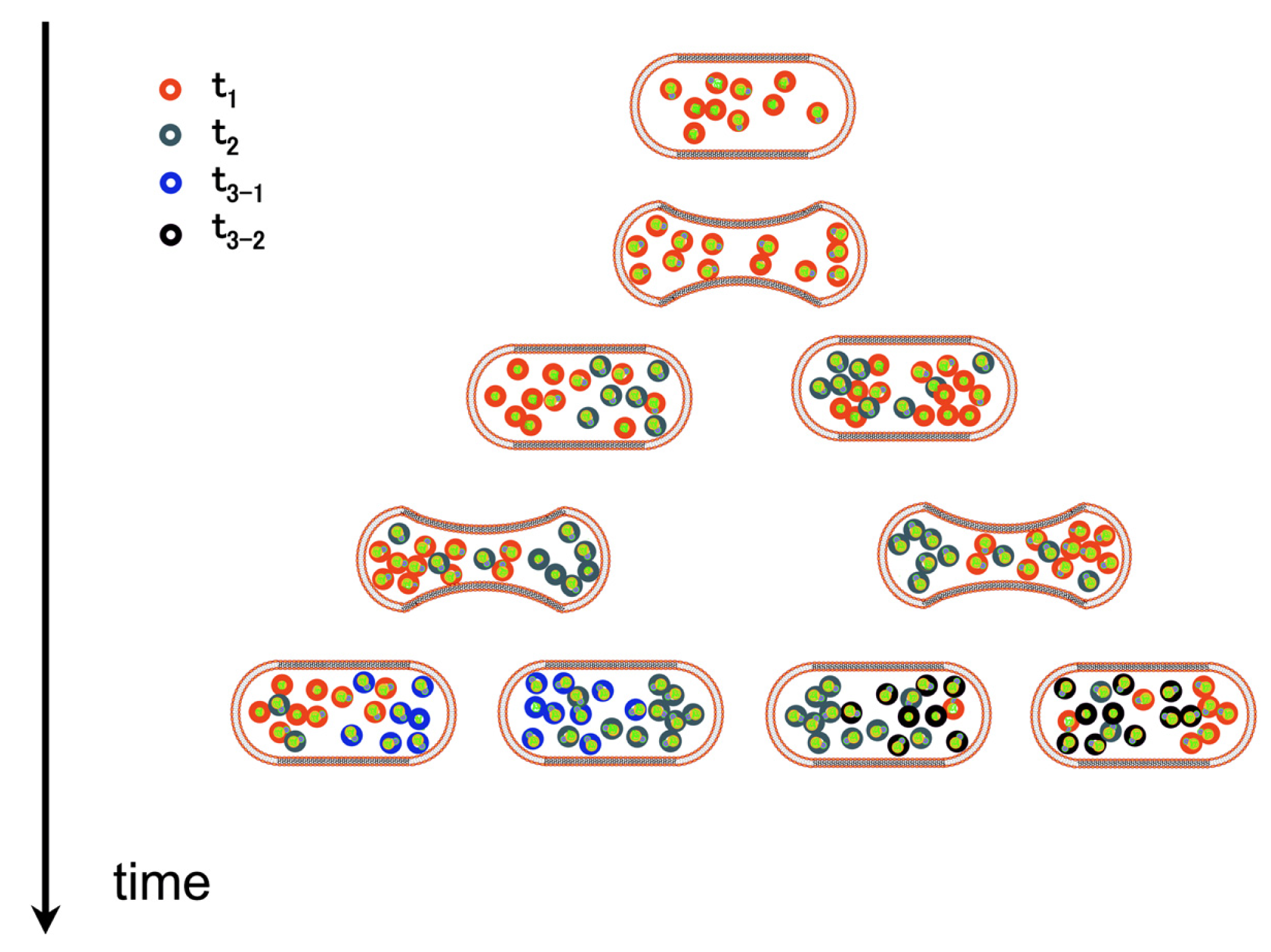
Birth, growth, maturation and senescence are the four ages of all cells. This is true of their components as well. In general the maturation step is ignored: cell's components are viewed as synthesized, used in their final form, then decaying and being either repaired or destroyed. Maturation and possibly functional improvement during ageing is rarely taken into account. Senescence and ageing are treated as equivalent. Yet, quite a few physico-chemical processes suggests that the state of cell components at any time should be seen as actively browsing through a series of ageing states. In fact, notwithstanding apoptosis (which may be a process to reset the system), most cells harbor a mixture of aged and young components, reflecting the overall history of their divisions (Figure 2).

Flexibility plays a considerable role here. We explore the notion that, before the deleterious senescence process is triggered, a process of maturation will create a population of molecules that will undergo different fates. Flexible regions in some cases will make the protein prone to be degraded by proteolytic systems. In other cases, via interaction with specific partners, it will distribute the protein among functional entities. Finally, facing a variety of challenges, flexibility might uncover novel properties of the protein that need time to be implemented. Hence the different individuals in a collection of a given gene product may have different fates, and react differently to similar challenges.
4.2. The Function of Post-Translational Modifications is Disputed
Two processes are generally considered important in the maturation of macromolecules: metabolism-driven modifications and intrinsic instabilities. Post-transcriptional and post-translational modifications show that some metabolism-dependent changes are genetically programmed. At the same time, basal metabolism is required to allow the cell to maintain its activity. Many metabolites are not innocuous. Some have intrinsic reactivity: dicarbonyls (including reducing carbohydrates) react with accessible amino groups resulting in glycation [56] and respiration produces free radicals or reactive oxygen species (ROS) that modify nucleic acids and proteins. The role of ROS is often perceived as negative. However, in addition to being a potential threat to cellular integrity, ROS are also used in a programmed way by the cell, for example in insects, where ROS-triggered protein alteration is required for the formation of the exoskeleton [57]. In the same way, laccase is involved in formation of the spore coat defense in Bacillus subtilis [58]. Superoxide is essential for longevity in Caenorhabditis elegans [59] and there is evidence in mice that several oncogenes actively promote a ROS detoxification program that is required for tumour initiation [60]. This raises questions such as: Is the ROS-dependent modification of a common amino acid, such as methionine oxidation, programmed or accidental [61,62]? Is an oxidized protein a prelude to senescence or a form of the protein that has evolved specifically with the function of regulation, for example?
Unprogrammed modification of macromolecules is often seen as leading to loss of function. This is certainly true of most modifications caused by reactive metabolites such as free radicals, but many modifications are programmed and used as control processes (phosphorylation in particular) or protection processes (methylations or acetylations [63]). Some, such as lysine carbamylation [64] or hypusinylation [65], are even essential for activity. Self-cleavage and formation of a pyruvoyl active center exists in proteins present in the three domains of life [66], inteins are spliced out of ribonucleoside diphosphate reductases [67], reactive centers are made of covalent adducts [68,69]… One also observes more subtle changes in the backbone of macromolecules such as (deoxy)ribose puckering in nucleic acids [70,71], proline puckering [72], isomerisation of aspartate and asparagine with associated deamidation [73], and there is considerable evidence that this could be functional.
Interestingly, the cell's components weather with variable rates (possibly programmed in the very sequence of informational biopolymers [74]), depending on their chemical nature and on their flexibility. This process occurs with a definite half-time, specific of the protein sequence. It allows the protein to display the possibility for time-measurement, i.e., behave as a clock coupled to a structural change. This allows the protein to change its interaction with relevant control mediators, at a pre-set time. Clocks are central devices in information processing. In particular they allow parallel processes the opportunity to synchronize. Accurate timing requires the presence of a stable clock-an unlikely feature of living cells-but cooperation of many inaccurate clocks may result in defining a proper timing [75], possibly to tell cells that they have to start expressing new regulatory pathways or initiate replication. Because this onset of a variety of behaviors which happens after the protein has been synthesized, under conditions when challenges may be taken into account, this process may promote antifragility.
Long proteins such as RNA polymerase subunits have flexible regions and their structure (and sequence) could then change as time elapses. Their length gives them features that make them robust [54], but could they improve over time? Should not we reinvestigate their structure/function in ageing cells? In the same way, repair systems are made of long proteins, and these proteins need to avoid the damages of age because they are essential to prevent premature death [76]. Rather than the cause of modification-derived senescence, it seems worth investigating whether flexibility and length could be the hallmark of antifragility. Indeed, for example, large unstructured hydrophilic flexible regions or proteins provide cellular resistance to dehydration [77].
4.3. Isomerisation of Asparagine and Aspartate Residues Creates Context-Dependent Clocks
To illustrate concretely these points, here is a detailed example of a specific time-dependent post- translational molecular mechanism. Flexibility favors cyclisation of aspartate and asparagine residues into L-succinimide (Figure 3).
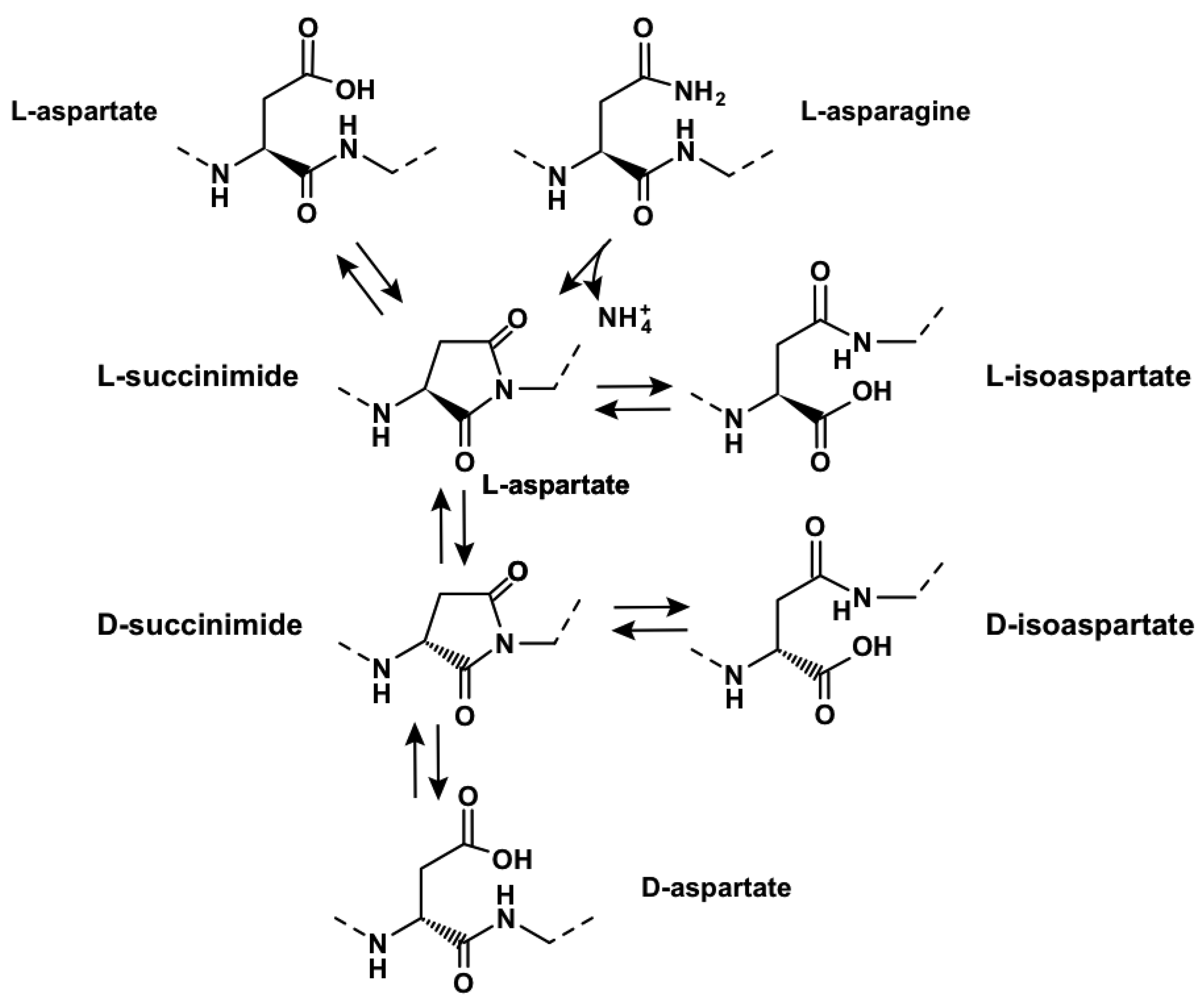
This process is both fast and frequent, in particular at asparagine-glycine (AsnGly) motifs because of the intrinsic flexibility of glycine [73], with an associated deamidation of asparagine. L-succinimide will either hydrolyse into L-isoaspartate spontaneously or after methylation (in 2/3 of the cases), or L-aspartate (in 1/3 of the cases). Subsequently, L-succinimide will isomerise into D-succinimide at a slow rate, and then lead to formation of D-isoaspartate and D-aspartate residues. As a consequence, proteins with flexible regions containing aspartate or asparagine residues will considerably change over time, leading to multiple states, depending on the past history of the protein. Remarkably, the beta subunit of RNA polymerase contains conserved AsnGly motifs. Because of the cost of resynthesising the protein, if destined to become rapidly non-functional after cyclisation-deamidation, these motifs should have been genetically rejected rather than preferred, unless deamidation is of positive biological value. In the same way, aspartates modified by phosphorylation or methylation are also prone to cyclisation. This may lead to loss of activity, creation of novel functional properties, such as regulatory properties or novel catalytic features (reviewed in [73]).
Many other changes occur as time elapses, in particular during the catalytic cycles in which enzymes perform their functions. In some cases, once a protein has been modified (e.g., alkylation of a cysteine residue in the Ada enzyme dealkylating DNA [78]), it must be degraded and resynthesized. However, this is extremely costly and should be limited to some rare situations when it is better to lose energy than control. This cannot be the rule, and inevitable time-dependent changes in macromolecules provide good reasons to suspect that some have been recruited for being functional in the long run, and even to alter their function in a context-related positive direction, allowing them to display antifragile features. This could account for the bias towards long proteins among those that are needed during stationary ageing, the maturation process preceding deleterious senescence.
5. Conclusions
Drawing an image of the environment is a way to foresee the future. This is illustrated in the way living organisms readily cope with changes in their environment, anticipating what it should be like, measuring differences with expectations and adjusting to reality. We propose that antifragility provides a way to allow the viability of cells and organisms in the long run by providing flexibility in structures and processes so that said cells can learn and implement versatile solutions to the problems posed by a variable environment during stationary life.
Extending the concept of induced-fit, we can see antifragile flexible structures and processes as molded on particular ordered structures and processes of the organism. Antifragility could use pre-set properties selected during evolution, such as the retention of asparagine or aspartate residues at specific positions in proteins, and a context-dependent ability to cyclize. This way, the cell could learn and memorize a particular moment in its history, and fixate it so that it could be recalled later on when similar conditions appear. Antifragility would thus be a way to implement a form of epigenetic memory and contribute to the overall mechanisms of network epistasis [50]. In the present context it has properties similar to those used in the market of financial derivatives: the cell develops a diverse population of molecules of processes such that, under challenging conditions, one or more will emerge and improve the cell's fate in a much better way than a uniform population would have [14]. At this point, an aged cell, when facing suddenly a medium where it could reproduce, would initiate a round of replication. We did not discuss here the corresponding required processes and functions, which may underlie the widely spread observation of VBNC [18], but the present view should be seen as a pre-requisite to explore this stage of life that represents up to 99% of the bacteria present in the environment.
This population view, with special properties coming from a few individuals, asks for a way to extract a common underlying order that can be memorized. How could order be identified? Remarkably, there exists a built-in property of large systems of relationships that may answer the question. The cell is constantly in the midst of a huge network of interacting partners. This constitutes an evolving graph. Can we identify some order in the graph? An important feature of large interaction graphs is that they comply with Ramsey's theory, which states that a sufficiently large system, no matter how random, must contain highly organized subsystems [79]. Antifragility is context- dependent. But there is no means for an antifragile structure to know whether the constraints that made it evolve came from the outside or from the inside. Antifragile structures and processes that would change state when some order is reached could memorize a diagram of that particular order. In this way, once time has taken its toll on the organism, a network of interactions would have been created via antifragile modifications of multiple elements, allowing it to trigger re-synthesis of some components that would fit in place in the graph, because the interaction graph itself would play the role of the original situation that enabled its creation.
This behavior is similar to that of the brain, which both records immediate cues in the environment and anticipates its future behavior without knowing whether the stimuli are endogenous or exogenous. This is performed via a set of hierarchical functions that combine bottom-up recording (from the senses to the higher levels of the brain) with top-down feedback used to prepare the sensing apparatus to what might soon appear [80]. An antifragile entity will sense and record “on the fly” memories from the bottom up (for example asparagine cyclisation and deamidation will depend on temperature changes, frequency of the involvement of catalytic activity). And gene expression will adjust, top- down, the answer of the cell as a whole.
When accounting for an immediate reaction to challenge, antifragility thus contributes to fitness. A remarkable property of antifragility is the fact that, by definition, an antifragile entity will react to changes by providing a solution that is not exactly programmed. Hence it does not have a fixed fitness: its response is context and time-dependent. This type of behavior is also the hallmark of tinkering, giving credence to Taleb's reflection.
Acknowledgments
We thank Nassim Taleb for providing us with an unpublished manuscript on antifragility. This work summarises the ongoing discussions of the Stanislas Noria Network, in particular with Eric Fourmentin, Andrew Moore, Bernard Munos and Patrick Tuen Wai Ng. AD wishes to dedicate this work to the memory of Jean Fourmentin, who was a precursor in the onset of genomics and synthetic biology. It has been supported by the FP7 European Union programme Microme KBBE-2007-3-2-08-222886-2 grant.
References
- Taleb, N.N. Antifragility - or - The property of disorder-loving systems. Edge, 2011. Available online: http://www.edge.org/q2011/q11_3.html#taleb (accessed on 24 October 2011). [Google Scholar]
- Perrow, C. Normal Accidents. Living with High-Risk Technologies; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar]
- Taleb, N.N. The Black Swan. The Impact of the Highly Improbable; Random House: New York, NY, USA, 2007. [Google Scholar]
- Archetti, M.; Scheuring, I. Review: Game theory of public goods in one-shot social dilemmas without assortment. J. Theor. Biol. 2011, 1–12, [Epub ahead of print]. [Google Scholar]
- Dasgupta, P. Trust and cooperation among economic agents. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 3301–3309. [Google Scholar]
- Herrera, C. Tinkering with the survival lottery during a public health crisis. J. Med. Philos. 2009, 34, 181–194. [Google Scholar]
- de Vany, A. Hollywood Economics How Extreme Uncertainty Shapes the Film Industry; Routledge: London, UK, 2004. [Google Scholar]
- Jacob, F. The Logic of Life: A History of Heredity; Pantheon Books: New York, NY, USA, 1974. [Google Scholar]
- Negoro, S.; Taniguchi, T.; Kanaoka, M.; Kimura, H.; Okada, H. Plasmid-determined enzymatic degradation of nylon oligomers. J. Bacteriol. 1983, 155, 22–31. [Google Scholar]
- Ohno, S. Birth of a unique enzyme from an alternative reading frame of the preexisted, internally repetitious coding sequence. Proc. Natl. Acad. Sci. USA 1984, 81, 2421–2425. [Google Scholar]
- Yap, Y.L.; Zhang, X.W.; Danchin, A. Relationship of SARS-CoV to other pathogenic RNA viruses explored by tetranucleotide usage profiling. BMC Bioinf. 2003, 4, 43. [Google Scholar]
- Rohde, H.; Qin, J.; Cui, Y.; Li, D.; Loman, N.J.; Hentschke, M.; Chen, W.; Pu, F.; Peng, Y.; Li, J.; et al. Open-Source genomic analysis of Shiga-Toxin-Producing E. coli O104:H4. N. Engl. J. Med. 2011, 365, 718–724. [Google Scholar]
- Binder, P.M.; Danchin, A. Life's demons: Information and order in biology. What subcellular machines gather and process the information necessary to sustain life? EMBO Rep 2011, 12, 495. [Google Scholar]
- Taleb, N.N. A map and simple heuristic to detect fragility, antifragility, and model error. NYU Poly 2011. Working paper series. [Google Scholar]
- Flatt, T.; Min, K.J.; D'Alterio, C.; Villa-Cuesta, E.; Cumbers, J.; Lehmann, R.; Jones, D.L.; Tatar, M. Drosophila germ-line modulation of insulin signaling and lifespan. Proc. Natl. Acad. Sci. USA 2008, 105, 6368–6373. [Google Scholar]
- Lartigue, C.; Glass, J.I.; Alperovich, N.; Pieper, R.; Parmar, P.P.; Hutchison, C.A., 3rd; Smith, H.O.; Venter, J.C. Genome transplantation in bacteria: Changing one species to another. Science 2007, 317, 632–638. [Google Scholar]
- Scott, M.; Hwa, T. Bacterial growth laws and their applications. Curr. Opin. Biotechnol. 2011, 22, 559–565. [Google Scholar]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar]
- Vendeville, A.; Lariviere, D.; Fourmentin, E. An inventory of the bacterial macromolecular components and their spatial organization. FEMS Microbiol. Rev. 2011, 35, 395–414. [Google Scholar]
- Muga, A.; Moro, F. Thermal adaptation of heat shock proteins. Curr. Protein Pept. Sci. 2008, 9, 552–566. [Google Scholar]
- Sippel, K.H.; Venkatakrishnan, B.; Boehlein, S.K.; Sankaran, B.; Quirit, J.G.; Govindasamy, L.; Agbandje-McKenna, M.; Goodison, S.; Rosser, C.J.; McKenna, R. Insights into Mycoplasma genitalium metabolism revealed by the structure of MG289, an extracytoplasmic thiamine binding lipoprotein. Proteins 2011, 79, 528–536. [Google Scholar]
- Lagesen, K.; Ussery, D.W.; Wassenaar, T.M. Genome update: the 1000th genome—A cautionary tale. Microbiology 2010, 156, 603–608. [Google Scholar]
- Fang, G.; Rocha, E.; Danchin, A. How essential are nonessential genes? Mol. Biol. Evol. 2005, 22, 2147–2156. [Google Scholar]
- Ougland, R.; Zhang, C.M.; Liiv, A.; Johansen, R.F.; Seeberg, E.; Hou, YM.; Remme, J.; Falnes, P.O. AlkB restores the biological function of mRNA and tRNA inactivated by chemical methylation. Mol. Cell 2004, 16, 107–116. [Google Scholar]
- Arraiano, C.M.; Andrade, J.M.; Domingues, S.; Guinote, I.B.; Malecki, M.; Matos, R.G.; Moreira, R.N.; Pobre, V.; Reis, F.P.; Saramago, M.; et al. The critical role of RNA processing and degradation in the control of gene expression. FEMS Microbiol. Rev. 2010, 34, 883–923. [Google Scholar]
- Danchin, A. A phylogenetic view of bacterial ribonucleases. Prog. Mol. Biol. Transl. Sci. 2009, 85, 1–41. [Google Scholar]
- Andrade, J.M.; Pobre, V.; Silva, I.J.; Domingues, S.; Arraiano, C.M. The role of 3′-5′ exoribonucleases in RNA degradation. Prog. Mol. Biol. Transl. Sci. 2009, 85, 187–229. [Google Scholar]
- Lehnik-Habrink, M.; Newman, J.; Rothe, F.M.; Solovyova, A.S.; Rodrigues, C.; Herzberg, C.; Commichau, F.M.; Lewis, R.J.; Stulke, J. RNase Y in Bacillus subtilis: A natively disordered protein that is the functional equivalent to RNase E from Escherichia coli. J. Bacteriol. 2011, 193, 5431–5441. [Google Scholar]
- Fang, M.; Zeisberg, W.M.; Condon, C.; Ogryzko, V.; Danchin, A.; Mechold, U. Degradation of nanoRNA is performed by multiple redundant RNases in Bacillus subtilis. Nucleic Acids Res. 2009, 37, 5114–5125. [Google Scholar]
- Dutoit, R.; Dubois, E.; Jacobs, E. Selection systems based on dominant-negative transcription factors for precise genetic engineering. Nucleic Acids Res. 2010, 38, e183. [Google Scholar]
- Dailidiene, D.; Tan, S.; Ogura, K.; Zhang, M.; Lee, A.H.; Severinov, K.; Berg, D.E. Urea sensitization caused by separation of Helicobacter pylori RNA polymerase beta and beta' subunits. Helicobacter 2007, 12, 103–111. [Google Scholar]
- Cohen, S.E.; Walker, GC. The transcription elongation factor NusA is required for stress-induced mutagenesis in Escherichia coli. Curr. Biol. 2010, 20, 80–85. [Google Scholar]
- Srivastava, D.B.; Darst, S.A. Derepression of bacterial transcription-repair coupling factor is associated with a profound conformational change. J. Mol. Biol. 2011, 406, 275–284. [Google Scholar]
- Shimizu, T.; Matsuoka, Y.; Shirasawa, T. Biological significance of isoaspartate and its repair system. Biol. Pharm. Bull. 2005, 28, 1590–1596. [Google Scholar]
- Tarrago, L.; Laugier, E.; Rey, P. Protein-repairing methionine sulfoxide reductases in photosynthetic organisms: Gene organization, reduction mechanisms, and physiological roles. Mol. Plant 2009, 2, 202–217. [Google Scholar]
- Van Schaftingen, E.; Collard, F.; Wiame, E.; Veiga-da-Cunha, M. Enzymatic repair of Amadori products. Amino Acids 2011. [Google Scholar]
- Hopfield, J.J. Kinetic proofreading: A new mechanism for reducing errors in biosynthetic processes requiring high specificity. Proc. Natl Acad. Sci. USA 1974, 71, 4135–4139. [Google Scholar]
- Chatterjee, I.; Neumayer, D.; Herrmann, M. Senescence of staphylococci: Using functional genomics to unravel the roles of ClpC ATPase during late stationary phase. Int. J. Med. Microbiol. 2010, 300, 130–136. [Google Scholar]
- Brotz-Oesterhelt, H.; Beyer, D.; Kroll, H.P.; Endermann, R.; Ladel, C.; Schroeder, W.; Hinzen, B.; Raddatz, S.; Paulsen, H.; Henninger, K.; et al. Dysregulation of bacterial proteolytic machinery by a new class of antibiotics. Nat. Med. 2005, 11, 1082–1087. [Google Scholar]
- Terui, Y.; Tabei, Y.; Akiyama, M.; Higashi, K.; Tomitori, H.; Yamamoto, K.; Ishihama, A.; Igarashi, K.; Kashiwagi, K. Ribosome modulation factor, an important protein for cell viability encoded by the polyamine modulon. J. Biol. Chem. 2010, 285, 28698–28707. [Google Scholar]
- Bubunenko, M.; Baker, T.; Court, D.L. Essentiality of ribosomal and transcription antitermination proteins analyzed by systematic gene replacement in Escherichia coli. J. Bacteriol. 2007, 189, 2844–2853. [Google Scholar]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar]
- Lu, C.; Ding, F.; Chowdhury, A.; Pradhan, V.; Tomsic, J.; Holmes, W.M.; Henkin, T.M.; Ke, A. SAM recognition and conformational switching mechanism in the Bacillus subtilis yitJ S box/SAM-I riboswitch. J. Mol. Biol. 2010, 404, 803–818. [Google Scholar]
- Orgel, L.E. The maintenance of the accuracy of protein synthesis and its relevance to ageing. Proc. Natl. Acad. Sci. USA 1963, 49, 517–521. [Google Scholar]
- Heindl, H.; Greenwell, P.; Weingarten, N.; Kiss, T.; Terstyanszky, G.; Weinzierl, R.O. Cation-pi interactions induce kinking of a molecular hinge in the RNA polymerase bridge-helix domain. Biochem. Soc. Trans. 2011, 39, 31–35. [Google Scholar]
- Weinzierl, R.O. The nucleotide addition cycle of RNA polymerase is controlled by two molecular hinges in the Bridge Helix domain. BMC Biol. 2010, 8, 134. [Google Scholar]
- Timsit, Y.; Acosta, Z.; Allemand, F.; Chiaruttini, C.; Springer, M. The role of disordered ribosomal protein extensions in the early steps of eubacterial 50 S ribosomal subunit assembly. Int. J. Mol. Sci. 2009, 10, 817–834. [Google Scholar]
- Ma, B.; Nussinov, R. Enzyme dynamics point to stepwise conformational selection in catalysis. Curr. Opin. Chem. Biol. 2010, 14, 652–659. [Google Scholar]
- Uversky, VN.; Dunker, A.K. Understanding protein non-folding. Biochim. Biophys. Acta 2010, 1804, 1231–1264. [Google Scholar]
- Lehner, B. Molecular mechanisms of epistasis within and between genes. Trends Genet. 2011, 27, 323–331. [Google Scholar]
- Varedi, K.S.; Ventura, A.C.; Merajver, S.D.; Lin, X.N. Multisite phosphorylation provides an effective and flexible mechanism for switch-like protein degradation. PLoS One 2010, 5, e14029. [Google Scholar]
- Perham, R.N. Swinging arms and swinging domains in multifunctional enzymes: catalytic machines for multistep reactions. Annu. Rev. Biochem. 2000, 69, 961–1004. [Google Scholar]
- Monsellier, E.; Ramazzotti, M.; Taddei, N.; Chiti, F. Aggregation propensity of the human proteome. PLoS Comput. Biol. 2008, 4, e1000199. [Google Scholar]
- Bastolla, U.; Demetrius, L. Stability constraints and protein evolution: the role of chain length, composition and disulfide bonds. Protein Eng. Des. Sel. 2005, 18, 405–415. [Google Scholar]
- Teufel, D.P.; Johnson, C.M.; Lum, J.K.; Neuweiler, H. Backbone-driven collapse in unfolded protein chains. J. Mol. Biol. 2011, 409, 250–262. [Google Scholar]
- Xue, M.; Rabbani, N.; Thornalley, P.J. Glyoxalase in ageing. Semin. Cell Dev. Biol. 2011, 22, 293–301. [Google Scholar]
- Vavricka, C.J.; Christensen, B.M.; Li, J. Melanization in living organisms: A perspective of species evolution. Protein Cell 2011, 1, 830–841. [Google Scholar]
- Fowler, Z.L.; Baron, C.M.; Panepinto, J.C.; Koffas, M.A. Melanization of flavonoids by fungal and bacterial laccases. Yeast 2011, 28, 181–188. [Google Scholar]
- Yang, W.; Hekimi, S. A mitochondrial superoxide signal triggers increased longevity in Caenorhabditis elegans. PLoS Biol. 2010, 8, e1000556. [Google Scholar]
- DeNicola, G.M.; Karreth, F.A.; Humpton, T.J.; Gopinathan, A.; Wei, C.; Frese, K.; Mangal, D.; Yu, K.H.; Yeo, C.J.; Calhoun, E.S.; et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 2011, 475, 106–109. [Google Scholar]
- Furuchi, T.; Sakurako, K.; Katane, M.; Sekine, M.; Homma, H. The role of protein L- isoaspartyl/D-aspartyl O-methyltransferase (PIMT) in intracellular signal transduction. Chem. Biodivers. 2010, 7, 1337–1348. [Google Scholar]
- Lee, B.C.; Gladyshev, VN. The biological significance of methionine sulfoxide stereochemistry. Free Radic. Biol. Med. 2011, 50, 221–227. [Google Scholar]
- Hu, L.I.; Lima, B.P.; Wolfe, A.J. Bacterial protein acetylation: The dawning of a new age. Mol. Microbiol. 2010, 77, 15–21. [Google Scholar]
- Young, P.G.; Smith, C.A.; Metcalf, P.; Baker, E.N. Structures of Mycobacterium tuberculosis folylpolyglutamate synthase complexed with ADP and AMPPCP. Acta Crystallogr. D Biol. Crystallogr. 2008, D64, 745–753. [Google Scholar]
- Bailly, M.; de Crecy-Lagard, V. Predicting the pathway involved in post-translational modification of elongation factor P in a subset of bacterial species. Biol. Direct 2010, 5, 3. [Google Scholar]
- Sekowska, A.; Coppee, J.Y.; Le Caer, J.P.; Martin-Verstraete, I.; Danchin, A. S-adenosylmethionine decarboxylase of Bacillus subtilis is closely related to archaebacterial counterparts. Mol. Microbiol. 2000, 36, 1135–1147. [Google Scholar]
- Anraku, Y.; Satow, Y. Reflections on protein splicing: structures, functions and mechanisms. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 409–421. [Google Scholar]
- Cooley, R.B.; Rhoads, T.W.; Arp, D.J.; Karplus, P.A. A diiron protein autogenerates a valine- phenylalanine cross-link. Science 2011, 332, 929. [Google Scholar]
- Ormo, M.; Cubitt, A.B.; Kallio, K.; Gross, L.A.; Tsien, R.Y.; Remington, S.J. Crystal structure of the Aequorea victoria green fluorescent protein. Science 1996, 273, 1392–1395. [Google Scholar]
- Kahn, T.R.; Fong, K.K.; Jordan, B.; Lek, J.C.; Levitan, R.; Mitchell, P.S.; Wood, C.; Hatcher, M.E. An FTIR investigation of flanking sequence effects on the structure and flexibility of DNA binding sites. Biochemistry 2009, 48, 1315–1321. [Google Scholar]
- Nikolova, E.N.; Kim, E.; Wise, A.A.; O'Brien, P.J.; Andricioaei, I.; Al-Hashimi, H.M. Transient Hoogsteen base pairs in canonical duplex DNA. Nature 2011, 470, 498–502. [Google Scholar]
- Moradi, M.; Babin, V.; Sagui, C.; Roland, C. A statistical analysis of the PPII propensity of amino acid guests in proline-rich peptides. Biophys. J. 2011, 100, 1083–1093. [Google Scholar]
- Robinson, N.; Robinson, A. Molecular Clocks Deamidation of Asparaginyl and Glutaminyl Residues in Peptides and Proteins; Althouse Press: Cave Junction, OR, USA, 2004; p. 443. [Google Scholar]
- Robinson, N.E.; Robinson, A.B. Prediction of protein deamidation rates from primary and threedimensional structure. Proc. Natl. Acad. Sci. USA 2001, 98, 4367–4372. [Google Scholar]
- Mendoza, J.; Challet, E. Brain clocks: from the suprachiasmatic nuclei to a cerebral network. Neuroscientist 2009, 15, 477–488. [Google Scholar]
- Krisko, A.; Radman, M. Protein damage and death by radiation in Escherichia coli and Deinococcus radiodurans. Proc. Natl. Acad. Sci. USA 2010, 107, 14373–14377. [Google Scholar]
- Krisko, A.; Smole, Z.; Debret, G.; Nikolic, N.; Radman, M. Unstructured hydrophilic sequences in prokaryotic proteomes correlate with dehydration tolerance and host association. J. Mol. Biol. 2010, 402, 775–782. [Google Scholar]
- Saget, B.M.; Walker, G.C. The Ada protein acts as both a positive and a negative modulator of Escherichia coli's response to methylating agents. Proc. Natl. Acad. Sci. USA 1994, 91, 9730–9734. [Google Scholar]
- Graham, R.L.; Rothschild, B.L.; Spencer, J.H. Ramsey Theory; John Wiley and Sons: Hoboken, NJ, USA, 1990. [Google Scholar]
- Hawkins, J.; Blakeslee, S. On Intelligence; Times Books, Henry Holt and Company: New York, NY, USA, 2004. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Danchin, A.; Binder, P.M.; Noria, S. Antifragility and Tinkering in Biology (and in Business) Flexibility Provides an Efficient Epigenetic Way to Manage Risk. Genes 2011, 2, 998-1016. https://doi.org/10.3390/genes2040998
Danchin A, Binder PM, Noria S. Antifragility and Tinkering in Biology (and in Business) Flexibility Provides an Efficient Epigenetic Way to Manage Risk. Genes. 2011; 2(4):998-1016. https://doi.org/10.3390/genes2040998
Chicago/Turabian StyleDanchin, Antoine, Philippe M. Binder, and Stanislas Noria. 2011. "Antifragility and Tinkering in Biology (and in Business) Flexibility Provides an Efficient Epigenetic Way to Manage Risk" Genes 2, no. 4: 998-1016. https://doi.org/10.3390/genes2040998
APA StyleDanchin, A., Binder, P. M., & Noria, S. (2011). Antifragility and Tinkering in Biology (and in Business) Flexibility Provides an Efficient Epigenetic Way to Manage Risk. Genes, 2(4), 998-1016. https://doi.org/10.3390/genes2040998