
Development and Validation of a 54K Genome-Wide Liquid SNP Chip Panel by Target Sequencing for Dairy Goat


1
State Key Laboratory of Animal Biotech Breeding, Key Laboratory of Animal Genetics and Breeding of the Ministry of Agriculture, Beijing Key Laboratory for Animal Genetic Improvement, College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
2
Inner Mongolia Academy of Agricultural & Animal Husbandry Sciences, Hohhot 010031, China
*
Author to whom correspondence should be addressed.
Genes 2023, 14(5), 1122; https://doi.org/10.3390/genes14051122
Submission received: 31 March 2023
/
Revised: 18 May 2023
/
Accepted: 19 May 2023
/
Published: 22 May 2023
(This article belongs to the Section Animal Genetics and Genomics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:As an important genotyping platform, SNP chips are essential for implementing genomic selection. In this article, we introduced the development of a liquid SNP chip panel for dairy goats. This panel contains 54,188 SNPs based on genotyping by targeted sequencing (GBTS) technology. The source of SNPs in the panel were from the whole-genome resequencing of 110 dairy goats from three European and two Chinese indigenous dairy goat breeds. The performance of this liquid SNP chip panel was evaluated by genotyping 200 additional goats. Fifteen of them were randomly selected for whole-genome resequencing. The average capture ratio of the panel design loci was 98.41%, and the genotype concordance with resequencing reached 98.02%. We further used this chip panel to conduct genome-wide association studies (GWAS) to detect genetic loci that affect coat color in dairy goats. A single significant association signal for hair color was found on chromosome 8 at 31.52–35.02 Mb. The TYRP1 gene, which is associated with coat color in goats, was identified to be located at this genomic region (chromosome 8: 31,500,048-31,519,064). The emergence of high-precision and low-cost liquid microarrays will improve the analysis of genomics and breeding efficiency of dairy goats.
1. Introduction
Applications of molecular-assisted markers have greatly improved the efficiency and accuracy of livestock selection and breeding [1]. The transition from “empirical breeding” to “precise breeding” with high efficiency not only reduces the shortcoming of conventional breeding, such as the uncertainty of the trait selection, but also significantly improves breeding efficiency and accuracy to develop the new high-yielding and high-quality breeds. Genomic selection (GS) is one of the landmark technologies of molecular breeding. It is characterized by the ability to use genome-wide genetic markers of animals, not just their phenotypes, to guide livestock selection and breeding. Since GS propulsion in 2001 [2], it has revolutionized the field of animal breeding. In the past two decades, genomic selection methods have been successfully applied to breeding practices in many livestock, including cattle [3,4,5], pigs [6,7,8], and chickens [9,10], with satisfactory results.
The accuracy of GS relies on SNP loci covering the whole genome; therefore, the high-throughput variant locus detection technology has been one of bottlenecks in GS. The development of the SNP chip array has advanced the application of GS. More recently, a liquid SNP chip panel based on genotyping by targeted sequencing (GBTS) technology has been developed to reduce the cost further and improve the accuracy of genomic selection. Compared with traditional SNP chips using magnetic beads, liquid-phase chips have many advantages, including low cost and high flexibility [11,12]. It has also been applied to the development of SNP chip arrays for many plant and animal species. For example, the recently developed 40K SNP liquid chip for maize has reduced the genotyping cost to as low as $14 [13].
Dairy goats are widely bred worldwide. Traditional breeding methods for dairy goats rely on manual experience and observation, which have low efficiency, long cycle time, and susceptibility to environmental factors. In contrast, genetic selection is a well-established method for a more objective and accurate assessment of production performance. However, this method has not yet been utilized in Chinese dairy goat breeding due to the small number of phenotype records involved and the lack of commercial SNP chips. Although GoatSNP50 Bead Chip [14] from Illumina Inc. is widely used in GWAS studies for dairy goats [15], new SNP chips with low cost and easy application are needed for Chinese dairy goat genome selection.
This article presents the development and validation of a 54K genome-wide liquid SNP chip panel for targeting dairy goat sequencing. The chip contains SNP-associated production traits from genomic regions. The chip was validated by genotyping 200 dairy goats and comparing their liquid SNP chip panel and WGS data. Developing a 54K genome-wide liquid SNP chip panel using target sequencing is a significant step forward in dairy goat breeding.
2. Materials and Methods
We are committed to ensuring that the ethical principles of animal experimentation are fully observed to ensure that the welfare and rights of animals are fully protected. All experiments were performed under the approval of the Ethics Review Committee of China Agricultural University (permit number: AW12401202-1-32).
2.1. Biological Material, DNA Extraction, and Sequencing
The resequenced animals used in this study consisted of five dairy goat breeds, including three European dairy goat breeds, namely Saanen dairy goats (N = 25), Toggenburg dairy goats (N = 27), and Alpine dairy goats (N = 16), and two Chinese indigenous dairy goat breeds, Guanzhong dairy goats (N = 19) and Laoshan dairy goats (N = 23). Genomic DNA from these 110 goats was extracted from ear punches (preserved in ethanol, -20 degrees Celsius) using Qiagen DNeasy columns (Qiagen, Hilden, Germany) following the manufacturer’s instructions. The extracted DNA was quantified using a Qubit fluorometer, and the quality was assessed using a Nanodrop spectrophotometer. Sequencing libraries were prepared and barcoded using the TruSeq sample preparation kit. The libraries were sequenced on the Illumina HiSeq X Ten platform using the 150 bp paired-end sequencing strategy.
2.2. Trimming, Mapping, and SNP Detection
All WGS reads were checked for sequencing quality using FastQC [16] and trimmed using TrimGalore (version 0.6.5) [17] according to the following parameters: “-g 20 -phred33—stringency 3—length 20-e 0.1.” All cleaned reads were mapped to the goat reference assembly ARS1 (GCF_001704415.1) using BWA-MEM (version 0.7.17) [18] with the default parameter.
To obtain high-quality SNPs for chip development, a Genome Analysis Toolkit (GATK, version 4.2.0.0) [19,20,21] was used to extract the variant according to the following parameters: “QD < 2.0,” “QUAL < 30.0,” “SOR > 3.0,” “MQ < 40.0,” “FS > 60.0,” “MORankSum < −12.5,” and “Read PosRankSum < −8.0.”
After this, PLINK1.9 [22] was used to further extract variants of 29 autosomal chromosomes and 2 contigs, which are unlocalized-scaffold of chromosome X. The filter settings were as follows: SNPs with missing rates (Miss) ≥0.1, and minor allele frequencies (MAF) < 0.05. Finally, a total of 33,716,044 SNPs remained for further analyses.
2.3. Development of the Dairy Goat SNP Panel
To begin with, we conducted selective sweeps according to differences between Chinese and European dairy goat breeds in milk production, milk quality, and other production traits. Selective signal sweep regions were detected according to combinations of three parameters: the XP-CLR approach implemented in the XP-CLR (version 1.1.2) [23], fixation index value (Fst), and nucleotide diversity (π) estimated using vcftools (version 0.1.16). Secondly, QTL loci related to fore udder (FU), feet, and legs (FL), general appearance (GA), rear udder (RU), suspensory ligament (SL), and teats (TE) were obtained from the goat QTLdb database [24] and overlapped with SNPs in the resequencing dataset to get candidate loci. Thirdly, we performed a GWAS analysis based on collected phenotypic information of the 110 resequenced individuals. Association analyses of 305 days of milk yield (MY), milk protein content, somatic cell count, and melatonin synthesis capacity were performed using the MLM in the GEMMA/0.96 [25]. Finally, to ensure uniform SNP density, we filled gaps using loci from the resequencing dataset with a MAF >0.5, and 62,492 candidate SNPs were selected.
After scoring according to the Targeted Capture Sequencing Probe Design System (Compass Biotechnology, Guilin, China), the final 54,188 loci were obtained. A total of 53,990 probes were designed based on the 54K loci, and all probes were 120 bp in length.
2.4. Validation of the 54K Liquid SNP Chip Panel
To get a more comprehensive overview of the panel and SNP performance, 200 additional dairy goat samples were genotyped using the 54K liquid SNP chip panel, of which 15 individuals were randomly selected for 30× whole-genome resequencing. The sequencing process and analysis of the raw data for the liquid SNP chip panel and resequencing are the same as in Section 2.1 and Section 2.2 above.
2.5. Genome-Wide Association Study
To eliminate the contribution of information from relatives, the de-regressive estimated breeding value (DEBV) calculated according to the method proposed by Garrick et al. [26] was used as the response variable in the GWAS in this study. The GWAS was performed with the following single-marker regression model by the GEMMA Software:
where y is the vector of the response variable (DEBVs in this study), m is the vector of SNP marker effects, and a is the vector of residual polygenic effects with a normal distribution , G is the realized relationship matrix constructed with markers and is the additive genetic variance, e is the vector of residual errors with a normal distribution , I being an identity matrix and the residual variance, and X and W are incidence matrices of y related to m and a, respectively.
3. Results
3.1. Design and Development of the 54K Liquid SNP Chip Panel
The sequencing of 110 dairy goats generated 72,083.59M reads, representing an average of 32.45 × coverage per individual (min = 25.3, max = 39.2). After alignment, on average, 99.84% of reads were mapped to the ARS1 assembly. In total, 33,716,044 SNPs were eventually obtained for subsequent chip development. After a series of filtering steps in the material approach (Figure 1A), the final 54,188 loci were identified. 54K loci are evenly distributed in the genome and cover the two contigs LWLT01000021.1 LWLT01000027.1, which were used to represent the unassembled X chromosome of the ARS1 reference genome (Figure 1B). Most of the SNPs are spaced within 100 KB (Figure 1C).
3.2. Genotyping Performance of the Liquid SNP Chip Panel
To validate the genotyping performance of the liquid SNP chip panel, 200 dairy goats were genotyped. The average number of reads obtained from targeted capture sequencing reached 14,927,888, of which the percentage of reads localized to the reference genome was 99.97% on average (Supplementary Materials Table S1). The average number of double-ended reads sequenced to the reference genome and at a distance that matched the length distribution of sequenced fragments (properly mapped ratio) reached 95.61% of all reads, with a small number of individuals below 92% (Figure 2A).
The detection efficiency of the 54K target loci was judged according to the sequencing depth of the loci. The average detection rate was 98.82% for sequencing depths greater than 0 (dp0%), the average detection efficiency decreased to 98.41% for sequencing depths greater than 5 (dp5%), and the average detection efficiency was 97.22% for sequencing depths of 20 (dp20%) as the threshold (Figure 2C, Supplementary Materials Table S2). Analysis of MAF values for captured SNPs showed that 97.04% had MAF values >0.05 (Figure 2B).
As a characteristic of GBTS, the flanking region of the target locus has a high sequencing depth and can capture variants beyond the designed target locus. In this experiment, a total of 100K high-quality SNPs were detected, which was twice as many as the target loci. Meanwhile, the results showed that the average sequencing depth of the target loci reached 91X, and the two flanking loci still had 60X and 26X sequencing depths at distances of 100 and 250 bp, respectively (Figure 2D, Supplementary Materials Table S3).
3.3. Comparison of Chip Panel and Resequencing Results
To investigate the accuracy of the liquid SNP chip panel detection effect, we randomly selected 15 individuals for simultaneous liquid SNP chip panel detection and whole-genome resequencing at a sequencing depth of 30X. It was shown that 98.02% (Figure 3A) and 97.78% (Figure 3C) of 54K target SNPs and a total of 100K SNP are the same as the WGS result. After further analyzing the loci with inconsistent genotypes, the causes of inconsistent loci were classified as follows: “GBTS miss” as no SNP was detected by GBTS, while SNP variants were detected in WGS (GBTS miss), on the opposite “WGS miss.” “GBTS hom” as only one allele was detected on GBTS (monomorphic genotype), while two alleles were detected in WGS (polymorphic genotype), on the opposite “WGS hom.” It was found that among the 54K SNP loci, 96.73% of the loci generated errors due to GBTS (GBTS miss+ GBTS hom) (Figure 3B). Among the total 100K SNP loci, the errors due to GBTS were 85% (GBTS miss+ GBTS hom) (Figure 3D). Among the 54K SNP loci, the “GBTS miss” was over 50%, while among the 100K SNP loci, “GBTS hom” dominated, accounting for 62.58%.
3.4. High-Resolution GWAS for Coat Color
To evaluate the effect of the liquid SNP chip panel, we conducted GWAS to detect genetic loci that affect coat color in dairy goats. In our sequenced population, the Toggenburg dairy goat had a signature brown pattern, while the Saanen dairy goat was pure white. We used the 54K SNP and 100K SNP datasets for detection (Figure 4). GWAS for this trait detected 7 and 23 associated SNPs with genome-wide significance. A significant association signal was found on chromosome 8 at 31.52–35.02 Mb. The TYRP1 gene is located in this genomic region (chromosome 8: 31,500,048–31,519,064).
4. Discussion
Using reliable and low-cost genotyping strategies is essential in animal breeding, including dairy goats. In this study, using resequencing data from Chinese and European dairy goat breeds, we developed a dairy goat liquid SNP chip panel. The main purpose is to set the scope using this chip, mainly for Chinese dairy goat breeding. In the process of SNP selection, trait relatedness was considered an important indicator. The specific traits covered by the liquid SNP chip panel included milk yield, fat yield, protein yield, fore udder, feet and legs, general appearance, rear udder, suspensory ligament, and teats. In addition, SNPs associated with melatonin synthesis capacity were also included. This is because in our previous work, melatonin synthesis capacity was found to play a vital role in milk-producing livestock and can be a critical selection indicator [27,28,29].
This 54K genome-wide liquid SNP chip panel is based on GBTS. After testing and verification, the designed capture efficiency of 54K SNPs can reach 98.41%, and the consistency of capture loci and resequencing can also reach 98.02%. Among other breeding chips based on targeted capture sequencing, the capture efficiency of the current chip can be considered highly satisfactory [30,31,32]. In addition, the GBTS could obtain variant SNP other than the target capture SNP. In previous publications, this was referred to as the “multiple single-nucleotide polymorphism (mSNP) approach” [33,34]. We also captured over twice as many high-quality SNP variants at the designed loci in the liquid SNP chip panel. Compared to other reports, we detected relatively fewer SNPs. This may be related to the fact that the conditions for screening consistent with resequencing were different in the variant screening process and/or the variable probe length, as well as in the size of our chip detective population. When comparing the resequencing results with the multi-capture SNP, it was found that more than 97% of the SNP were identical, further demonstrating the high quality of mSNPs obtained by targeted capture sequencing.
By performing GWAS analysis on dairy goat hair color using the captured 54K SNP dataset and 100K SNP dataset, respectively, TYRP1, a gene associated with goat and other animals’ coat color [35,36,37,38], was successfully localized at chromosome 8: 31,500,048-31,519,064. Meanwhile, it was found that the Manhattan and QQ plots produced by the 100K dataset were better than the 54K SNP dataset. Due to the difficulty of recording phenotypic data of the experimental population, it is not practical to perform GWAS studies for other traits. However, it is possible to find more associated SNPs if more individual phenotypes can be obtained, especially with twice as many SNPs as target loci. By analyzing the reliability of GBTS for SNP detection, the data showed that the full range of SNP obtained using GBTS is equivalent or even superior to the target SNP. This finding further illustrates the advantages of liquid-phase microarrays based on targeted capture sequencing.
Of course, the liquid SNP chip panel-based GBTS had some shortcomings during the analysis. By comparing liquid SNP chip panel detection to resequencing of genotypically inconsistent SNP, it was found that most errors came from GBTS, where variants were not detected, or heterozygous alleles were considered to be homozygote alleles. The reason for this phenomenon is most likely related to the allele drop-out (ADO) that occurs during the PCR process. This occurs when only one of the two alleles present in a sample is amplified to a detectable level [39]. This phenomenon indicates that GBTS is not as accurate as resequencing.
Compared with SNP chips based on the magnetic bead principle, liquid SNP chip panels based on GBTS have obvious advantages, including low cost and a higher number of detection SNPs than design SNPs. In addition, the quantity of liquid SNP chip panels can be produced according to the required testing quantity, which further reduces costs. However, due to the longer time for data analysis and processing with liquid SNP chip panels, this is considered a disadvantage compared with the bead SNP chip. This disadvantage may have its benefit, because as the detection population expands, the SNPs that are not originally detected as polymorphic may be re-identified, resulting in a richer set of variant SNP datasets.
5. Conclusions
Overall, a low-cost, high-accuracy dairy goat whole-genome liquid chip based on resequencing data was developed. This chip can be widely used in GWAS and GS for large-scale genotyping, offering a new tool for the molecular breeding and genetic improvement of dairy goats.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14051122/s1, Supplementary Methods: Selective signatures associated with phenotypic traits based on SNPs. Table S1: Reads mapping ratios of 200 goats; Table S2: Capture ratios above dp0, dp5, dp10, dp15, and dp20 in 200 goats, respectively; Table S3: Average read depth around the 54K sites for probe design.
Author Contributions
Conceptualization, writing—review and editing, project administration, G.L.; methodology, data curation, writing—original draft, S.G.; data curation, writing—original draft, W.L.; writing—review and editing, H.J.; writing—review and editing, supervision, L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Inner Mongolia Autonomous Region Science and Technology Major Special Project, grant number (2021ZD0023-1); Biological Breeding Project-Topic4, grant number (2022ZD0401404); National Key Research and Development Program, grant number (2021YFD1300903).
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Review Committee of China Agricultural University for studies involving animals. (Permit number: AW12401202-1-32).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to the experiment based on these data is not over yet.
Acknowledgments
The authors would like to express their sincere gratitude to the following organizations for their generous support and assistance throughout the project: Inner Mongolia Shengjian Biotechnology Co., Ltd., Inner Mongolia Huayile Livestock Technology Co., Ltd., Inner Mongolia Grassland Hongbao Food Co., Ltd., and Qingdao Samuels Biotechnology Co., Ltd. Your valuable contributions have played an instrumental role in the successful completion of this research endeavor.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wallace, J.G.; Rodgers-Melnick, E.; Buckler, E.S. On the Road to Breeding 4.0: Unraveling the Good, the Bad, and the Boring of Crop Quantitative Genomics. Annu. Rev. Genet. 2016, 52, 421–444. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Guillenea, A.; Su, G.; Lund, M.S.; Karaman, E. Genomic prediction in Nordic Red dairy cattle considering breed origin of alleles. J. Dairy Sci. 2022, 105, 2426–2438. [Google Scholar] [CrossRef] [PubMed]
- Bonifazi, R.; Calus, M.P.L.; Napel, J.T.; Veerkamp, R.F.; Michenet, A.; Savoia, S.; Cromie, A.; Vandenplas, J. International single-step SNPBLUP beef cattle evaluations for Limousin weaning weight. Genet. Sel. Evol. 2022, 54, 57. [Google Scholar] [CrossRef]
- Bolormaa, S.; MacLeod, I.M.; Khansefid, M.; Marett, L.C.; Wales, W.J.; Miglior, F.; Baes, C.F.; Schenkel, F.S.; Connor, E.E.; Manzanilla-Pech, C.I.V.; et al. Sharing of either phenotypes or genetic variants can increase the accuracy of genomic prediction of feed efficiency. Genet. Sel. Evol. 2022, 54, 60. [Google Scholar] [CrossRef]
- Ros-Freixedes, R.; Johnsson, M.; Whalen, A.; Chen, C.-Y.; Valente, B.D.; Herring, W.O.; Gorjanc, G.; Hickey, J.M. Genomic prediction with whole-genome sequence data in intensely selected pig lines. Genet. Sel. Evol. 2022, 54, 65. [Google Scholar] [CrossRef]
- Ye, H.; Zhang, Z.; Ren, D.; Cai, X.; Zhu, Q.; Ding, X.; Zhang, H.; Zhang, Z.; Li, J. Genomic Prediction Using LD-Based Haplotypes in Combined Pig Populations. Front. Genet. 2022, 13, 843300. [Google Scholar] [CrossRef]
- Jibrila, I.; Napel, J.; Vandenplas, J.; Bergsma, R.; Veerkamp, R.F.; Calus, M.P.L. Impact of genomic preselection on subsequent ssGBLUP evaluation of preselected animals for scarcely recorded feed intake in pigs. J. Anim. Breed. Genet. 2023, 140, 253–263. [Google Scholar] [CrossRef]
- Li, X.; Nie, C.; Liu, Y.; Chen, Y.; Lv, X.; Wang, L.; Zhang, J.; Yang, W.; Li, K.; Zheng, C.; et al. The Genetic Architecture of Early Body Temperature and Its Correlation With Salmonella Pullorum Resistance in Three Chicken Breeds. Front. Genet. 2020, 10, 1287. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Ben-Jemaa, S.; Perini, F.; Cendron, F.; Biscarini, F.; Lasagna, E.; Penasa, M.; Cassandro, M. Genome-wide mapping of signatures of selection using a high-density array identified candidate genes for growth traits and local adaptation in chickens. Genet. Sel. Evol. 2023, 55, 20. [Google Scholar] [CrossRef]
- Mamanova, L.; Coffey, A.J.; Scott, C.E.; Kozarewa, I.; Turner, E.H.; Kumar, A.; Howard, E.; Shendure, J.; Turner, D.J. Target-enrichment strategies for next-generation sequencing. Nat. Methods 2010, 7, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Sekine, D.; Oku, S.; Nunome, T.; Hirakawa, H.; Tsujimura, M.; Terachi, T.; Toyoda, A.; Shigyo, M.; Sato, S.; Tsukazaki, H. Development of a genome-wide marker design workflow for onions and its application in target amplicon sequencing-based genotyping. Dna. Res. Int. J. Rapid Publ. Rep. Genes Genomes 2022, 29, dsac020. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yang, Q.; Huang, F.; Zheng, H.; Sang, Z.; Xu, Y.; Zhang, C.; Wu, K.; Tao, J.; Prasanna, B.M.; et al. Development of high-resolution multiple-SNP arrays for genetic analyses and molecular breeding through genotyping by target sequencing and liquid chip. Plant Commun. 2021, 2, 100230. [Google Scholar] [CrossRef] [PubMed]
- Tosser-Klopp, G.; Bardou, P.; Bouchez, O.; Cabau, C.; Crooijmans, R.; Dong, Y.; Donnadieu-Tonon, C.; Eggen, A.; Heuven, H.C.M.; Jamli, S.; et al. Design and Characterization of a 52K SNP Chip for Goats. PLoS ONE 2014, 9, e86227. [Google Scholar] [CrossRef]
- Massender, E.; Oliveira, H.R.; Brito, L.F.; Maignel, L.; Jafarikia, M.; Baes, C.F.; Sullivan, B.; Schenkel, F.S. Genome-wide association study for milk production and conformation traits in Canadian Alpine and Saanen dairy goats. J. Dairy Sci. 2023, 106, 1168–1189. [Google Scholar] [CrossRef]
- Andrews, S.; Krueger, F.; Segonds-Pichon, A.; Biggins, L.; Krueger, C.; Wingett, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://qubeshub.org/resources/fastqc (accessed on 10 January 2022.).
- Krueger, F.; James, F.; Ewels, P.; Afyounian, E.; Schuster-Boeckler, B. FelixKrueger/TrimGalore: V0.6.7-DOI via Zenodo (0.6.7). Zenodo 2021. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to High-Confidence Variant Calls: The Genome Analysis Toolkit Best Practices Pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Patterson, N.; Reich, D. Population differentiation as a test for selective sweeps. Genome Res. 2010, 20, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-L.; Park, C.A.; Reecy, J.M. Bringing the Animal QTLdb and CorrDB into the future: Meeting new challenges and providing updated services. Nucleic Acids Res. 2021, 50, D956–D961. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Garrick, D.J.; Taylor, J.F.; Fernando, R.L. Deregressing estimated breeding values and weighting information for genomic regression analyses. Genet. Sel. Evol. 2009, 41, 55. [Google Scholar] [CrossRef]
- Yang, M.; Shi, J.; Tian, J.; Tao, J.; Chai, M.; Wang, J.; Xu, Z.; Song, Y.; Zhu, K.; Ji, P.; et al. Exogenous melatonin reduces somatic cell count of milk in Holstein cows. Sci. Rep. 2017, 7, 43280. [Google Scholar] [CrossRef]
- Tao, J.; Yang, M.; Wu, H.; Ma, T.; He, C.; Chai, M.; Zhang, X.; Zhang, J.; Ding, F.; Wang, S.; et al. Effects of AANAT overexpression on the inflammatory responses and autophagy activity in the cellular and transgenic animal levels. Autophagy 2018, 14, 1850–1869. [Google Scholar] [CrossRef]
- Wu, H.; Yao, S.; Wang, T.; Wang, J.; Ren, K.; Yang, H.; Ma, W.; Ji, P.; Lu, Y.; Ma, H.; et al. Effects of Melatonin on Dairy Herd Improvement (DHI) of Holstein Cow with High SCS. Molecules 2021, 26, 834. [Google Scholar] [CrossRef]
- Si, Z.; Jin, S.; Li, J.; Han, Z.; Li, Y.; Wu, X.; Ge, Y.; Fang, L.; Zhang, T.; Hu, Y. The design, validation, and utility of the “ZJU CottonSNP40K” liquid chip through genotyping by target sequencing. Ind. Crop. Prod. 2022, 188, 115629. [Google Scholar] [CrossRef]
- Qiao, X.; Su, R.; Wang, Y.; Wang, R.; Yang, T.; Li, X.; Chen, W.; He, S.; Jiang, Y.; Xu, Q.; et al. Genome-wide Target Enrichment-aided Chip Design: A 66 K SNP Chip for Cashmere Goat. Sci. Rep. 2017, 7, 8621. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, H.; Tao, J.; Ren, Y.; Xu, C.; Wu, K.; Zou, C.; Zhang, J.; Xu, Y. Development of multiple SNP marker panels affordable to breeders through genotyping by target sequencing (GBTS) in maize. Mol. Breed. 2019, 39, 37. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Liang, L.; Xiang, J.; Zhang, F.; Zhang, C.; Li, Y.; Liang, J.; Zheng, T.; Zhang, F.; et al. Rice3K56 is a high-quality SNP array for genome-based genetic studies and breeding in rice (Oryza sativa L.). Crop. J. 2023. [Google Scholar] [CrossRef]
- Liu, J.; Peng, W.; Yu, F.; Lin, W.; Shen, Y.; Yu, W.; Gong, S.; Huang, H.; You, W.; Luo, X.; et al. Development and validation of a 40-K multiple-SNP array for Pacific abalone (Haliotis discus hannai). Aquaculture 2022, 558, 738393. [Google Scholar] [CrossRef]
- Alshanbari, F.; Castaneda, C.; Juras, R.; Hillhouse, A.; Mendoza, M.N.; Gutiérrez, G.A.; De León, F.A.P.; Raudsepp, T. Comparative FISH-Mapping of MC1R, ASIP, and TYRP1 in New and Old World Camelids and Association Analysis With Coat Color Phenotypes in the Dromedary (Camelus dromedarius). Front. Genet. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ding, P.; Chen, S.; Zhao, S.; Wang, J.; Lai, S. Analysis of MC1R, MITF, TYR, TYRP1, and MLPH Genes Polymorphism in Four Rabbit Breeds with Different Coat Colors. Animals 2021, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Otto, M.; Ammann, P.; Keller, I.; Drögemüller, C.; Leeb, T. The brown coat colour of Coppernecked goats is associated with a non-synonymous variant at the TYRP1 locus on chromosome. Anim. Genet. 2014, 46, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Liu, X.; Geng, L.; Ma, R.; Li, L.; Li, J.; Zhang, C.; Liu, Z.; Gong, Y.; Li, X. Illumina-sequencing based transcriptome study of coat color phenotypes in domestic goats. Genes Genom. 2017, 39, 817–830. [Google Scholar] [CrossRef]
- Shestak, A.G.; Bukaeva, A.A.; Saber, S.; Zaklyazminskaya, E.V. Allelic Dropout Is a Common Phenomenon That Reduces the Diagnostic Yield of PCR-Based Sequencing of Targeted Gene Panels. Front. Genet. 2021, 12, 620337. [Google Scholar] [CrossRef]
Figure 1.
Design and development of the 54K liquid SNP chip panel. (A) The graphical summary of the pipeline for liquid SNP chip panel design. (B) 54K SNP density of liquid SNP chip panel. (C) Spacing of adjacent SNPs.
Figure 1.
Design and development of the 54K liquid SNP chip panel. (A) The graphical summary of the pipeline for liquid SNP chip panel design. (B) 54K SNP density of liquid SNP chip panel. (C) Spacing of adjacent SNPs.
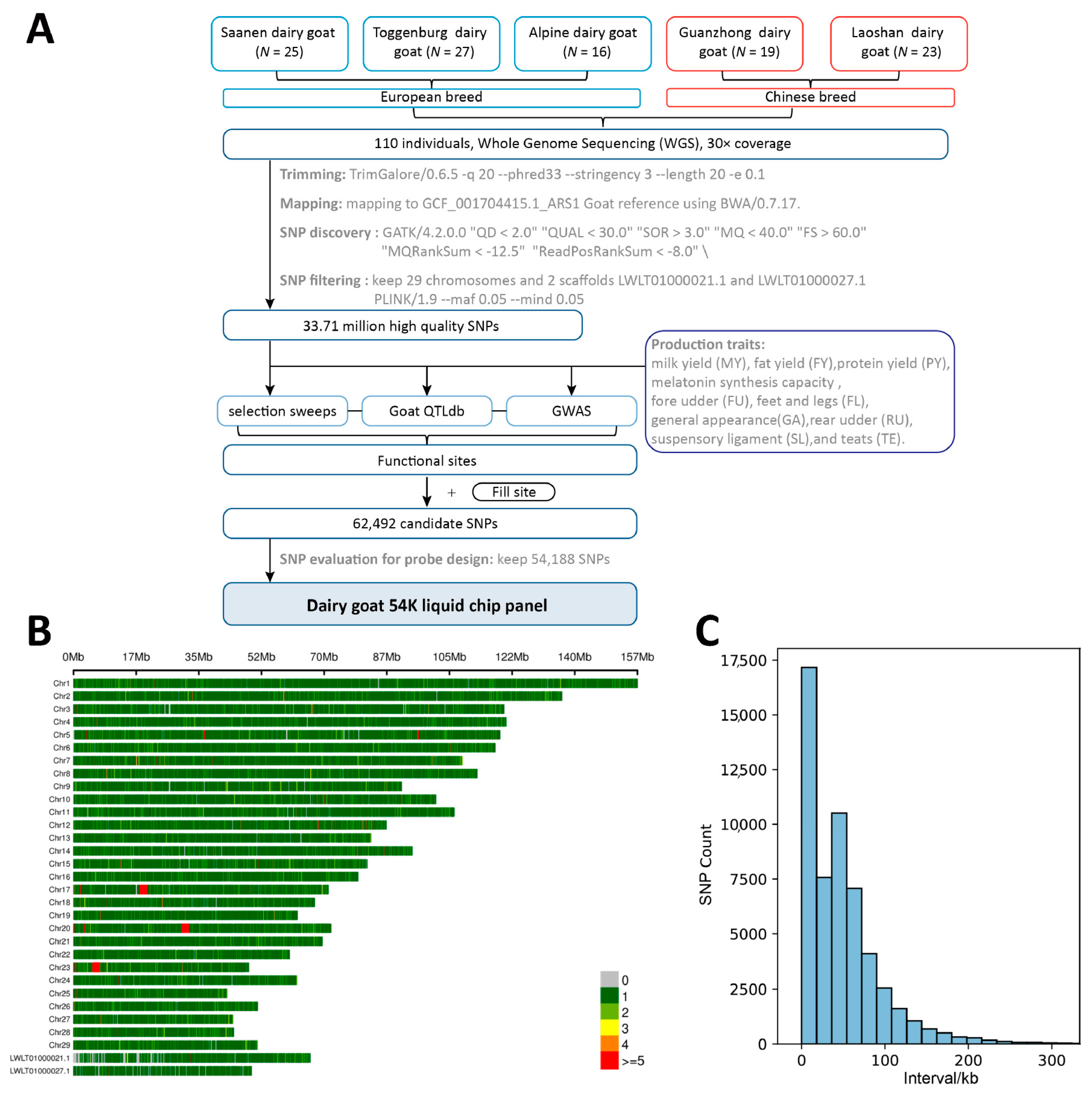
Figure 2.
Genotyping performance of the liquid SNP chip panel: (A) reads properly mapped ratio; (B) minor allele frequency (MAF) of captured 54K SNPs; (C) average detection rate of different sequencing depths; (D) average sequencing depth of the target loci and 500 bp flanking region.
Figure 2.
Genotyping performance of the liquid SNP chip panel: (A) reads properly mapped ratio; (B) minor allele frequency (MAF) of captured 54K SNPs; (C) average detection rate of different sequencing depths; (D) average sequencing depth of the target loci and 500 bp flanking region.
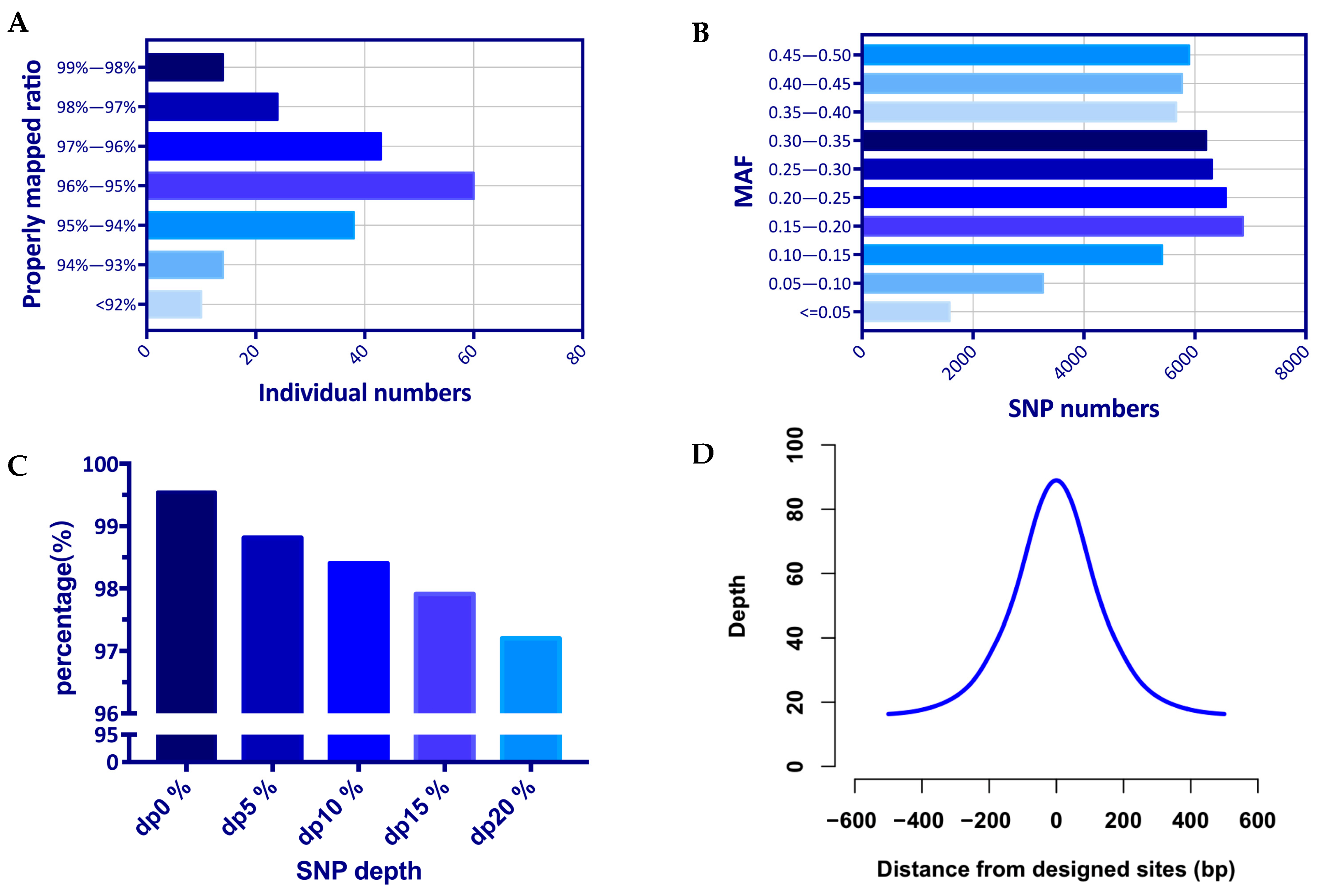
Figure 3.
Comparison of chip panel and resequencing results: (A) 54K SNP dataset with same allele ratio; (B) classification of inconsistent causes in the 54K dataset; (C) 100K SNP dataset with same allele ratio; (D) classification of inconsistent causes in the 100K dataset.
Figure 3.
Comparison of chip panel and resequencing results: (A) 54K SNP dataset with same allele ratio; (B) classification of inconsistent causes in the 54K dataset; (C) 100K SNP dataset with same allele ratio; (D) classification of inconsistent causes in the 100K dataset.
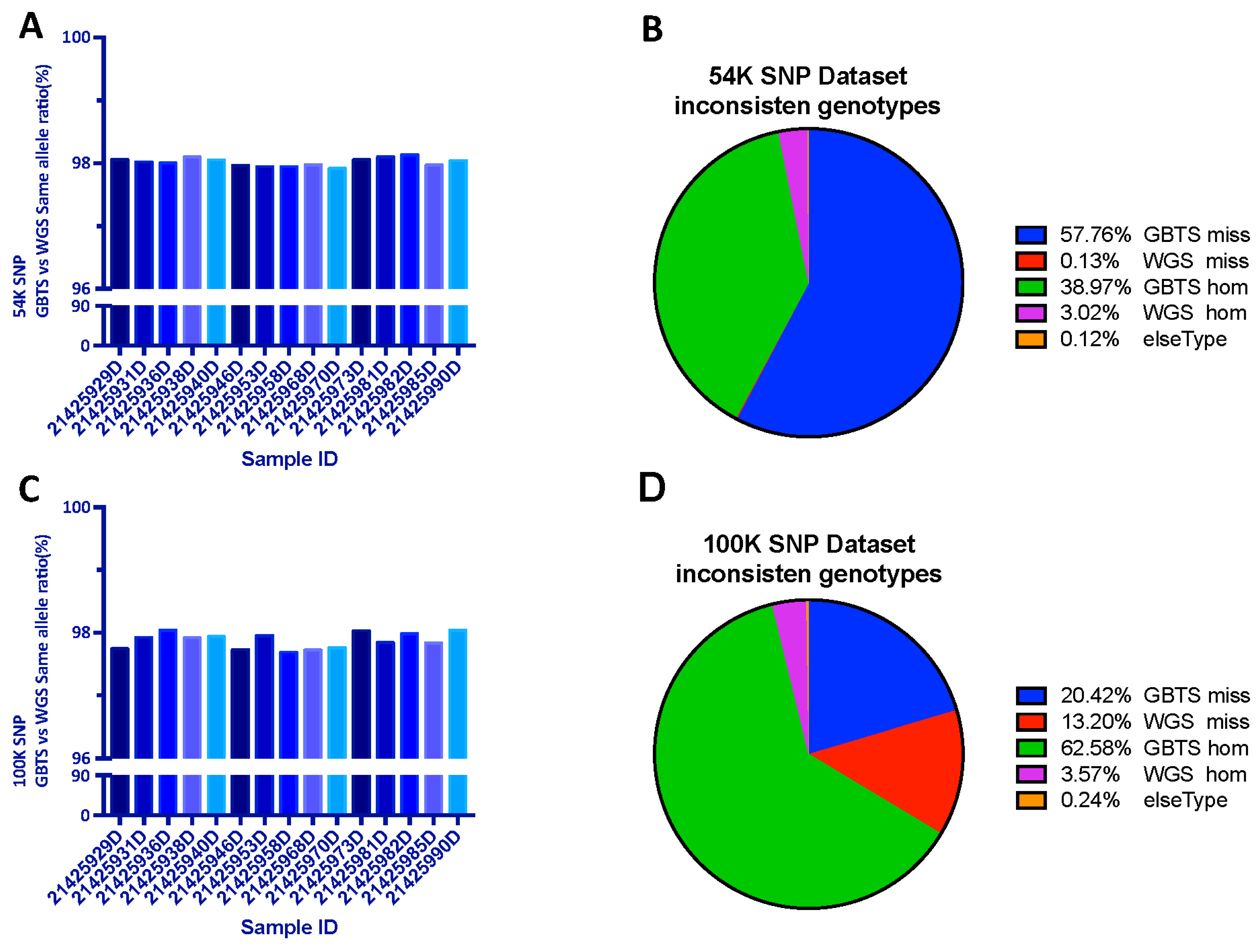
Figure 4.
High-resolution GWAS for coat color: (A,B) Manhattan and quantile-quantile plot of GWAS for coat color using 54K SNP dataset. The red line indicates the threshold for genome-wide significance (p < 5 × 10−8) and the blue line for suggestive associations (p < 1 × 10−5); (C,D) Manhattan and quantile-quantile plot of GWAS for coat color using 100 K SNP dataset.
Figure 4.
High-resolution GWAS for coat color: (A,B) Manhattan and quantile-quantile plot of GWAS for coat color using 54K SNP dataset. The red line indicates the threshold for genome-wide significance (p < 5 × 10−8) and the blue line for suggestive associations (p < 1 × 10−5); (C,D) Manhattan and quantile-quantile plot of GWAS for coat color using 100 K SNP dataset.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guan, S.; Li, W.; Jin, H.; Zhang, L.; Liu, G. Development and Validation of a 54K Genome-Wide Liquid SNP Chip Panel by Target Sequencing for Dairy Goat. Genes 2023, 14, 1122. https://doi.org/10.3390/genes14051122
AMA Style
Guan S, Li W, Jin H, Zhang L, Liu G. Development and Validation of a 54K Genome-Wide Liquid SNP Chip Panel by Target Sequencing for Dairy Goat. Genes. 2023; 14(5):1122. https://doi.org/10.3390/genes14051122
Chicago/Turabian StyleGuan, Shengyu, Weining Li, Hai Jin, Lu Zhang, and Guoshi Liu. 2023. "Development and Validation of a 54K Genome-Wide Liquid SNP Chip Panel by Target Sequencing for Dairy Goat" Genes 14, no. 5: 1122. https://doi.org/10.3390/genes14051122
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.