
The Independent Domestication of Timopheev’s Wheat: Insights from Haplotype Analysis of the Brittle rachis 1 (BTR1-A) Gene

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Phenotyping
2.2. Amplification and Sequencing of the BTR1-A and BTR1-B Gene Regions
2.3. BTR1-A/btr1-A Marker Development
2.4. Sequencing, Phylogenetic, Haplotypic, and Functional Analyses
3. Results
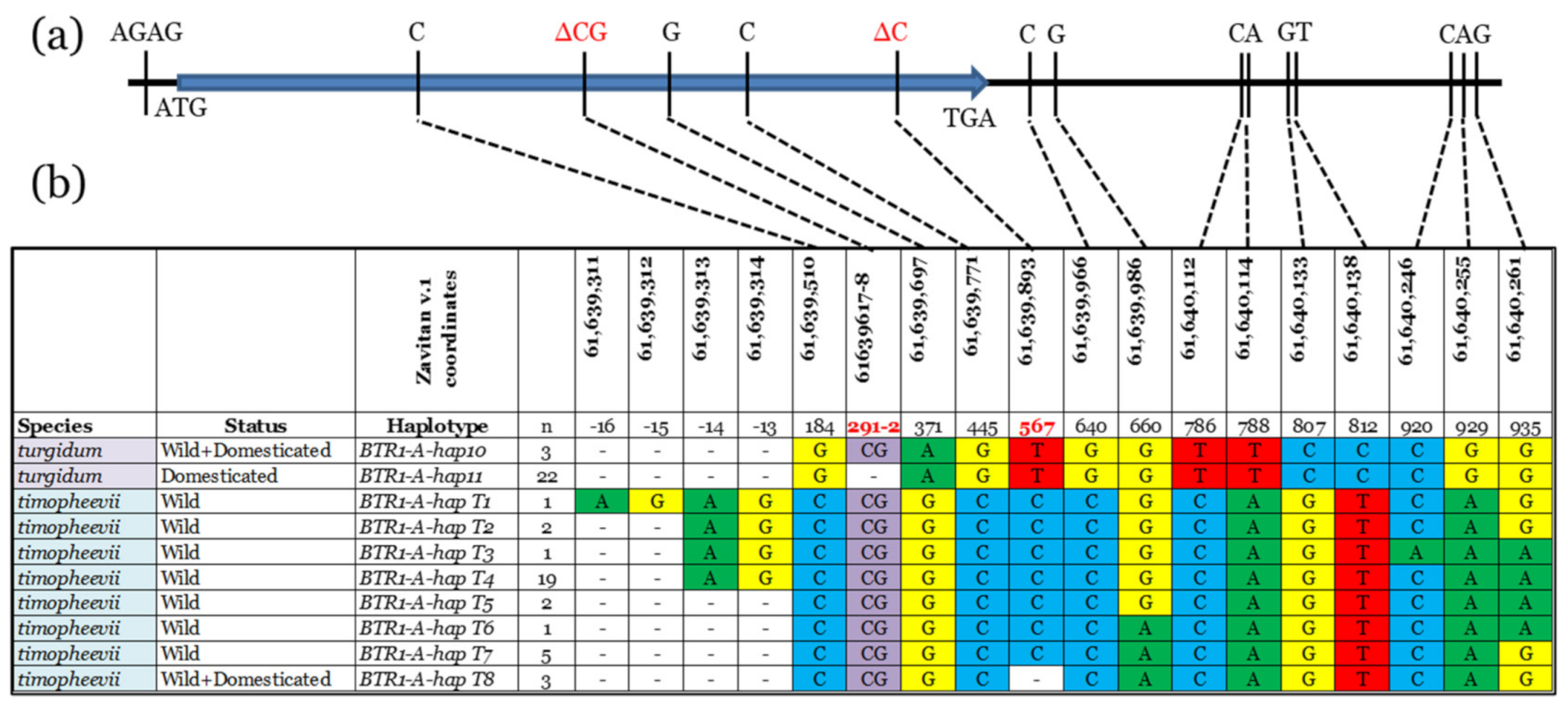
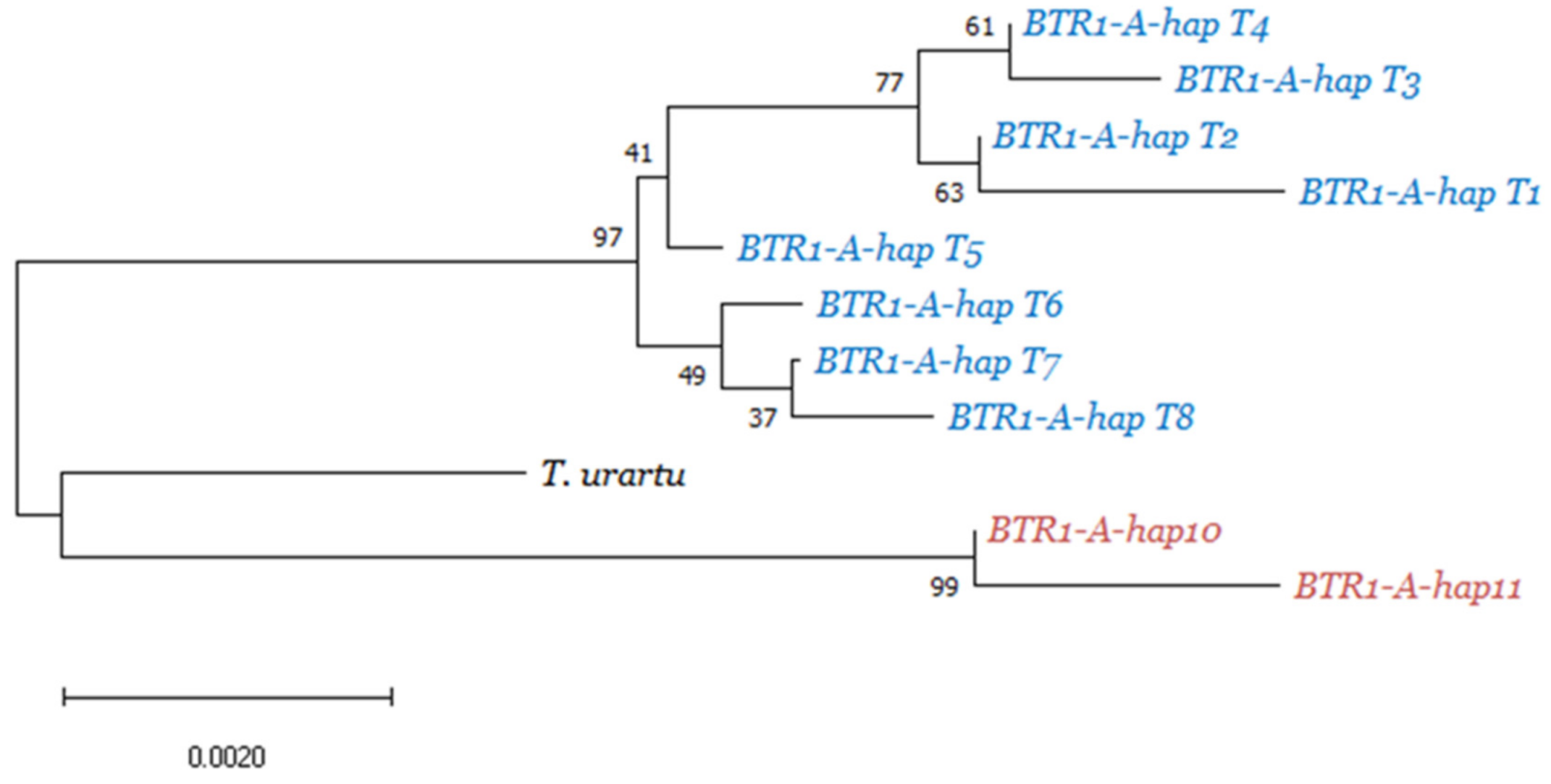
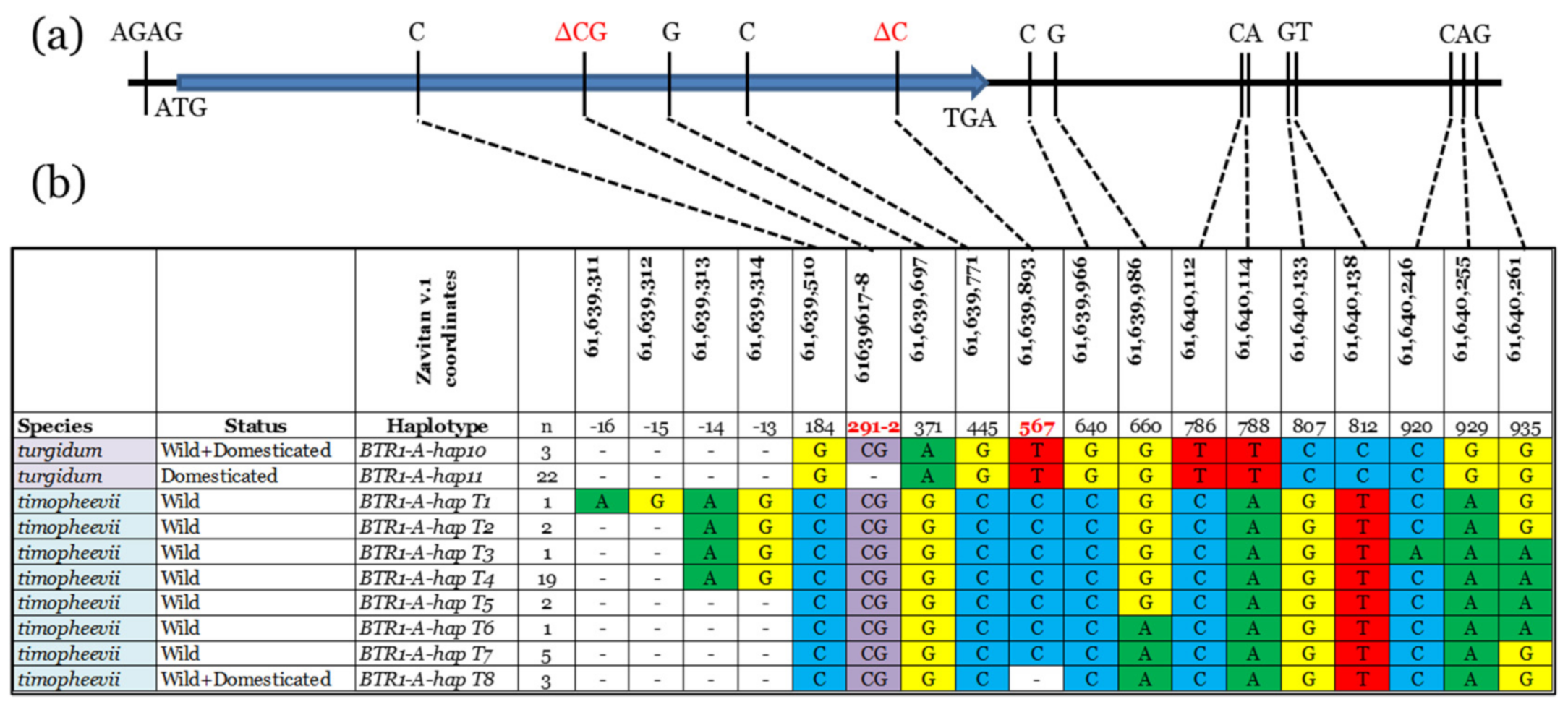
3.1. BTR1-A Haplotype Analysis
3.2. BTR1-B Haplotype Analysis
4. Discussion
4.1. The Triticum Turgidum Lineage
4.2. Triticum Timopheevii Domestication
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubcovsky, J.; Dvorák, J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef] [Green Version]
- Ogihara, Y.; Tsunewaki, K. Diversity and evolution of chloroplast DNA in Triticum and Aegilops as revealed by restriction fragment analysis. Theor. Appl. Genet. 1988, 76, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, J.; di Terlizzi, P.; Zhang, H.B.; Resta, P. The evolution of polyploid wheats: Identification of the A genome donor species. Genome 1993, 36, 21–31. [Google Scholar] [CrossRef]
- Wagenaar, E.B. Studies on the genome constitution of Triticum timopheevi zhuk. ii. The T. timopheevi complex and its origin. Evolution 1966, 20, 150–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, Y. Evolution of polyploid Triticum wheats under cultivation: The role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosulishvili, M.; Bedoshvili, D.; Maisaia, I. A consolidated list of Triticum species and varieties of Georgia to promote repatriation of local diversity from foreign genebanks. Ann. Agrar. Sci. 2017, 15, 61–70. [Google Scholar] [CrossRef]
- Aaronsohn, A. Agricultural and Botanical Explorations in Palestine; Bureau Plant Industry Bulletin, U.S.D.A.; US Government Printing Office: Washington, DC, USA, 1910; pp. 1–63.
- Oliveira, H.R.; Jacocks, L.; Czajkowska, B.I.; Kennedy, S.L.; Brown, T.A. Multiregional origins of the domesticated tetraploid wheats. PLoS ONE 2020, 15, e0227148. [Google Scholar] [CrossRef]
- Mori, N.; Kondo, Y.; Ishii, T.; Kawahara, T.; Valkoun, J.; Nakamura, C. Genetic diversity and origin of timopheevi wheat inferred by chloroplast DNA fingerprinting. Breed. Sci. 2009, 59, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World, 3rd ed.; Oxford University Press: New York, NY, USA, 2000; p. 316. [Google Scholar]
- Jones, G.; Valamoti, S.; Charles, M. Early crop diversity: A “new” glume wheat from northern Greece. Veget. Hist. Archaebot. 2000, 9, 133–146. [Google Scholar] [CrossRef]
- Czajkowska, B.I.; Bogaard, A.; Charles, M.; Jones, G.; Kohler-Schneider, M.; Muller-Bieniek, A.; Brown, T.A. Ancient DNA typing indicates that the “new” glume wheat of early Eurasian agriculture is a cultivated member of the Triticum timopheevii group. J. Archaeol. Sci. 2020, 123, 105258. [Google Scholar] [CrossRef]
- Ulaş, B.; Fiorentino, G. Recent attestations of “new” glume wheat in Turkey: A reassessment of its role in the reconstruction of Neolithic agriculture. Veget. Hist. Archaeobot. 2020, 1–17. [Google Scholar] [CrossRef]
- Maan, S.S. Cytoplasmic and cytogenetic relationships among tetraploid Triticum species. Euphytica 1973, 22, 287–300. [Google Scholar] [CrossRef]
- Maestra, B.; Naranjo, T. Structural chromosome differentiation between Triticum timopheevii and T. turgidum and T. aestivum. Theor. Appl. Genet. 1999, 98, 744–750. [Google Scholar] [CrossRef]
- Naranjo, T. Chromosome structure of durum wheat. Theor. Appl. Genet. 1990, 79, 397–400. [Google Scholar] [CrossRef]
- Jiang, J.; Gill, B.S. Different species-specific chromosome translocations in Triticum timopheevii and T. turgidum support the diphyletic origin of polyploid wheats. Chromosome Res. 1994, 2, 59–64. [Google Scholar] [CrossRef]
- Gill, B.S.; Chen, P.D. Role of cytoplasm-specific introgression in the evolution of the polyploid wheats. Proc. Natl. Acad. Sci. USA 1987, 84, 6800–6804. [Google Scholar] [CrossRef] [Green Version]
- Gill, B.S.; Friebe, B. Cytogenetics, Phylogeny and Evolution of Cultivated Wheats; FAO: Rome, Italy, 2002; Available online: http://www.fao.org/3/y4011e/y4011e07.htm (accessed on 22 January 2021).
- Wang, G.Z.; Miyashua, N.T.; Tsunewaki, K. Plasmon analyses of Triticum (wheat) and Aegilops: PCR-single-strand conformational polymorphism (PCR-SSCP) analyses of organellar DNAs. Proc. Natl. Acad. Sci. USA 1997, 94, 14570–14577. [Google Scholar] [CrossRef] [Green Version]
- Pourkheirandish, M.; Dai, F.; Sakuma, S.; Kanamori, H.; Distelfeld, A.; Willcox, G.; Kawahara, T.; Matsumoto, T.; Kilian, B.; Komatsuda, T. On the Origin of the Non-brittle Rachis Trait of Domesticated Einkorn Wheat. Front. Plant Sci. 2018, 8, 2031. [Google Scholar] [CrossRef] [Green Version]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Nave, M.; Avni, R.; Çakır, E.; Portnoy, V.; Sela, H.; Pourkheirandish, M.; Ozkan, H.; Iago, H.; Komatsuda, T.; Dvorak, J.; et al. Wheat domestication in light of haplotype analyses of the Brittle rachis 1 genes (BTR1-A and BTR1-B). Plant Sci. 2019, 285, 193–199. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R.; et al. Evolution of the Grain Dispersal System in Barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Avni, R.; Nave, M.; Eilam, T.; Sela, H.; Alekperov, C.; Peleg, Z.; Dvorak, J.; Korol, A.; Distelfeld, A. Ultra-dense genetic map of durum wheat × wild emmer wheat developed using the 90K iSelect SNP genotyping assay. Mol. Breed. 2014, 34, 1549–1562. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [Green Version]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Schiller, M.R. The carboxy-terminus, a key regulator of protein function. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 85–102. [Google Scholar] [CrossRef]
- Zeng, X.; Mishina, K.; Jia, J.; Distelfeld, A.; Maughan, P.J.; Kikuchi, S.; Sassa, H.; Komatsuda, T. The brittle rachis trait in species belonging to the Triticeae and its controlling genes Btr1 and Btr2. Front. Plant Sci. 2020, 11, 1000. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nave, M.; Taş, M.; Raupp, J.; Tiwari, V.K.; Ozkan, H.; Poland, J.; Hale, I.; Komatsuda, T.; Distelfeld, A. The Independent Domestication of Timopheev’s Wheat: Insights from Haplotype Analysis of the Brittle rachis 1 (BTR1-A) Gene. Genes 2021, 12, 338. https://doi.org/10.3390/genes12030338
Nave M, Taş M, Raupp J, Tiwari VK, Ozkan H, Poland J, Hale I, Komatsuda T, Distelfeld A. The Independent Domestication of Timopheev’s Wheat: Insights from Haplotype Analysis of the Brittle rachis 1 (BTR1-A) Gene. Genes. 2021; 12(3):338. https://doi.org/10.3390/genes12030338
Chicago/Turabian StyleNave, Moran, Mihriban Taş, John Raupp, Vijay K. Tiwari, Hakan Ozkan, Jesse Poland, Iago Hale, Takao Komatsuda, and Assaf Distelfeld. 2021. "The Independent Domestication of Timopheev’s Wheat: Insights from Haplotype Analysis of the Brittle rachis 1 (BTR1-A) Gene" Genes 12, no. 3: 338. https://doi.org/10.3390/genes12030338