Landscape of Overlapping Gene Expression in The Equine Placenta

 ,
, 
 , ,
, ,
Abstract
1. Introduction
2. Methods
3. Results and Discussion
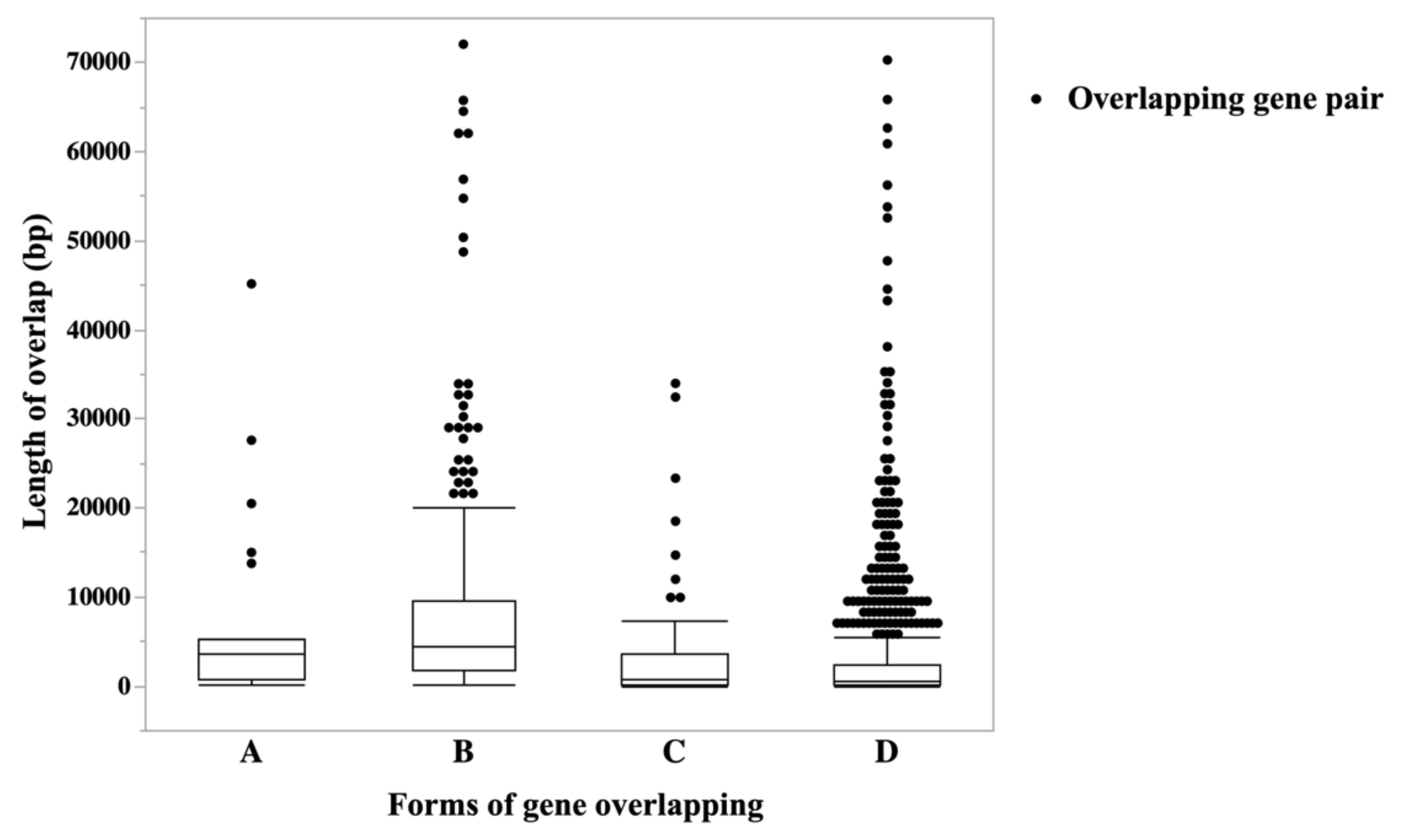
3.1. Identification and Grouping of Overlapping Genes in Placental Transcripts
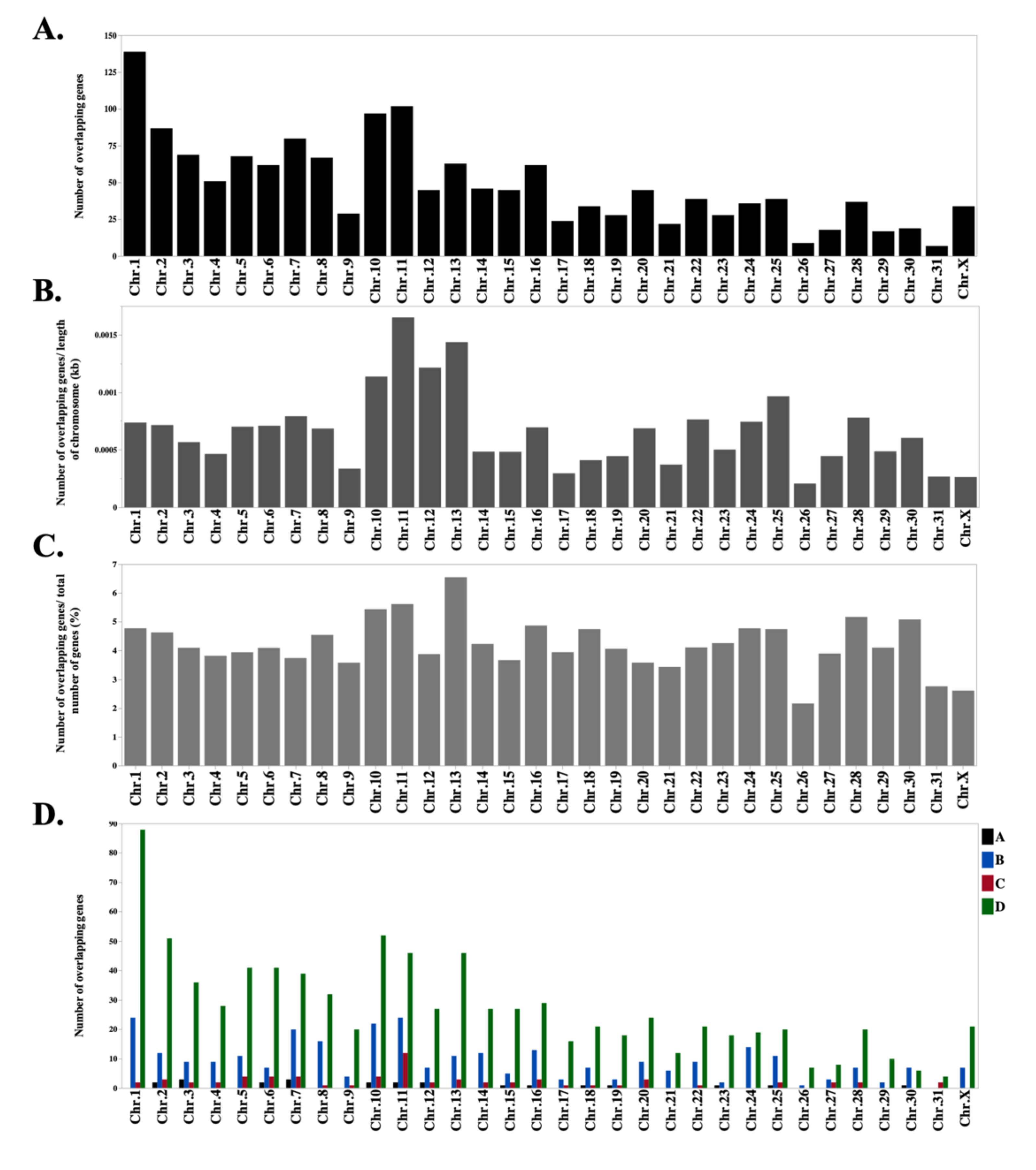
3.2. Overlapping Gene Expression Across The Equine Transcriptome
3.3. Widespread Correlation Between Sense and Antisense Gene Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zackary, I.Z.; Chisholm, S.W. Properties of overlapping genes are conserved across microbial genomes. Genome Res. 2004, 11, 2268–2272. [Google Scholar]
- Barrell, B.G.; Air, G.M.; Hutchison, C.A., III. Overlapping genes in bacteriophage Φx174. Nature 1976, 264, 34. [Google Scholar] [CrossRef] [PubMed]
- Normark, S.; Bergström, S.; Edlund, T.; Grundström, T.; Jaurin, B.; Lindberg, F.P.; Olsson, O. Overlapping genes. Ann. Rev. Gene. 1983, 17, 499–525. [Google Scholar] [CrossRef]
- Lamb, R.A.; Horvath, C.M. Diversity of coding strategies in influenza viruses. Trends Genet. 1991, 7, 261–266. [Google Scholar] [CrossRef]
- Vamsi, V.; Makalowski, W.; Galdzicki, M.; Sood, R.; Makalowska, I. Mammalian overlapping genes: The comparative perspective. Genome Res. 2004, 14, 280–286. [Google Scholar]
- Sanna, C.R.; Li, W.-H.; Zhang, L. Overlapping genes in the human and mouse genomes. BMC Genom. 2008, 9, 169. [Google Scholar] [CrossRef]
- Wood, E.J.; Kwanrutai, C.-I.; Jia, H.; Lipovich, L. Sense-antisense gene pairs: Sequence, transcription, and structure are not conserved between human and mouse. Front. Genet. 2013, 4, 183. [Google Scholar] [CrossRef]
- Nakayama, T.; Asai, S.; Takahashi, Y.; Maekawa, O.; Kasama, Y. Overlapping of genes in the human genome. Int. J. Biomed. Sci. IJBS 2007, 3, 14. [Google Scholar]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 88. [Google Scholar] [CrossRef]
- Guil, S.; Esteller, M. Cis-acting noncoding rnas: Friends and foes. Nat. Struct. Mol. Biol. 2012, 19, 1068. [Google Scholar] [CrossRef]
- Boi, S.; Solda, G.; Tenchini, M.L. Shedding light on the dark side of the genome: Overlapping genes in higher eukaryotes. Curr. Genom. 2004, 5, 509–524. [Google Scholar] [CrossRef]
- Zinad, H.S.; Natasya, I.; Werner, A. Natural antisense transcripts at the interface between host genome and mobile genetic elements. Front. Microbiol. 2017, 8, 2292. [Google Scholar] [CrossRef]
- Murray, S.C.; Haenni, S.; Howe, F.C.; Fischl, H.; Chocian, K.; Nair, A.; Mellor, J. Sense and antisense transcription are associated with distinct chromatin architectures across genes. Nucl. Acids Res. 2015, 43, 7823–7837. [Google Scholar] [CrossRef]
- Morrissy, A.S.; Griffith, M.; Marra, M.A. Extensive relationship between antisense transcription and alternative splicing in the human genome. Genome Res. 2011, 21, 1203–1212. [Google Scholar] [CrossRef]
- Shendure, J.; Church, G.M. Computational discovery of sense-antisense transcription in the human and mouse genomes. Genome Biol. 2002, 3, research0044.1–research0044.14. [Google Scholar] [CrossRef]
- Munroe, S.H.; Zhu, J. Overlapping transcripts, double-stranded RNA and antisense regulation: A genomic perspective. Cell. Mol. Life Sci. CMLS 2006, 63, 2102–2118. [Google Scholar] [CrossRef]
- Lehner, B.; Williams, G.; Campbell, R.D.; Sanderson, C.M. Antisense transcripts in the human genome. Trends Genet. 2002, 18, 63–65. [Google Scholar] [CrossRef]
- Fahey, M.E.; Moore, T.F.; Higgins, D.G. Overlapping antisense transcription in the human genome. Int. J. Genom. 2002, 3, 244–253. [Google Scholar] [CrossRef]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawa, J. Antisense transcription in the mammalian transcriptome. Science 2005, 309, 1564–1566. [Google Scholar]
- Kim, K.; Jutooru, I.; Chadalapaka, G.; Johnson, G.; Frank, J.; Burghardt, R.; Kim, S.; Safe, S. Hotair is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene 2013, 32, 1616. [Google Scholar] [CrossRef]
- Han, Y.; Liu, Y.; Gui, Y.; Cai, Z. Long intergenic non-coding RNA Tug1 Is overexpressed in urothelial carcinoma of the bladder. J. Surg. Oncol. 2013, 107, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.; Boczek, N.J.; Berres, M.V.; Ma, X.; Smith, D.I. lsinct5 is over expressed in breast and ovarian cancer and affects cellular proliferation. RNA Biol. 2011, 8, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.H.; Spieker, T.; Koschmieder, S.; Humberg, J.; Jungen, D.; Bulk, E.; Hascher, A.; Wittmer, D.; Marra, A.; Hillejan, L. The long noncoding malat-1 RNA indicates a poor prognosis in non-small cell lung cancer and induces migration and tumor growth. J. Thorac. Oncol. 2011, 6, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Kalbfleisch, T.S.; Rice, E.S.; DePriest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; Brendan, L.O.; Fiddes, I.T.; Vershinina, A.O.; Saremi, N.F. Improved reference genome for the domestic horse increases assembly contiguity and composition. Commun. Biol. 2018, 1, 197. [Google Scholar] [CrossRef] [PubMed]
- Kalbfleisch, T.S.; Rice, E.S.; De Priest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; Brendan, L.O.; Fiddes, I.T.; Vershinina, A.; Petersen, J.L. Equcab3, an updated reference genome for the domestic horse. bioRxiv, 2018; 306928. [Google Scholar] [CrossRef]
- Sun, M.; Hurst, L.D.; Carmichael, G.G.; Chen, J. Evidence for a preferential targeting of 3′-Utrs by Cis-encoded natural antisense transcripts. Nucl. Acids Res. 2005, 33, 5533–5543. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Miao, S.; Hurst, L.D.; Carmichael, G.G.; Rowley, J.D. Genome-wide analysis of coordinate expression and evolution of human cis-encoded sense-antisense transcripts. Trends Genet. 2005, 21, 326–329. [Google Scholar] [CrossRef]
- Engström, P.G.; Suzuki, H.; Ninomiya, N.; Akalin, A.; Sessa, L.; Lavorgna, G.; Brozzi, A.; Luzi, L.; Tan, S.L.; Yang, L. Complex loci in human and mouse genomes. PLoS GENET. 2006, 2, e47. [Google Scholar] [CrossRef]
- Ito, M.; Sferruzzi-Perri, A.N.; Edwards, C.A.; Adalsteinsson, S.T.; Allen, S.E.; Loo, T.-H.; Kitazawa, M.; Kaneko-Ishino, T.; Ishino, F.; Stewart, C.L. A trans-homologue interaction between reciprocally imprinted mir-127 and RTL1 regulates placenta development. Development 2015, 142, 2425–2430. [Google Scholar] [CrossRef]
- Proudhon, C.; Bourc’his, D. Identification and resolution of artifacts in the interpretation of imprinted gene expression. Brief. Funct. Genom. 2010, 9, 374–384. [Google Scholar] [CrossRef][Green Version]
- Dini, P.; Daels, P.; Loux, S.C.; Esteller-Vico, A.; Carossino, M.; Scoggin, K.E.; Ball, B.A. Kinetics of the chromosome 14 microrna cluster ortholog and its potential role during placental development in the pregnant mare. BMC Genom. 2018, 19, 954. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, V.; Abecasis, G.; Durbin, R. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Cheng, J.; Galili, T.; Bostock, M.; Palmer, J. d3heatmap: Interactive Heat Maps Using ‘htmlwidgets’ and ‘D3.js’. 2018. Available online: https://github.com/rstudio/d3heatmap (accessed on 26 June 2019).
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. Panther: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, J.; Shen, B. Genome-wide analysis of the transcription factor binding preference of human bi-directional promoters and functional annotation of related gene pairs. BMC Systems Biol. 2011, 5, S2. [Google Scholar] [CrossRef]
- Balbin, O.A.; Malik, R.; Dhanasekaran, S.M.; Prensner, J.R.; Cao, X.; Wu, Y.-M.; Robinson, D.; Wang, R.; Chen, G.; Beer, D.G. The landscape of antisense gene expression in human cancers. Genome Res. 2015, 25, 1068–1079. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sense | Antisense | |||||
|---|---|---|---|---|---|---|
| Group | Median (%) | Min (%) | Max (%) | Median (%) | Min (%) | Max (%) |
| A * (n = 23) | 8.4 | 0.01 | 81.8 | 100 | 100 | 100 |
| B (n = 297) | 100 | 100 | 100 | 9.6 | 0.06 | 87.8 |
| C (n = 66) | 4.7 | <0.01 | 98.3 | 5.9 | <0.01 | 93.6 |
| D (n = 875) | 3.1 | <0.01 | 99.8 | 3.5 | <0.01 | 99.9 |
| Biotypes | A * | B | C | D |
|---|---|---|---|---|
| mRNA:mRNA (47%) | 7 | 42 | 29 | 520 |
| mRNA:lnc-RNA (44%) | 12 | 205 | 31 | 309 |
| lnc-RNA:lnc-RNA (4.7%) | 1 | 17 | 4 | 37 |
| mRNA:miRNA (2.6%) | 2 | 23 | - | 1 |
| lnc-RNA:tRNA (0.5%) | - | - | 3 | 4 |
| miRNA:miRNA (0.3%) | - | 4 | - | - |
| lnc-RNA:miRNA (0.3%) | - | 4 | - | - |
| mRNA:miscRNA (0.2%) | - | 1 | - | 2 |
| mRNA:snRNA (0.07%) | - | - | - | 1 |
| lnc-RNA:snRNA (0.07%) | 1 | - | - | - |
| lnc-RNA:miscRNA (0.07%) | - | - | - | 1 |
| Sense | Antisense | Spearman Correlation | Corrected p-Value |
|---|---|---|---|
| NPHP3 | UBA5 | 0.9112 | 0.000001 |
| MSMP | RGP1 | 0.9001 | 0.0057 |
| PSPC1 | MPHOSPH8 | 0.8778 | 0.0042 |
| LOC100059263 | PGRMC1 | 0.8535 | 0.000001 |
| LYRM2 | ANKRD6 | 0.8392 | 0.0367 |
| LOC102147537 | THAP1 | 0.7845 | 0.0009 |
| NDRG2 | SLC39A2 | 0.7728 | 0.0004 |
| GLS2 | SPRYD4 | 0.7155 | 0.0018 |
| EML6 | RTN4 | 0.7105 | 0.002 |
| KIAA0895 | ANLN | 0.7054 | 0.0071 |
| TAF10 | ILK | 0.6946 | 0.0028 |
| LOC111775727 | C11H17orf49 | 0.6933 | 0.0262 |
| LOC111774163 | CNN2 | 0.6813 | 0.0037 |
| TUBGCP6 | SELENOO | 0.6793 | 0.0038 |
| P2RY11 | EIF3G | 0.6681 | 0.0047 |
| LOC106782172 | MTO1 | 0.6673 | 0.0047 |
| TMEM259 | GRIN3B | 0.6500 | 0.0064 |
| LOC100063824 | CCDC51 | 0.6477 | 0.0429 |
| PPP2R3B | LOC102148365 | 0.6429 | 0.0072 |
| RABEP2 | ATP2A1 | 0.6286 | 0.0091 |
| MYO19 | ZNHIT3 | 0.6236 | 0.0098 |
| BCCIP | DHX32 | 0.6035 | 0.0647 |
| ENTPD8 | NOXA1 | 0.5865 | 0.0169 |
| ASRGL1 | LOC111775953 | 0.5862 | 0.0749 |
| MYO15B | LOC111775588 | 0.5622 | 0.0719 |
| CIDEB | NOP9 | 0.5493 | 0.0275 |
| LOC111772592 | ELOVL6 | 0.5475 | 0.0281 |
| LOC106783384 | TNPO2 | 0.5469 | 0.043 |
| NR2C2AP | RFXANK | 0.5271 | 0.0359 |
| TMCO6 | LOC111767785 | 0.5184 | 0.0396 |
| GNE | CLTA | 0.5119 | 0.0427 |
| TNFRSF17 | LOC111767561 | 0.5001 | 0.0576 |
| LOC100053030 | GAS8 | 0.4987 | 0.0493 |
| STAT2 | IL23A | 0.4985 | 0.0494 |
| BAK1 | LOC111769307 | −0.5034 | 0.0468 |
| MYBBP1A | SPNS2 | −0.5172 | 0.0402 |
| AHSA2 | USP34 | −0.5227 | 0.0456 |
| LOC111773182 | LRRC72 | −0.6117 | 0.0455 |
| PCYT1B | PDK3 | −0.6362 | 0.048 |
| IGSF10 | MED12L | −0.6552 | 0.008 |
| CD72 | TESK1 | −0.7735 | 0.0012 |
| LOC111774293 | KRI1 | −0.7918 | 0.0192 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dini, P.; Norris, J.; El-Sheikh Ali, H.; Loux, S.C.; Carossino, M.; Esteller-Vico, A.; Bailey, E.; Kalbfleisch, T.; Daels, P.; Ball, B.A. Landscape of Overlapping Gene Expression in The Equine Placenta. Genes 2019, 10, 503. https://doi.org/10.3390/genes10070503
Dini P, Norris J, El-Sheikh Ali H, Loux SC, Carossino M, Esteller-Vico A, Bailey E, Kalbfleisch T, Daels P, Ball BA. Landscape of Overlapping Gene Expression in The Equine Placenta. Genes. 2019; 10(7):503. https://doi.org/10.3390/genes10070503
Chicago/Turabian StyleDini, Pouya, Jamie Norris, Hossam El-Sheikh Ali, Shavahn C. Loux, Mariano Carossino, Alejandro Esteller-Vico, Ernest Bailey, Theodore Kalbfleisch, Peter Daels, and Barry A. Ball. 2019. "Landscape of Overlapping Gene Expression in The Equine Placenta" Genes 10, no. 7: 503. https://doi.org/10.3390/genes10070503
APA StyleDini, P., Norris, J., El-Sheikh Ali, H., Loux, S. C., Carossino, M., Esteller-Vico, A., Bailey, E., Kalbfleisch, T., Daels, P., & Ball, B. A. (2019). Landscape of Overlapping Gene Expression in The Equine Placenta. Genes, 10(7), 503. https://doi.org/10.3390/genes10070503