Transcriptome Analysis of Diurnal Gene Expression in Chinese Cabbage
by
Jin A. Kim
1,*,†, 
Donghwan Shim
2,†,
Shipra Kumari
1,
Ha-eun Jung
1,
Ki-Hong Jung
3,
Heesu Jeong
4,
Woe-Yeon Kim
5,
Soo In Lee
1 and 
Mi-Jeong Jeong
1 1
Department of Agricultural Biotechnology, National Academy of Agricultural Science, Rural Development Administration, 370, Nongsaengmyeong-ro, Wansan-gu, Jeonju-si 560-500, Korea
2
Department of Forest Bio-resources, National Institute of Forest Science, Suwon 16631, Korea
3
Department of Genetic Engineering and Crop Biotech Institute, Kyung Hee University, Yongin 17104, Korea
4
Department of Plant Science, College of Agriculture and Life Sciences, Seoul National University, San 56-1, Sillim-dong, Gwanak-gu, Seoul 151-744, Korea
5
Division of Applied Life Science (BK21Plus), Institute of Agricultural and Life Science (IALS), Gyeongsang National University, 501 Jinju-daero, Jinju 52828, Korea
*
Author to whom correspondence should be addressed.
†
These authors contribute equally to this work.
Genes 2019, 10(2), 130; https://doi.org/10.3390/genes10020130
Submission received: 16 December 2018
/
Revised: 31 January 2019
/
Accepted: 4 February 2019
/
Published: 11 February 2019
(This article belongs to the Special Issue Genetic Regulation of Circadian Rhythm in Plants)
Abstract
:Plants have developed timing mechanisms that enable them to maintain synchrony with daily environmental events. These timing mechanisms, i.e., circadian clocks, include transcriptional/translational feedback loops that drive 24 h transcriptional rhythms, which underlie oscillations in protein abundance, thus mediating circadian rhythms of behavior, physiology, and metabolism. Circadian clock genes have been investigated in the diploid model plant Arabidopsis thaliana. Crop plants with polyploid genomes—such as Brassica species—have multiple copies of some clock-related genes. Over the last decade, numerous studies have been aimed at identifying and understanding the function of paralogous genes with conserved sequences, or those that diverged during evolution. Brassica rapa’s triplicate genomes retain sequence-level collinearity with Arabidopsis. In this study, we used RNA sequencing (RNAseq) to profile the diurnal transcriptome of Brassica rapa seedlings. We identified candidate paralogs of circadian clock-related genes and assessed their expression levels. These genes and their related traits that modulate the diurnal rhythm of gene expression contribute to the adaptation of crop cultivars. Our findings will contribute to the mechanistic study of circadian clock regulation inherent in polyploidy genome crops, which differ from those of model plants, and thus will be useful for future breeding studies using clock genes.
1. Introduction
Light is a primary energy source for plants—it regulates various stages of plant development and maintains high photosynthetic efficiency [1]. Sessile plants have developed timing mechanisms to maintain synchrony with the Earth’s rotation, and function according to daily environmental events, such as sunrise and sunset [2]. The subsequent transduction of environmental cues regulates, directly or indirectly, diverse developmental processes including photomorphogenesis, floral transition, leaf movements, stomatal conductance, photosynthetic capacity, volatile emissions [3,4,5], and interactions with abiotic and biotic factors [6,7]. These processes seem to have a diurnal rhythm, which occurs during daylight. Circadian rhythm is a biological process that behaves rhythmically during 24 h periods. Under diurnal conditions (light and temperature cycles), organismal genes are expressed due to free running circadian oscillations.
Recently, diurnally expressed genes have been identified in Arabidopsis thaliana [8]; the light signal transduction system—involving photo receptors—has been reported [9,10]; and a computational model of the plant clock system, comprising the main transcriptional feedback loop (loop I) and two additional associated loops (Loops II and III) [11], has been developed. High-throughput RNA sequencing (RNAseq) methods offer dramatic improvements over prior techniques [12,13]. RNA sequencing analysis of circadian transcriptomes derived from wild-type and period-null mutant (per0) Drosophila brains was performed. The results identified novel splicing events and transcripts regulated by the circadian clock. Furthermore, transcriptomic studies performed using several major crop plants including tomato, rice, polar, and lettuce, have demonstrated the expression of diurnal genes under environmental cycles and self-sustaining oscillated circadian genes under constant light or temperature [14,15,16].
The circadian clock coordinates physiological traits such as flowering time, photosynthesis, and growth, as well as internal metabolic and hormonal signals in plants with daily and seasonal environmental changes [17,18,19,20]. This coordination enhances plant fitness, growth, and environmental adaptation [21]. Clock-regulated genes are required for primary metabolism, including CO2 assimilation, starch accumulation/degradation in leaves, and nutrient (lipids and fatty acids) storage in developing seeds [18,20,22,23,24]. Additionally, potato tuberization and abiotic stress response-related secondary metabolic pathways are regulated by clock-related genes [25,26,27,28].
Crop plants that have undergone whole genome duplication events contain multiple copies of genes with or without functional redundancy [29,30,31,32,33,34,35,36,37]. Clock-related genes are conserved among angiosperm evolutionary lineages [33,38,39,40,41,42]. Although accumulation of genome sequences and comparative genome analysis techniques have accelerated the identification of circadian genes in polyploidy genomes, their functional specialization remains unclear. This raises interesting questions regarding how multiple members of gene families contribute to the circadian clock mechanism, and whether the clock model developed for Arabidopsis is applicable to plants with polyploid genomes. Functional studies involving paralogs improve our understanding of the unique physiology of each crop plant and establish a breeding strategy.
The polyploid genus Brassica is comprised of a variety of vegetable crops, such as Chinese cabbage, bok choy, turnip, and broccoletto, as well as oilseed crops, such as turnip rape and sarson, that are of worldwide agricultural importance. It is noteworthy that derived products such as processed oil and kimchi add economic value [43,44]. In the B. rapa genome, after multiple genome duplications and gene losses, circadian clock genes were preferentially retained relative to their neighboring genes [41,45]. Previous studies have described the structures of a pseudo-response regulator (PRR) and circadian clock-associated 1/late elongated hypocotyl (CCA1/LHY) gene families [40,41]. RNA sequencing reads from six tissues of B. rapa accession Chiifu-401-42—the genome of which has been sequenced—indicated widespread transcription of the Chinese cabbage genome [46]. We were interested in the role that the clock genes retained in the B. rapa genome play in the clock network, in addition to the developmental and metabolic processes regulated by them. Here we used RNAseq to profile the circadian transcriptome of leaves derived from wild-type B. rapa. Our findings will contribute to studies on the mechanisms of circadian clock regulation inherent in polyploidy genome crops—which differ from those of model plants—and to plant breeding strategies that require clock genes.
2. Materials and Methods
2.1. Plant Growth and Harvest
Chinese cabbage (B. rapa pekinensis) accession Chiifu-401-42 seeds were sown in individual pots containing soil and grown in a growth chamber (Eyelatron, FLI-301NH, Japan) under a 16/8 h light/dark cycle for 2 weeks, using a cool-white fluorescent lamp. Plants were entrained under a 12/12 h light/dark cycle for 1 week. The plants were watered every three days, and temperatures were maintained at 23 °C until 3–5 true leaves emerged. Aerial parts of three-week old plants were harvested at ZT (zeitgeber Time) 0, 4, 8, 12, 16, 20, and 24, with two biological repeats (Figure 1).
2.2. Library Preparation and RNA Sequencing
Total RNA was isolated from B. rapa leaf tissue using RNeasy Plant Mini Kits (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions. RNA quality was determined using a 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA), and only samples with an RNA integrity number > 8 were used for library preparation. Preparation of each single-end complementary DNA (cDNA) library was conducted according to the TruSeq RNA Sample Preparation Guide (Illumina, San Diego, CA, USA). Complementary DNA libraries sequencing was performed using an Illumina HiSeq2000 sequencer. Base calls were made using CASAVA software (Illumina).
2.3. Transcript Quantification, Differential Expression Analysis, and Gene Annotation
Single-end reads were cleaned by processing with PRINSEQ-lite, v. 0.20.4 software (http://prinseq.sourceforge.net/). Sequences < 50 bp long, with at least one quality score < 10, a mean quality score of < 20, exact duplicates or reverse-complement exact duplicates were filtered out, and sequences with a quality score threshold of 20 were trimmed from both the 5′ and 3′ ends [47].
The clean reads of each sample were aligned to the reference transcriptome (B.rapa_197_transcript_primaryTranscriptOnly.fa of Phytozome V9.0, https://genome.jgi.doe.gov/portal/pages/dynamicOrganismDownload.jsf?organism=Phytozome) using Bowtie software [48]. RSEM v.1.3.0 software (http://deweylab.github.io/RSEM/) was used to generate read counts, and the trimmed mean of M-values (TMM)-normalized fragments per kb of exon per million (FPKM) reads mapped for each transcript [49]. For differential expression analysis, negative binomial dispersion across samples was calculated using the EdgeR v. 3.16.5 software [50]. Genes with more than a twofold change in expression, and a false discovery rate (FDR)-adjusted p-value ≤0.05, were considered to be differentially expressed. Information on gene annotation was supported by the B.rapa_197_annotation profile of Phytozome V9.0 (https://phytozome.jgi.doe.gov).
To identify rhythmically expressed genes, we applied the JTK-CYCLE circadian transcript analysis software. This is a non-parametric statistical algorithm designed to identify circadian-regulated transcripts and estimates their period, phase, and amplitude [51]. The dataset of B. rapa gene expression levels that we generated was used for the JTK-CYCLE analysis with a 24 h period. For comparison, we used the Arabidopsis microarray dataset from NCBI (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE3416). Genes with a q-value < 0.05 were classified as rhythmic. Circadian-regulated genes from JTK-CYCLE were clustered using the MeV v.4.8.1 software [52]. Transcription factors (TFs) were classified into families using plant TF database (PlantTFDB) version 4.0 for TF annotation.
2.4. Quantitative Real-Time Polymerase Chain Reaction
Gene-specific primers were designed using Primer3 software (http://frodo.wi.mit.edu/primer3/) (Table S1). Three-week-old plants grown under a 12/12 h light/dark cycle, following growth under a 16/8 h light/dark cycle for 2 weeks, were used as input samples for qRT-PCR (quantitative real-time PCR). Plant tissues were collected at 4 h intervals, for a 24 h period under light/dark conditions, and for 72 h under continuous light conditions. Complementary DNA was synthesized using cDNA EcoDry Premix (Oligo dT) (Clontech, TaKaRa, Shiga, Japan). For cDNA synthesis, 5 ng of total RNA was incubated with oligo (dT) primers at 42 °C for 60 min, and heated at 70 °C for 10 min to stop the reaction. qRT PCR reactions were performed in three technical replicates using a Bio-Rad system (CFX Connect Real-Time PCR) and SYBR Green I Master Mix in a 20 µL final volume. Relative expression levels were determined as the expression ratio of target gene and β-actin in the same cDNA preparation.
2.5. Data Deposition
Raw read files were submitted to the sequence reads archive (SRA), NCBI database (accession number SRP166211).
3. Results
3.1. Sequencing and Statistical Evaluation of the Brassica rapa Transcriptome
To examine the transcriptome of Chinese cabbage (B. rapa, pekinensis), we entrained two-week-old seedlings under a 12/12 h light–dark cycle for 1 week. Two biological repeats for each sample were harvested at ZT 0, 4, 8, 12, 16, 20, and 24 h (Figure 1).
We isolated RNA from the aerial parts of the plants at seven time points and synthesized cDNA libraries for RNAseq, generating 1,363,558,142 raw reads. After filtering out low quality and unpaired reads, 1,363,198,067 high quality reads were used for mapping the B. rapa transcriptome. We obtained a mapping rate over 70% on the B. rapa reference transcript sequences (Table 1).
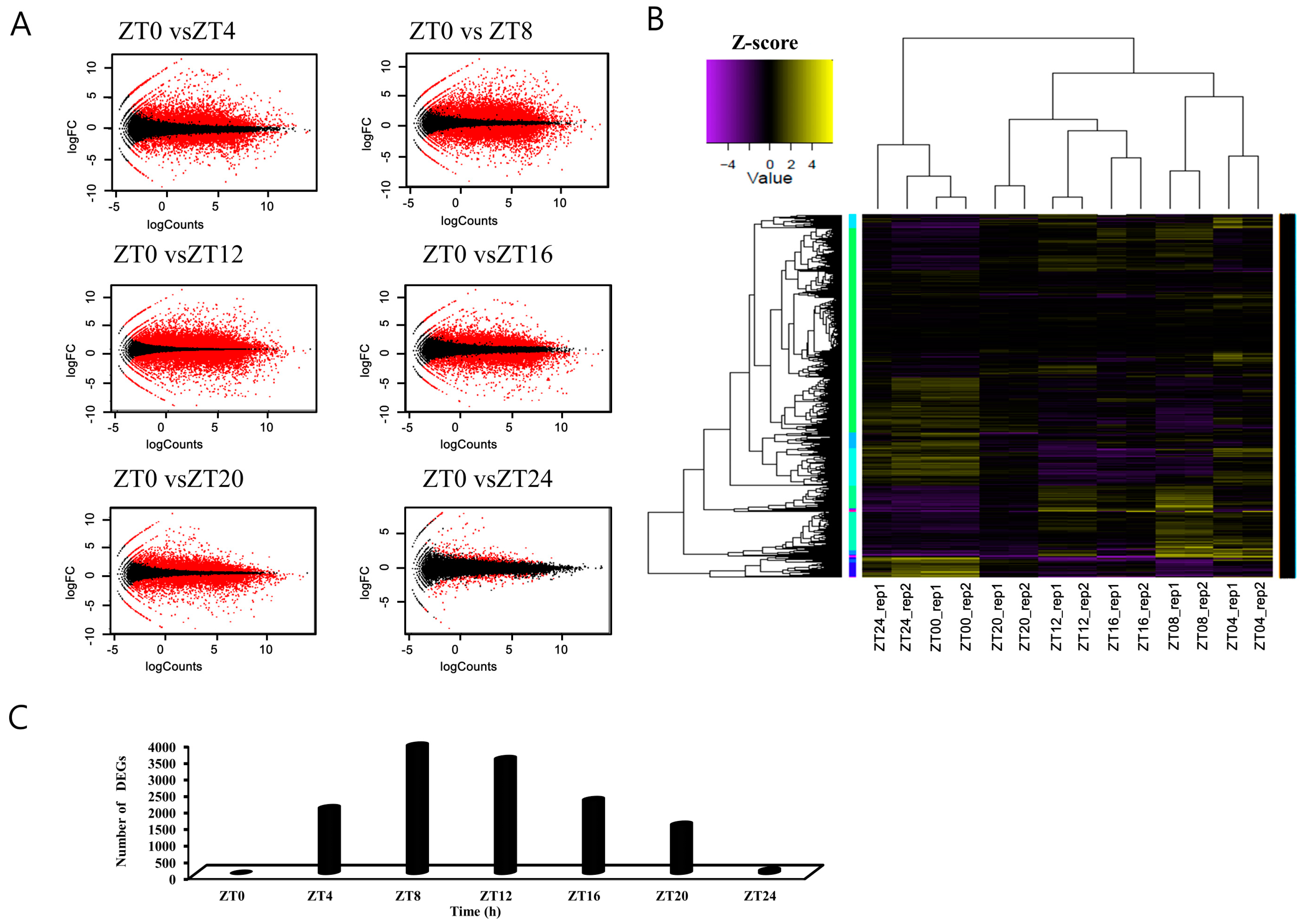
To statistically evaluate the samples used for comparative transcriptomic analysis, we mapped high quality reads to the B. rapa (Phytozome v9.0) database and calculated transcript abundance. We identified differentially expressed genes (DEGs) based on pairwise sample comparisons with ZT0, and visualized DEGs distribution using MA (intensity ratio) plots (Figure 2A,C and Figure S2). In the pairwise ZT comparisons, each sample’s two biological replicates were highly correlated (range ± 1). ZT24 samples harvested during the same phase as the following day ZT0 were also highly correlated (Figure 2B and Figure S1).
3.2. Identification of Differentially Expressed Genes in Pairwise Sample Comparisons
In total, we detected 35,759 DEGs (87.2% of the 41,019 annotated genes) at one or more time points (using a cutoff of an at least twofold change and p <0.05, FDR p-value) in pairwise sample comparisons (Figure 2B). Initially, we compared the gene expression of each time point sample to ZT0 (Figure 2C). Among the DEGs, 1973, 3870, 3461, 2217, and 1471 putative transcripts were differentially expressed at ZT4, 8, 12, 16, and 20, respectively. Comparative analysis between ZT0 and ZT24 resulted in 96 DEGs (Figure 2C). After sunrise (ZT0), the number of DEGs was highest during the evening and at sunset (ZT8 and ZT12, respectively) and decreased sharply thereafter (Figure 2C).
3.3. Conserved Phasing of Diurnally Expressed Genes in B. rapa
Our results identified 11,699 of 41,019 putative unique transcripts (28.5%) in Chinese cabbage, and 5747 of 22,810 transcripts (25.2%) in Arabidopsis as circadian-regulated. Circadian-related transcripts were 3.3% more abundant in Chinese cabbage compared to Arabidopsis (Tables S3 and S4). Circadian-regulated genes clustered into 25 groups (Table S5). In the B. rapa dataset, transcripts with daytime expression peaks were classified into various groups, and those with nighttime peaks were classified into three groups (Figure 3). The phases of cycling genes were compared between A. thaliana and B. rapa datasets. Under diurnal conditions, most orthologs exhibited similar phases, but some showed an inverse expression (data not shown).
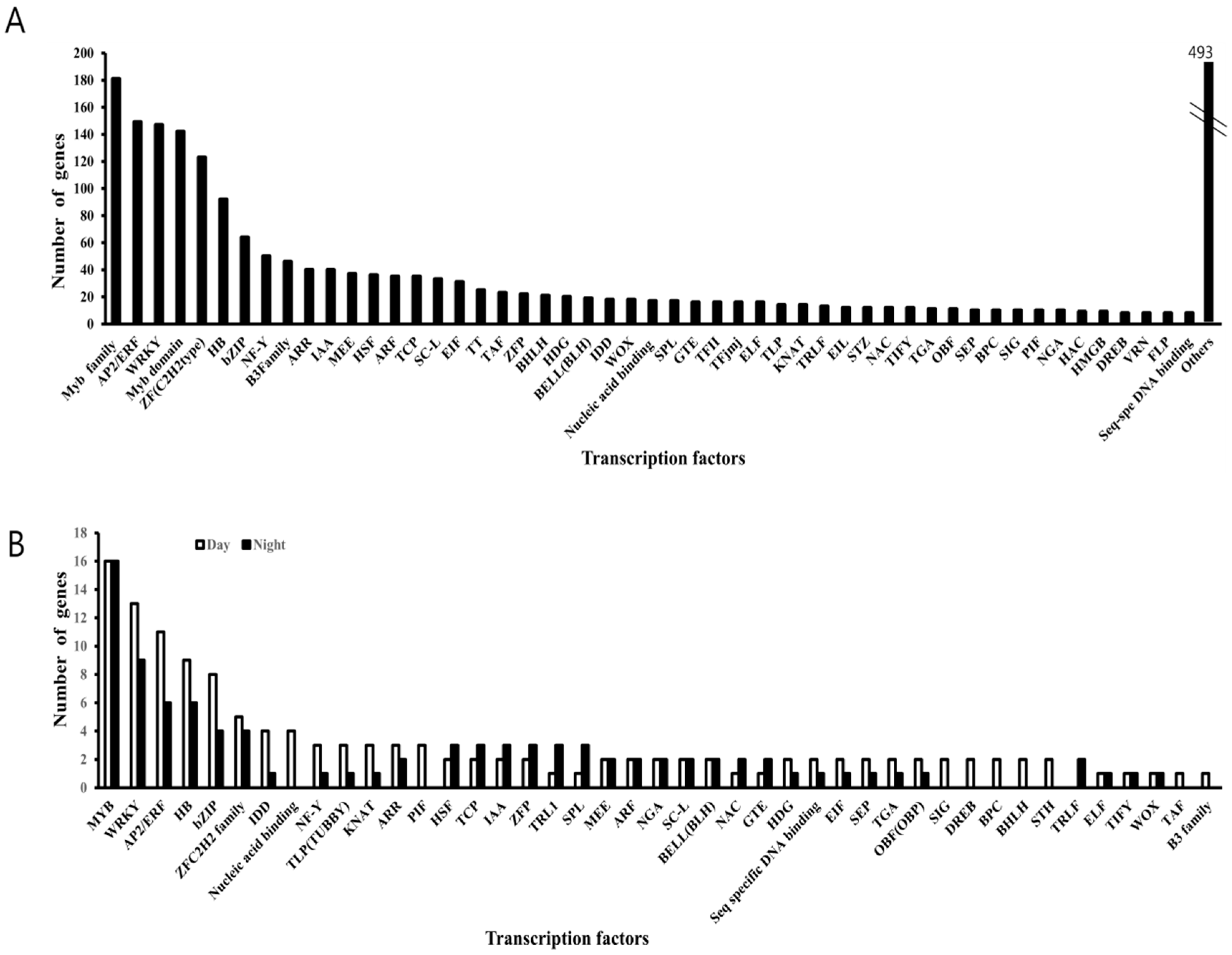
In the present study, 2,249 DEGs were identified as transcription factors (TFs). Functional annotation of these TFs based on their sequence homology and domain sequence similarities with the existing TF database of B. rapa resulting in 51 TF families which were over-represented (32.99% of total TFs); Myb family (181), AP2/ERF (149), WRKY (147), Myb domain (142), and ZF (C2H2 type) (123) TFs (Figure 4A). Among total TFs, 232 genes were circadian clock-regulated based on the DEGs pattern. MYB and WRKY TFs act as key regulators of many processes in plants, of which 16 MYB and 13 WRKY members appeared significantly upregulated during the daytime. The remaining 16 MYB and 9 WRKY members were downregulated during the daytime. Similarly, AP2/ERF, HB, bZIP, ZF (C2H2 type), IDD, NF-Y, TLP, PIF, and TCP are key regulators of plant growth and development. Of these families, 11, 9, 8, 5, 4, 3, 3, 3, and 2 members during daytime and 6, 6, 4, 4, 1, 1, 1, 0, and 3 during nighttime showed peaks of expression, respectively (Figure 4B). The rhythmic expression pattern suggests that these TFs may be related to the circadian function in B. rapa. Of the 51, forty four transcription factors have exhibited significant circadian enrichment (Figure 4B). TFs like Myb-related, basic leucine zipper (bZIP), and multiprotein bridging factor 1 (MBF1) were known associates of the circadian clock-regulated processes [53,54,55,56].
3.4. Expression Anaysis of Clock-Related Gene Paralogs in Chinese Cabbage
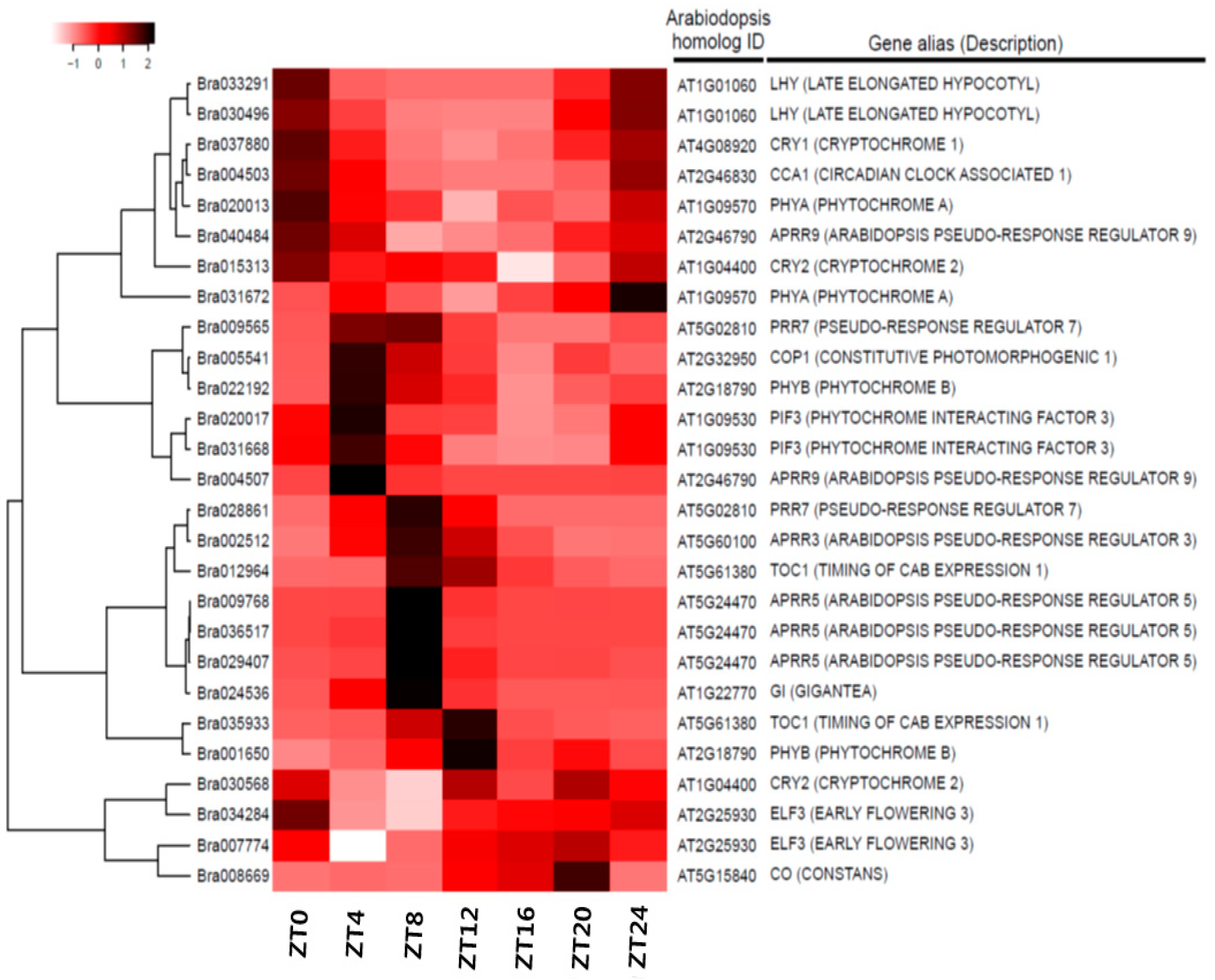
Using Arabidopsis microarray datasets, we screened circadian pathway genes included in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways (Figure S3) and determined their expression levels. Heat maps showed that the expression of these genes changed over time, and all exhibited clear peaks of expression except for PHYA, COP1, and CRY2 (Figure S4). Based on a study of Arabidopsis clock regulation [57,58], we identified putative orthologs of the 21 key Arabidopsis clock-related genes (CRY1, CRY2, PHYA, PHYB, COP1, PIF3, CCA1/LHY, GI, TOC1, ELF3, ELF4, RVE4, RVE8, LUX, FKF1, PRR1, PRR3, PRR5, PRR7, and PRR9) present in the B. rapa dataset (Table 2 and Table S2) and confirmed daily fluctuations in their expression. One to three orthologous copies of individual Arabidopsis clock-related genes displayed highly conserved cycling profiles under diurnal conditions (Figure 5 and Figure S3). The expression cycling profiles of clock-related genes derived from the time course dataset showed similar expression patterns to those of Arabidopsis clock-related genes (Figure S4). The expression pattern of each ortholog resembled that of its corresponding gene in Arabidopsis with respect to, for example, the times at which their expression was highest and lowest (Figure 5 and Figure S4). However, some paralogous copies showed patterns, peak times, or intensities of expression that differed from their paralogs and their orthologs in Arabidopsis. For example, Bra030568—a CRY2 ortholog—did not exhibit expression cycling, and the expression levels of Bra012964 and Bra035933—orthologs of TOC1—peaked at ZT8 and ZT12, respectively (Figure 5).
The RNAseq profiles and phase calls of the listed genes were estimated by calculating the FPKM values, and their expression was compared to their orthologs in Arabidopsis (Figure S5). Expression levels of Bra037880 and Bra024536, orthologs of cryptochrome circadian regulator 1 (CRY1) and GI, were highest and lowest at ZT12 and ZT8, respectively (Figure 6C,G). For Bra004503, a CCA1 ortholog, expression levels were highest at ZT0 and lowest at ZT8-–ZT16 (Figure 5 and Figure S5O). Single copies of these genes were found for the Arabidopsis orthologs, and their expression patterns were highly conserved. Two homologous copies of phytochrome A (PHYA), phytochrome B (PHYB), constitutively photomorphogenic 1 (COP1), reveille 8 (RVE8), late elongated hypocotyl (LHY), and pseudo-response regulator (PRR3) were also found. One of the two paralogs (Bra020013, Bra022192, Bra005541, Bra029778, Bra033291, and Bra002512) showed expression patterns identical to their Arabidopsis orthologs (Figure 5 and Figure S5A,B,E,K,N,Q). However, Bra031672, Bra001650, Bra021818, Bra034074, Bra030496, and Bra020263 were rarely expressed. In Arabidopsis, CRY2 expression did not exhibit a circadian pattern (Figures S4 and S5D). Our results also showed that the two copies of CRY2 maintained steady expression over the course of 1 day (Figure 5 and Figure S5D). The timing of peak expression for the two paralogs of ELF3, PRR1, and PRR7 differed. The expression of Bra007774 (Arabidopsis ELF3) peaked at ZT4, and that of Bra034284 peaked at ZT8 (Figure 5 and Figure S5H). Among the three ELF4 homologs, the expression of Bra017035 peaked early, while Bra000165 and Bra004991 exhibited identical expression patterns (Figure 5 and Figure S5I). However, expression level peaks for the three homologs of Arabidopsis ELF4 occurred at two different times (Figure 5 and Figure S5I). The RVE4 homolog, Bra005754, was expressed at a low level with no diurnal pattern, but the expression of Bra005751 and Bra009562 began decreasing at sunrise, reached ZT8 at nadir, and subsequently increased after ZT16 (Figure 5 and Figure S5J). No homologs of ZTL or FKF1 were found in the B. rapa transcriptome dataset (Table 2 and Table S2). Although each of these genes contains three functional domains—the LOV domain, the F-box motif, and the kelch repeats—and shows high sequence homology [59], only LKP2 matched three homologous genes in our dataset. Bra038832, an LKP2 homolog, showed a circadian expression pattern, but the expression levels of Bra038830 and Bra038831 remained steady over 1 day (Figure 5 and Figure S5M). Of the two PRR1 homologs, Bra012964 expression peaked at ZT8. Furthermore, Bra035933 expression was lower than Bra012964 (Figure 5 and Figure S5P). The three PRR5 homologs were expressed identical to that in Arabidopsis. However, Bra009768 expression peaked at ZT8 (Figure 5 and Figure S5R). The expression pattern of one Bra009565 homolog was identical to that of Arabidopsis PRR7, but that of Bra028861 peaked later at ZT8 (Figure 5 and Figure S5S). Bra033809 and Bra018204 expression peaked at ZT12, but that of the former was lower. The PRR9 homolog, Bra004507, showed the same expression pattern as in Arabidopsis, but that of Bra040484 differed (Figure 5 and Figure S5T). We confirmed the free-running expression of all clock-related genes under continuous light for 72 h by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) (Figure 6 and Figure S6). Expression of most orthologous copies of clock-related genes exhibited a cyclic pattern.
3.5. Differentially Expression of Glucosinolate-Related Gene Paralogs in Chinese Cabbage
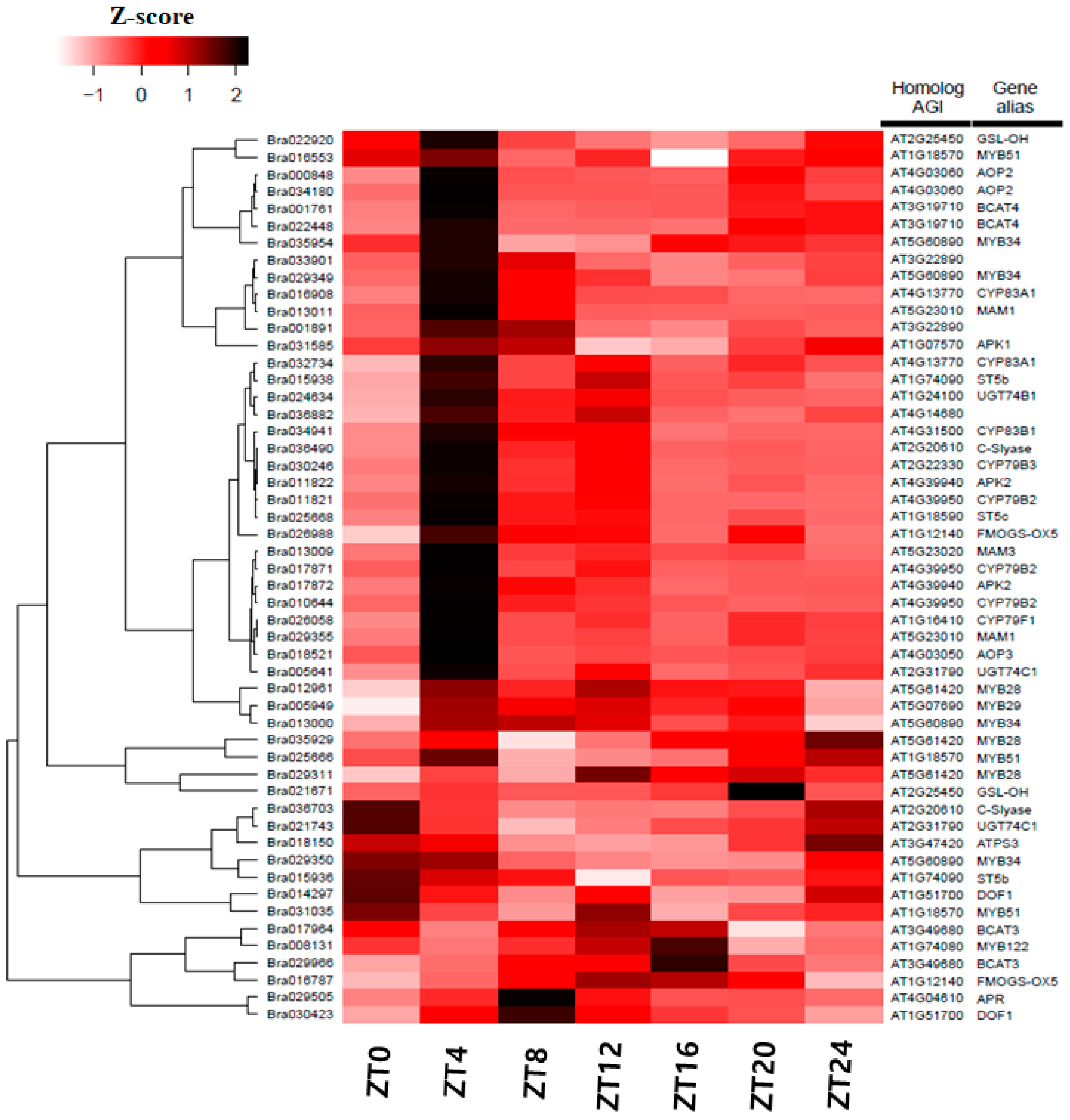
We identified orthologous genes related to the glucosinolate pathway in the B. rapa dataset. Expression of their paralogous genes as a heat map is shown in Figure 7. Though most genes were expressed during the daily cycle—at 4 h after sunrise (ZT4)—genes showed a peak expression. Some MYB TFs were also involved in indolic and aliphatic glucosinolate synthesis. Many Myb transcription factors, except MYB34s, showed peaks at different time points. Most paralogous members showed similar peak time points. Branched-chain aminotransferase (BCAT4) and methylthioalkylmalate synthase (MAM1) involved in the early steps of side chain elongation, and two AOP2 and AOP3 responsible for the side chain modification step in the glucosinolate pathway [60] had peak expression at ZT4. We found that three sulpho-transferases (ST) were involved in the core structure of glucosinolate biosynthesis. Among them, one ST5b (Bra015938) and ST5c (Bra025668) showed high expression at ZT4, and the other ST5b (Bra015936) at ZT0. Expression level then gradually decreased during a 24 h period. Similarly, one FMOGS-OX5 (Bra026988) showed peak expression at ZT4 but the other (Bra016787) at ZT16.
Additionally, other genes such as one DOF1 (Bra014297) had peak expression at ZT0, while others were at ZT8 (Bra030423), ZT4 during the light period (one GSL-OH (Bra022920)), and at ZT20 during the dark period (Bra021671). All CYP genes like CYP79B2, CYP79B3, CYP79F1, CYP83A1, CYP83B1, and CYP83B2 were highly expressed at ZT4 and showed expression throughout the 24 h cycle.
4. Discussion
Photocycles drive oscillations in gene expression. At least 10–30% of genes expressed in Arabidopsis are regulated in a circadian manner [21,62], and up to 89% of transcripts in Arabidopsis display rhythmic expression patterns under a broad set of environmental diurnal/circadian conditions. Indeed, approximately 60% of rice and poplar transcripts displayed rhythmic expression under diurnal/circadian conditions [16].
4.1. Circadian-Regulated Genes in the Brassica Genome
The Brassica genome underwent a whole-genome triplication event after divergence from A. thaliana [45], in addition to several very recent genome duplications. These events make the genus Brassica useful for studies of polyploid genomes’ evolution. The B. rapa genome, which was the first to be sequenced among the Brassica species [45], is 529 Mbp with an approximately 220 Mbp euchromatic region (42% of the genome) [36]. In this study, 28.5% of the 41,019 putative unique transcripts in B. rapa were estimated to be circadian-regulated. These findings are similar to those in Arabidopsis [63], in which 25% of 22,811 transcripts were circadian-regulated. These data suggest that the number of genes whose expression shows a daily cycle and the total number of genes in the Chinese cabbage genome are twofold those in Arabidopsis. In seedling leaf tissues, 35,759 genes (87% of the total of 41,020 annotated genes) were expressed at more than one time point. This is higher than the 32,335 genes (78.8% of the 41,020 annotated genes) expressed in at least one tissue of B. rapa Chiifu [46]. These results suggest that the expression of many genes undergo a daily cycle. In plants, many TFs act as key regulators of various plant functions such as cell cycle, metabolic and physiological pathways, and responses to the environment. More than 2000 TFs have been found in the Arabidopsis genome [64,65,66,67]. Circadian clocks, as timekeeping systems, generate endogenous rhythms with periods of about a day. In the core system based on a transcriptional–translational negative feedback loop, CCA1 and LHY encode closely related single MYB domain transcription factors [68,69]. In addition, various TFs are involved in the regulation of light signal transduction and circadian rhythm [70]. Among the 2269 TFs found in the B. rapa dataset, approximately 14% from each group were found to have a daily cycle of expression (Figure 4B). In Arabidopsis, similar TF genes are regulated by blue light [71]. Nine transcription factor families in Arabidopsis, including NAC, bHLH, C2H2, AP2/ERF, the MYB superfamily, HB, WRKY, bZIP, and MADS are either induced or repressed by blue light [71].
4.2. Circadian Genes Conserved in Brassica after Genome Duplication Events
After triploidization, B. rapa genome segments retained 36–70% of their original genes during fractionation [45]. At the genomic level, approximately double the number of Arabidopsis genes were maintained due to genome shrinkage in the B. rapa genome [36]. After triplication and fractionation, circadian clock-related genes were retained in B. rapa at a significantly higher rate than other gene sets. Furthermore, 65% of the clock-related genes were retained more than one copy—two or three [72]. Our findings are in agreement with previously published findings (Table 2). Two single myeloblastosis (Myb) domain transcription factors play critical roles in the central loop of the Arabidopsis clock [11,73]. The orthologs of LHY/CCA1 are found as three gene family members in B. rapa. One LHY and CCA1 orthologs (Bra033291 and Bra004503) were similarly expressed as those in Arabidopsis, but the expression of Bra030496 was lower in B. rapa (Figure 5 and Figure S5N,O). Among the 18 RVE homologs with roles in clock function or regulation of clock output pathways in Arabidopsis [56,74,75,76,77], our transcriptome dataset contained three and two orthologs of REV4 and REV8, respectively (Table 2). The expression pattern of each was similar to that of its ortholog in Arabidopsis, but the expression of orthologs of REV4 and REV8, Bra005754 and Bra034074, was lower (Figure 5 and Figure S5J,K).
4.3. Expression Patterns of Paralogous Gene Copies
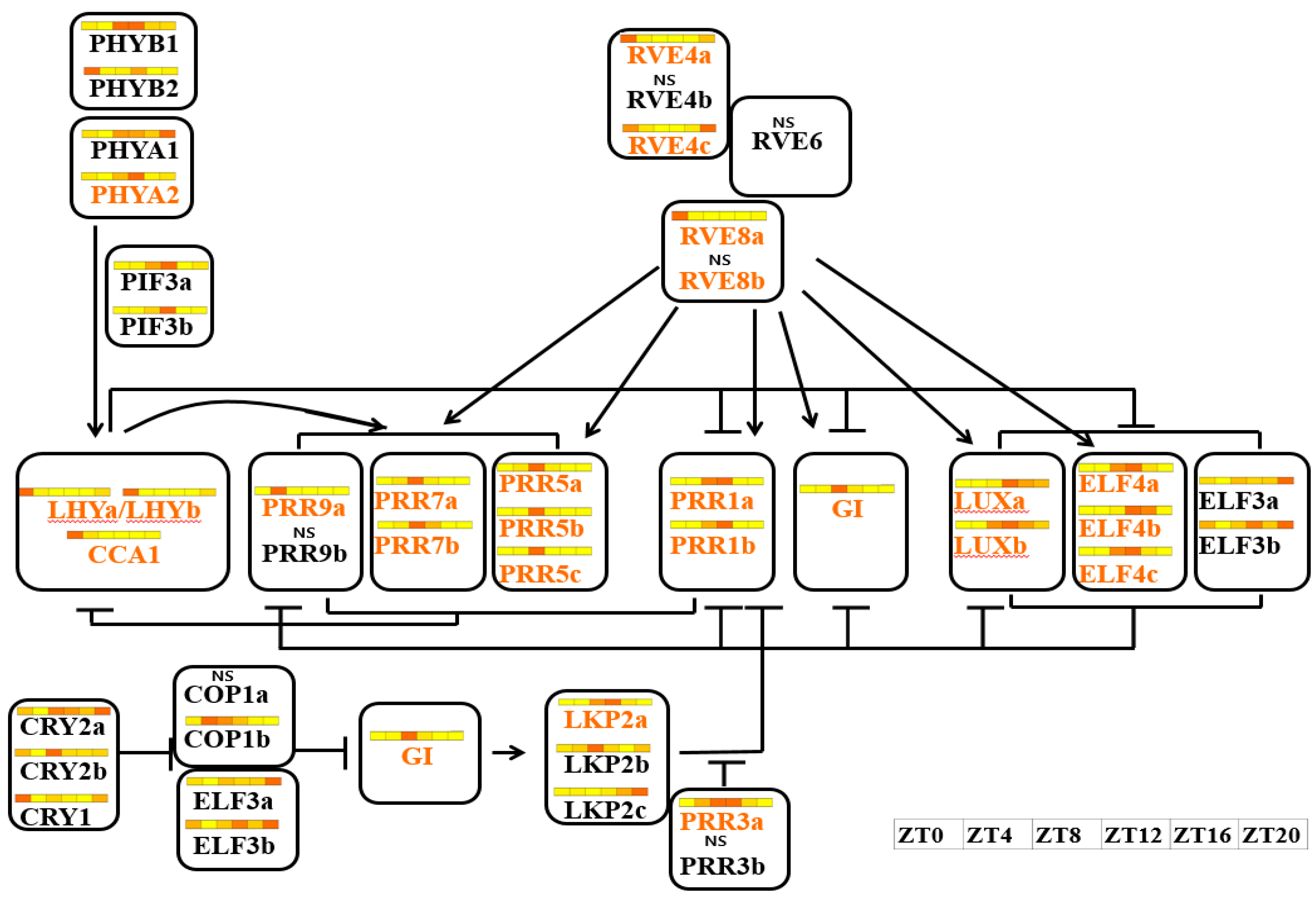
Pseudo-Response Regulator gene, found in the moss Physcomitrella patens, has been generated very early in evolution [78]. After its divergence from Carica papaya, only a single copy of each PRR gene was retained in Arabidopsis. However, after genome triplication in B. rapa, earlier work based on microsynteny analysis estimated a significantly higher rate of retention than that of their neighboring genes [41]. Eleven PRR genes were identified in the B. rapa genome. PRR3b and PRR9b detected as a result of rearrangements and partial deletions were predicted to be nonfunctional or neo-functional [41]. We confirmed the rare expression of PRR3b, with very low FPKM values and no specific qRT-PCR evidence (Figure 5 and Figure S5Q). Although it was not detected by qRT-PCR (Figure 5 and Figure S5Q), RNAseq showed that PRR9b expression was higher than PRR9a (Figure 5 and Figure S5T). PRR3 modulates the stability of TOC1 (PRR1) through a direct protein–protein interaction [79]. In the B. rapa genome, the expression of two PRR1 orthologs peaked at different time points (Table 2 and Figure S5P), suggesting that the circadian clock in B. rapa is regulated by a novel mechanism.
In Arabidopsis, ZTL and its close relatives LKP2 and FKF1 determine the length of the circadian period by regulating the degradation of TOC1 [80,81]. These three genes encode F-box proteins with an N-terminal LOV domain, a central F-box, and a series of Kelch repeats at the C-terminus [41,59]. Although their sequences are similar, ZTL and FKF1 were not found and three tightly linked copies of Arabidopsis LKP2 were found in B. rapa [41]. Based on their domain structures, FKF1 in Arabidopsis has one less repeat (four) compared to ZTL and LKP2 (five each). In B. rapa, LKP2a (Bra038832) has one less repeat (four) compared to LKP2b (Bra038831) and LKP2c (Bra038830) (five each) [41]. Interestingly, the expression of LKP2a (Bra038832) peaked at around ZT8, similar to Arabidopsis FKF1 (Figures S5M,U,V and S6). LKP2b and LKP2c did not show a rhythmic expression pattern. Arabidopsis ZTL, LKP2, and FKF1 play overlapping roles in circadian period determination. Although ZTL predominates, FKF1 plays the major role in flowering time determination [80,81,82]. The functional specialization of the three copies of LKP2 in B. rapa warrants further research. Homologous genes with multiple copies are considered to be functionally similar. Orthologous copies have redundant function and may show functional specialization or sub-functionalization compared to the ancestral copy [83]. However, one or more members may acquire new functions, while others retain their original function [84,85,86,87,88]. Such changes may be advantageous for preserving fitness. We found different expression levels or phase shifts among paralogous copies (Figure S5). Further research on the function of each paralogous copy is needed to explain the circadian rhythm of gene expression in crop species.
4.4. Regulation of Circadian-Mediated Biological Processes in Crops
The understanding of the circadian clock and its regulatory processes can be effectively utilized to enhance plant growth and fitness [20,57,89,90]. The synthesis of glucosinolates, a major secondary metabolite in B. rapa, has been reported as a light-regulated pathway. In the first 4 h of re-illumination of dark-adapted plants, the expression of many glucosinolate genes increased, inducing accumulation of mRNA [91]. BCAT4 and MAM1 are involved in the early side chain elongation process, and AOP2 in the secondary modification of the side chain, which showed higher expression levels under the light condition compared to darkness. They maintained high expression levels when the plants were kept under continuous light [60,91]. Reciprocally, glucosinolate genotypes altered the periodicity of a clock output unrelated to glucosinolates, the photochemical state of photosystem II [92]. These findings suggest that the manipulation of circadian clock genes can be valuable for improving plant metabolism. Studies on the expression patterns of clock-related genes in crop species will enable the formulation of effective crop-breeding strategies.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/10/2/130/s1. Figure S1: Statistical analysis of differentially expressed genes across repeat samples; Figure S2: Statistical analysis of significantly expressed genes at each time point versus ZT0; Figure S3: Circadian-related genes in Arabidopsis and B. rapa.; Figure S4: Expression of clock-related genes over time in Arabidopsis and PCC values between Br and At; Figure S5: Expression of clock-related genes in Arabidopsis and B. rapa. Bra numbers indicate orthologs of PHYA (A), PHYB (B), CRY1 (C), CRY2 (D), COP1 (E), PIF3 (F), GI (G), ELF3 (H), ELF4 (I), RVE4 (J), RVE8 (K), LUX (L), LKP2 (M), LHY (N), CCA1 (O), PRR1 (P), PRR3 (Q), PRR5 (R), PRR7 (S), PRR9 (T), ZTL(U), and FKF1(V); Figure S6: Free-running expression of clock-related genes in B. rapa; Table S1: qRT-PCR primers used in this work.; Table S2: B. rapa_circadian_normalized_FPKM and orthologous identification; Table S3: JTK-cycle results of the Arabidopsis data set; Table S4: JTK-cycle results of the B. rapa data set; Table S5: Clustering results of cycling genes in B. rapa.
Author Contributions
Conceptualization: J.A.K.; methodology and bioinformatical analysis: D.S.; formal analysis: S.K.; validation: H.-e.J.; formal analysis: K.-H.J. and H.J.; resources: W.Y.K., S.I.L., and M.-J.J.; writing—original draft preparation: J.A.K.; writing—review and editing: D.S., S.K.
Funding
This work was supported by grants from the Research Program for Agricultural Science & Technology Development, National Academy of Agricultural Science (project No. PJ01247203), and from the BioGreen 21 Program of the Rural Development Administration, Republic of Korea (project No. PJ01327303).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hangarter, R.P. Gravity, light and plant form. Plant Cell Environ. 1997, 20, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.E.; Millar, A.J. The circadian clock. A plant’s best friend in a spinning world. Plant Physiol. 2003, 132, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Yakir, E.; Hilman, D.; Harir, Y.; Green, R.M. Regulation of output from the plant circadian clock. FEBS J. 2007, 274, 335–345. [Google Scholar] [CrossRef]
- Covington, M.F.; Maloof, J.N.; Straume, M.; Kay, S.A.; Harmer, S.L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol. 2008, 9, R130. [Google Scholar] [CrossRef] [PubMed]
- Fenske, M.P.; Imaizumi, T. Circadian Rhythms in Floral Scent Emission. Front. Plant Sci. 2016, 7, 462. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.; Shin, J.; Davis, S.J. Abiotic stress and the plant circadian clock. Plant Signal. Behav. 2011, 6, 223–231. [Google Scholar] [CrossRef]
- McClung, C.R. Beyond Arabidopsis: The circadian clock in non-model plant species. Semin. Cell Dev. Biol. 2013, 24, 430–436. [Google Scholar] [CrossRef]
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef]
- Lin, C. Blue light receptors and signal transduction. Plant Cell 2002, 14, S207–S225. [Google Scholar] [CrossRef]
- Kircher, S.; Kozma-Bognar, L.; Kim, L.; Adam, E.; Harter, K.; Schafer, E.; Nagy, F. Light quality-dependent nuclear import of the plant photoreceptors phytochrome A and B. Plant Cell 1999, 11, 1445–1456. [Google Scholar] [CrossRef]
- Harmer, S.L. The circadian system in higher plants. Annu. Rev. Plant Biol. 2009, 60, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Marguerat, S.; Bahler, J. RNA-seq: From technology to biology. Cell Mol. Life Sci. 2010, 67, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.E.; Grant, G.R.; Paquin, C.; Qian, J.; Nitabach, M.N. Deep sequencing the circadian and diurnal transcriptome of Drosophila brain. Genome Res. 2012, 22, 1266–1281. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Aoki, K.; Nagano, A.J.; Honjo, M.N.; Fukuda, H. Circadian oscillation of the lettuce transcriptome under constant light and light–dark conditions. Front. Plant Sci. 2016, 7, 1114. [Google Scholar] [CrossRef] [PubMed]
- Facella, P.; Lopez, L.; Carbone, F.; Galbraith, D.W.; Giuliano, G.; Perrotta, G. Diurnal and circadian rhythms in the tomato transcriptome and their modulation by cryptochrome photoreceptors. PLoS ONE 2008, 3, e2798. [Google Scholar] [CrossRef] [PubMed]
- Filichkin, S.A.; Breton, G.; Priest, H.D.; Dharmawardhana, P.; Jaiswal, P.; Fox, S.E.; Michael, T.P.; Chory, J.; Kay, S.A.; Mockler, T.C. Global profiling of rice and poplar transcriptomes highlights key conserved circadian-controlled pathways and cis-regulatory modules. PLoS ONE 2011, 6, e16907. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.R. Comes a time. Curr. Opin. Plant Biol. 2008, 11, 514–520. [Google Scholar] [CrossRef]
- Farre, E.M.; Weise, S.E. The interactions between the circadian clock and primary metabolism. Curr. Opin. Plant Biol. 2012, 15, 293–300. [Google Scholar] [CrossRef]
- Farre, E.M. The regulation of plant growth by the circadian clock. Plant Biol. (Stuttg.) 2012, 14, 401–410. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, H.S.; Choi, S.H.; Jang, J.Y.; Jeong, M.J.; Lee, S.I. The importance of the circadian clock in regulating plant metabolism. Int. J. Mol. Sci. 2017, 18, 2680. [Google Scholar] [CrossRef]
- Michael, T.P.; Mockler, T.C.; Breton, G.; McEntee, C.; Byer, A.; Trout, J.D.; Hazen, S.P.; Shen, R.; Priest, H.D.; Sullivan, C.M.; et al. Network discovery pipeline elucidates conserved time-of-day-specific cis-regulatory modules. PLoS Genet. 2008, 4, e14. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, T.L.; Field, C.B. Circadian rhythms in photosynthesis: Oscillations in carbon assimilation and stomatal conductance under constant conditions. Plant Physiol. 1991, 96, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Rikin, A.; Dillwith, J.W.; Bergman, D.K. Correlation between the circadian rhythm of resistance to extreme temperatures and changes in fatty acid composition in cotton seedlings. Plant Physiol. 1993, 101, 31–36. [Google Scholar] [CrossRef]
- Hudson, K.A. The circadian clock-controlled transcriptome of developing soybean seeds. Plant Genome-US 2010, 3, 3–13. [Google Scholar] [CrossRef]
- Kim, J.A.; Jung, H.E.; Hong, J.K.; Hermand, V.; Robertson McClung, C.; Lee, Y.H.; Kim, J.Y.; Lee, S.I.; Jeong, M.J.; Kim, J.; et al. Reduction of GIGANTEA expression in transgenic Brassica rapa enhances salt tolerance. Plant Cell Rep. 2016, 35, 1943–1954. [Google Scholar] [CrossRef]
- Izawa, T.; Mihara, M.; Suzuki, Y.; Gupta, M.; Itoh, H.; Nagano, A.J.; Motoyama, R.; Sawada, Y.; Yano, M.; Hirai, M.Y.; et al. Os-GIGANTEA Confers Robust Diurnal Rhythms on the Global Transcriptome of Rice in the Field. Plant Cell 2011, 23, 1741–1755. [Google Scholar] [CrossRef]
- Xie, Q.G.; Lou, P.; Hermand, V.; Aman, R.; Park, H.J.; Yun, D.J.; Kim, W.Y.; Salmela, M.J.; Ewers, B.E.; Weinig, C.; et al. Allelic polymorphism of GIGANTEA is responsible for naturally occurring variation in circadian period in Brassica rapa. Proc. Natl. Acad. Sci. USA 2015, 112, 3829–3834. [Google Scholar] [PubMed]
- Navarro, C.; Abelenda, J.A.; Cruz-Oro, E.; Cuellar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of flowering and storage organ formation in potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, S.P.; Lan, T.H.; Feldmann, K.A.; Paterson, A.H. Comparative mapping of Arabidopsis thaliana and Brassica oleracea chromosomes reveals islands of conserved organization. Genetics 1994, 138, 499–510. [Google Scholar]
- Grant, D.; Cregan, P.; Shoemaker, R.C. Genome organization in dicots: Genome duplication in Arabidopsis and synteny between soybean and Arabidopsis. Proc. Natl. Acad. Sci. USA 2000, 97, 4168–4173. [Google Scholar] [CrossRef]
- Bowers, J.E.; Abbey, C.; Anderson, S.; Chang, C.; Draye, X.; Hoppe, A.H.; Jessup, R.; Lemke, C.; Lennington, J.; Li, Z.K.; et al. A high-density genetic recombination map of sequence-tagged sites for Sorghum, as a framework for comparative structural and evolutionary genomics of tropical grains and grasses. Genetics 2003, 165, 367–386. [Google Scholar]
- Gebhardt, C.; Walkemeier, B.; Henselewski, H.; Barakat, A.; Delseny, M.; Stuber, K. Comparative mapping between potato (Solanum tuberosum) and Arabidopsis thaliana reveals structurally conserved domains and ancient duplications in the potato genome. Plant J. 2003, 34, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Salathia, N.; Lynn, J.R.; Millar, A.J.; King, G.J. Detection and resolution of genetic loci affecting circadian period in Brassica oleracea. Theor. Appl. Genet. 2007, 114, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Hou, S.B.; Feng, Y.; Yu, Q.Y.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.H.; Kwon, S.J.; Yang, T.J.; Seol, Y.J.; Jin, M.; Kim, J.A.; Lim, M.H.; Kim, J.S.; Baek, S.; Choi, B.S.; et al. Genome-wide comparative analysis of the Brassica rapa gene space reveals genome shrinkage and differential loss of duplicated genes after whole genome triplication. Genome Biol. 2009, 10, R111. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.H.; Kwon, S.J.; Seol, Y.J.; Kim, J.A.; Jin, M.; Kim, J.S.; Lim, M.H.; Lee, S.I.; Hong, J.K.; Park, T.H.; et al. Sequence and structure of Brassica rapa chromosome A3. Genome Biol. 2010, 11, R94. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Ashikari, M.; Miura, K.; Yamashino, T.; Mizuno, T. The evolutionarily conserved OsPRR quintet: Rice pseudo-response regulators implicated in circadian rhythm. Plant Cell Physiol. 2003, 44, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Takata, N.; Saito, S.; Saito, C.T.; Uemura, M. Phylogenetic footprint of the plant clock system in angiosperms: Evolutionary processes of pseudo-response regulators. BMC Evol. Biol. 2010, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Kim, J.S.; Hong, J.K.; Lee, Y.H.; Choi, B.S.; Seol, Y.J.; Jeon, C.H. Comparative mapping, genomic structure, and expression analysis of eight pseudo-response regulator genes in Brassica rapa. Mol. Genet. Genomics 2012, 287, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Lou, P.; Wu, J.; Cheng, F.; Cressman, L.G.; Wang, X.W.; McClung, C.R. Preferential retention of circadian clock genes during diploidization following whole genome triplication in Brassica rapa. Plant Cell 2012, 24, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Lee, S.; Seo, P.J.; Yang, M.S.; Park, C.M. Identification and molecular characterization of a Brachypodium distachyon GIGANTEA gene: Functional conservation in monocot and dicot plants. Plant Mol. Biol. 2010, 72, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Chung, T.Y.; King, G.J.; Jin, M.; Yang, T.J.; Jin, Y.M.; Kim, H.I.; Park, B.S. A sequence-tagged linkage map of Brassica rapa. Genetics 2006, 174, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Campo, C. Biology of Brassica Coenospecies, 1st ed.; Elsevier: Amsterdam, The Netherlands; New York, NY, USA, 1999. [Google Scholar]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Wang, X.; Yu, J.; Wu, J.; Li, W.; Huang, J.; Dong, C.; Hua, W.; Liu, S. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genomics 2013, 14, 689. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.; Edwards, R. Fast identification and removal of sequence contamination from genomic and metagenomic datasets. PLoS ONE 2011, 6, e17288. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11–17. [Google Scholar]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Robinson, M.D.; Smyth, G.K. Small-sample estimation of negative binomial dispersion, with applications to SAGE data. Biostatistics 2008, 9, 321–332. [Google Scholar] [CrossRef]
- Hughes, M.E.; Hogenesch, J.B.; Kornacker, K. JTK_CYCLE: An efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythms 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.F.; Wang, Z.Y. Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J. 2005, 43, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.P.; Schultz, T.F.; Pruneda-Paz, J.L.; Borevitz, J.O.; Ecker, J.R.; Kay, S.A. LUX ARRHYTHMO encodes a Myb domain protein essential for circadian rhythms. Proc. Natl. Acad. Sci. USA 2005, 102, 10387–10392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Chen, Y.H.; Wang, Z.Y.; Chen, Z.L.; Gu, H.Y.; Qu, L.J. Constitutive expression of CIR1 (RVE2) affects several circadian-regulated processes and seed germination in Arabidopsis. Plant J. 2007, 51, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.Y.; Harmer, S.L. Wheels within wheels: The plant circadian system. Trends Plant Sci. 2014, 19, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Pokhilko, A.; Fernandez, A.P.; Edwards, K.D.; Southern, M.M.; Halliday, K.J.; Millar, A.J. The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops. Mol. Syst. Biol. 2012, 8, 574. [Google Scholar] [CrossRef]
- Takase, T.; Nishiyama, Y.; Tanihigashi, H.; Ogura, Y.; Miyazaki, Y.; Yamada, Y.; Kiyosue, T. LOV KELCH PROTEIN2 and ZEITLUPE repress Arabidopsis photoperiodic flowering under non-inductive conditions, dependent on FLAVIN-BINDING KELCH REPEAT F-BOX1. Plant J. 2011, 67, 608–621. [Google Scholar] [CrossRef]
- Schuster, J.; Knill, T.; Reichelt, M.; Gershenzon, J.; Binder, S. BRANCHED-CHAIN AMINOTRANSFERASE4 is part of the chain elongation pathway in the biosynthesis of methionine-derived glucosinolates in Arabidopsis. Plant Cell 2006, 18, 2664–2679. [Google Scholar] [CrossRef]
- Zang, Y.X.; Kim, H.U.; Kim, J.A.; Lim, M.H.; Jin, M.; Lee, S.C.; Kwon, S.J.; Lee, S.I.; Hong, J.K.; Park, T.H.; et al. Genome-wide identification of glucosinolate synthesis genes in Brassica rapa. FEBS J. 2009, 276, 3559–3574. [Google Scholar] [CrossRef]
- Mockler, T.C.; Michael, T.P.; Priest, H.D.; Shen, R.; Sullivan, C.M.; Givan, S.A.; McEntee, C.; Kay, S.A.; Chory, J. The DIURNAL project: DIURNAL and circadian expression profiling, model-based pattern matching, and promoter analysis. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Blasing, O.E.; Gibon, Y.; Gunther, M.; Hohne, M.; Morcuende, R.; Osuna, D.; Thimm, O.; Usadel, B.; Scheible, W.R.; Stitt, M. Sugars and circadian regulation make major contributions to the global regulation of diurnal gene expression in Arabidopsis. Plant Cell 2005, 17, 3257–3281. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.Y.; He, K.; Liu, D.; Bai, S.N.; Gu, X.C.; Wei, L.P.; Luo, J.C. DATF: A database of Arabidopsis transcription factors. Bioinformatics 2005, 21, 2568–2569. [Google Scholar] [CrossRef] [PubMed]
- Iida, K.; Seki, M.; Sakurai, T.; Satou, M.; Akiyama, K.; Toyoda, T.; Konagaya, A.; Shinozaki, K. RARTF: Database and tools for complete sets of Arabidopsis transcription factors. DNA Res. 2005, 12, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Davuluri, R.V.; Sun, H.; Palaniswamy, S.K.; Matthews, N.; Molina, C.; Kurtz, M.; Grotewold, E. AGRIS: Arabidopsis gene regulatory information server, an information resource of Arabidopsis cis-regulatory elements and transcription factors. BMC Bioinform. 2003, 4, 25. [Google Scholar] [CrossRef]
- Riano-Pachon, D.M.; Ruzicic, S.; Dreyer, I.; Mueller-Roeber, B. PlnTFDB: An integrative plant transcription factor database. BMC Bioinform. 2007, 8, 42. [Google Scholar] [CrossRef]
- Carre, I.A.; Kim, J.Y. MYB transcription factors in the Arabidopsis circadian clock. J. Exp. Bot. 2002, 53, 1551–1557. [Google Scholar] [CrossRef]
- Green, R.M.; Tobin, E.M. The role of CCA1 and LHY in the plant circadian clock. Dev. Cell 2002, 2, 516–518. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Jeong, J.; Kang, M.Y.; Kim, J.; Paek, N.C.; Choi, G. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis. Nat. Commun. 2014, 5, 4636. [Google Scholar] [CrossRef]
- Jiao, Y.L.; Yang, H.J.; Ma, L.G.; Sun, N.; Yu, H.Y.; Liu, T.; Gao, Y.; Gu, H.Y.; Chen, Z.L.; Wada, M.; et al. A genome-wide analysis of blue-light regulation of Arabidopsis transcription factor gene expression during seedling development. Plant Physiol. 2003, 133, 1480–1493. [Google Scholar] [CrossRef]
- Hofmann, N.R. Evolution of the circadian clock in a whole-genome context. Plant Cell 2012, 24, 2239. [Google Scholar] [CrossRef]
- McClung, C.R. The genetics of plant clocks. Adv. Genet. 2011, 74, 105–139. [Google Scholar] [PubMed]
- Kuno, N.; Moller, S.G.; Shinomura, T.; Xu, X.M.; Chua, N.H.; Furuya, M. The novel MYB protein EARLY-PHYTOCHROME-RESPONSIVE1 is a component of a slave circadian oscillator in Arabidopsis. Plant Cell 2003, 15, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- Farinas, B.; Mas, P. Functional implication of the MYB transcription factor RVE8/LCL5 in the circadian control of histone acetylation. Plant J. 2011, 66, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Rawat, R.; Takahashi, N.; Hsu, P.Y.; Jones, M.A.; Schwartz, J.; Salemi, M.R.; Phinney, B.S.; Harmer, S.L. REVEILLE8 and PSEUDO-REPONSE REGULATOR5 form a negative feedback loop within the Arabidopsis circadian clock. PLoS Genet. 2011, 7, e1001350. [Google Scholar] [CrossRef] [PubMed]
- Rawat, R.; Schwartz, J.; Jones, M.A.; Sairanen, I.; Cheng, Y.F.; Andersson, C.R.; Zhao, Y.D.; Ljung, K.; Harmer, S.L. REVEILLE1, a Myb-like transcription factor, integrates the circadian clock and auxin pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 16883–16888. [Google Scholar] [CrossRef] [PubMed]
- Satbhai, S.B.; Yamashino, T.; Mizuno, T.; Aoki, S. Heterologous expression and functional characterization of a Physcomitrella pseudo response regulator homolog, PpPRR2, in Arabidopsis. Biosci. Biotechnol. Biochem. 2011, 75, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Para, A.; Farre, E.M.; Imaizumi, T.; Pruneda-Paz, J.L.; Harmon, F.G.; Kay, S.A. PRR3 is a vascular regulator of TOC1 stability in the Arabidopsis circadian clock. Plant Cell 2007, 19, 3462–3473. [Google Scholar] [CrossRef] [PubMed]
- Somers, D.E.; Schultz, T.F.; Milnamow, M.; Kay, S.A. ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis. Cell 2000, 101, 319–329. [Google Scholar] [CrossRef]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.H.; Tobin, E.M.; et al. F-Box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Lasswell, J.; Rogg, L.E.; Cohen, M.A.; Bartel, B. FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell 2000, 101, 331–340. [Google Scholar] [CrossRef]
- McClung, C.R. A modern circadian clock in the common angiosperm ancestor of monocots and eudicots. BMC Biol. 2010, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G.; Baumlein, H.; Mock, H.P.; Himmelbach, A.; Schweizer, P. The multigene family encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant Physiol. 2006, 142, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Markov, G.V.; Paris, M.; Bertrand, S.; Laudet, V. The evolution of the ligand/receptor couple: A long road from comparative endocrinology to comparative genomics. Mol. Cell Endocrinol. 2008, 293, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.H.; Huang, Y.; Liu, C.; Ruan, Y.; Shen, W.H. Functional conservation and divergence of J-domain-containing ZUO1/ZRF orthologs throughout evolution. Planta 2014, 239, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fan, T.; Song, J.; Sun, W.; Xia, K.; Liao, J.; Zhang, M. Functional conservation and divergence of four ginger AP1/AGL9 MADS-box genes revealed by analysis of their expression and protein-protein interaction, and ectopic expression of AhFUL gene in Arabidopsis. PLoS ONE 2014, 9, e114134. [Google Scholar] [CrossRef]
- Roulin, A.; Auer, P.L.; Libault, M.; Schlueter, J.; Farmer, A.; May, G.; Stacey, G.; Doerge, R.W.; Jackson, S.A. The fate of duplicated genes in a polyploid plant genome. Plant J. 2013, 73, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.E.; Ewers, B.E.; Williams, D.G.; Xie, Q.; Lou, P.; Xu, X.; McClung, C.R.; Weinig, C. The genetic architecture of ecophysiological and circadian traits in Brassica rapa. Genetics 2011, 189, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, Y.; Yeom, M.; Kim, J.H.; Nam, H.G. FIONA1 is essential for regulating period length in the Arabidopsis circadian clock. Plant Cell 2008, 20, 307–319. [Google Scholar] [CrossRef]
- Huseby, S.; Koprivova, A.; Lee, B.R.; Saha, S.; Mithen, R.; Wold, A.B.; Bengtsson, G.B.; Kopriva, S. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar] [CrossRef]
- Kerwin, R.E.; Jimenez-Gomez, J.M.; Fulop, D.; Harmer, S.L.; Maloof, J.N.; Kliebenstein, D.J. Network quantitative trait loci mapping of circadian clock outputs identifies metabolic pathway-to-clock linkages in Arabidopsis. Plant Cell 2011, 23, 471–485. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chinese cabbage cultivation. Chinese cabbage seedlings were grown in a growth chamber in individual pots under a 16/8-h light/dark cycle for 2 weeks, with a cool-white fluorescent lamp. Plants were entrained for 1 week under a 12/12 h light/dark cycle. Aerial parts of three-week-old plants were harvested at ZT 0, 4, 8, 12, 16, 20, and 24 h, with two biological replicates. The plants were watered every three days, and the temperature was maintained at 23 °C until 3–5 true leaves emerged. White and black boxes indicate day (light on) and night (light off), respectively. ZT: zeitgeber time; ZT0: sunrise.
Figure 1.
Chinese cabbage cultivation. Chinese cabbage seedlings were grown in a growth chamber in individual pots under a 16/8-h light/dark cycle for 2 weeks, with a cool-white fluorescent lamp. Plants were entrained for 1 week under a 12/12 h light/dark cycle. Aerial parts of three-week-old plants were harvested at ZT 0, 4, 8, 12, 16, 20, and 24 h, with two biological replicates. The plants were watered every three days, and the temperature was maintained at 23 °C until 3–5 true leaves emerged. White and black boxes indicate day (light on) and night (light off), respectively. ZT: zeitgeber time; ZT0: sunrise.

Figure 2.
Statistical analysis of differentially expressed genes across samples. MA plots showing pairwise comparisons of transcript levels across samples. ZT0, 4, 8, 12, 16, 20, and 24 indicate time points after the light-on time. Y-axis: log2 fold change (logFC) between the two samples; X-axis: log2 average count normalized to the size factor. Red dots: transcripts with a logFC significantly higher than 2 or lower than −2; black dots: transcripts with a logFC of −2 to 2 (A). Clustered heatmap showing the Pearson correlation matrix for pairwise sample comparisons. The color key was adjusted based on the log2-centered values for optimal visual detection of differences, and the dendrograms show the distance between samples (B). The number of significantly expressed genes (FDR p <0.05) at each time point versus ZT0 (C).
Figure 2.
Statistical analysis of differentially expressed genes across samples. MA plots showing pairwise comparisons of transcript levels across samples. ZT0, 4, 8, 12, 16, 20, and 24 indicate time points after the light-on time. Y-axis: log2 fold change (logFC) between the two samples; X-axis: log2 average count normalized to the size factor. Red dots: transcripts with a logFC significantly higher than 2 or lower than −2; black dots: transcripts with a logFC of −2 to 2 (A). Clustered heatmap showing the Pearson correlation matrix for pairwise sample comparisons. The color key was adjusted based on the log2-centered values for optimal visual detection of differences, and the dendrograms show the distance between samples (B). The number of significantly expressed genes (FDR p <0.05) at each time point versus ZT0 (C).

Figure 3.
Clustering of 11,699 clock-regulated genes according to their diurnal cycle expression patterns in Chinese cabbage seedlings using JTK-CYCLE [51]. Thick boxes indicate groups that peaked during night time. Using JTK-CYCLE, 11,699 circadian-regulated genes were identified in leaves of Chinese cabbage seedlings growing under a 12/12 h light/dark cycle. These genes were clustered based on their diurnal expression. The k-means clustering was performed on fragments per kb of exon per million (FPKM) average values of the two repeats at 4 h intervals over 24 h using the MeV v. 4.8.1 software [52]. The number of genes in each cluster is shown.
Figure 3.
Clustering of 11,699 clock-regulated genes according to their diurnal cycle expression patterns in Chinese cabbage seedlings using JTK-CYCLE [51]. Thick boxes indicate groups that peaked during night time. Using JTK-CYCLE, 11,699 circadian-regulated genes were identified in leaves of Chinese cabbage seedlings growing under a 12/12 h light/dark cycle. These genes were clustered based on their diurnal expression. The k-means clustering was performed on fragments per kb of exon per million (FPKM) average values of the two repeats at 4 h intervals over 24 h using the MeV v. 4.8.1 software [52]. The number of genes in each cluster is shown.

Figure 4.
Classification of transcription factors (TFs) in the B. rapa transcriptome data set. (A) Total number of differentially expressed genes (DEGs) encoding TFs. (B) Forty-four transcription factors are circadian-regulated, where the white bar graphs represent genes upregulated in the daytime (ZR0–ZT12), and the black bar graph represents genes upregulated in the night time (ZR12–ZT24).
Figure 4.
Classification of transcription factors (TFs) in the B. rapa transcriptome data set. (A) Total number of differentially expressed genes (DEGs) encoding TFs. (B) Forty-four transcription factors are circadian-regulated, where the white bar graphs represent genes upregulated in the daytime (ZR0–ZT12), and the black bar graph represents genes upregulated in the night time (ZR12–ZT24).

Figure 5.
Expression of candidate clock-related genes over time in B. rapa. Heat map shows expression patterns of candidate clock-related genes. Candidate circadian-regulated genes have significantly different expression in B. rapa over time. The orthologs of Arabidopsis circadian-related genes identified based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways were screened in B. rapa (right). Dendrogram: hierarchical clustering of unigenes based on their expression pattern similarities. Gene ID numbers of the B. rapa sequence (Phytozome V9.0) (left). ZT0–24, time points after the light-on time (bottom).
Figure 5.
Expression of candidate clock-related genes over time in B. rapa. Heat map shows expression patterns of candidate clock-related genes. Candidate circadian-regulated genes have significantly different expression in B. rapa over time. The orthologs of Arabidopsis circadian-related genes identified based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways were screened in B. rapa (right). Dendrogram: hierarchical clustering of unigenes based on their expression pattern similarities. Gene ID numbers of the B. rapa sequence (Phytozome V9.0) (left). ZT0–24, time points after the light-on time (bottom).

Figure 6.
Circadian clock-related genes conserved in B. rapa. On the Arabidopsis clock network, all orthologous genes of B. rapa overlapped. The Arabidopsis clock network was adapted and modified from Lou et al. [41] and Hsu and Harmer [57]. Regulatory relationships in other networks are extrapolated from the Arabidopsis network rather than being derived empirically. Arrowheads and t-shaped arrows indicate positive and negative regulatory relationships, respectively. Yellow characters represent genes with free-running periods under continuous light. Colored tiles in bars indicate expression levels over 1 day. Expression levels decrease as the color changes from dark orange to white.
Figure 6.
Circadian clock-related genes conserved in B. rapa. On the Arabidopsis clock network, all orthologous genes of B. rapa overlapped. The Arabidopsis clock network was adapted and modified from Lou et al. [41] and Hsu and Harmer [57]. Regulatory relationships in other networks are extrapolated from the Arabidopsis network rather than being derived empirically. Arrowheads and t-shaped arrows indicate positive and negative regulatory relationships, respectively. Yellow characters represent genes with free-running periods under continuous light. Colored tiles in bars indicate expression levels over 1 day. Expression levels decrease as the color changes from dark orange to white.

Figure 7.
Expression pattern of glucosinolate-related genes in B. rapa. Heatmap shows significant differentially expressed candidate genes involved in the glucosinolate pathway in B. rapa [61]. Dendrogram: hierarchical clustering of unigenes based on their expression pattern similarities. Gene sequence IDs of the B. rapa (Phytozome V9.0) (left). ZT0–24: time points after the light-on time.
Figure 7.
Expression pattern of glucosinolate-related genes in B. rapa. Heatmap shows significant differentially expressed candidate genes involved in the glucosinolate pathway in B. rapa [61]. Dendrogram: hierarchical clustering of unigenes based on their expression pattern similarities. Gene sequence IDs of the B. rapa (Phytozome V9.0) (left). ZT0–24: time points after the light-on time.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Statistical analysis of Brassica rapa RNA sequencing (RNAseq) reads. ZT: zeitgeber Time.
| Index | Sample Name | Num. of Raw Reads | Num. of Cleaned Reads | Mean Length of Cleaned Reads (bp) | Num. of Mapped Reads to Brassica rapa Transcripts | Percent of Mapped Reads |
|---|---|---|---|---|---|---|
| 1 | ZT0_rep1 | 102,435,614 | 102,408,172 | 98.25 | 73,819,707 | 72.08% |
| 2 | ZT0_rep2 | 96,521,352 | 96,496,334 | 98.28 | 69,622,768 | 72.15% |
| 3 | ZT4_rep1 | 87,345,158 | 87,321,395 | 98.13 | 63,413,638 | 72.62% |
| 4 | ZT4_rep2 | 100,754,314 | 100,727,654 | 98.21 | 73,690,375 | 73.16% |
| 5 | ZT8_rep1 | 97,511,686 | 97,485,539 | 98.24 | 69,267,392 | 71.05% |
| 6 | ZT8_rep2 | 96,598,065 | 96,572,858 | 98.23 | 70,045,246 | 72.53% |
| 7 | ZT12_rep1 | 95,002,635 | 94,977,251 | 98.25 | 69,348,349 | 73.02% |
| 8 | ZT12_rep2 | 96,254,940 | 96,229,049 | 98.28 | 70,228,180 | 72.98% |
| 9 | ZT16_rep1 | 92,587,264 | 92,562,453 | 98.27 | 66,796,081 | 72.16% |
| 10 | ZT16_rep2 | 96,368,659 | 96,342,993 | 98.25 | 68,985,090 | 71.6% |
| 11 | ZT20_rep1 | 98,509,742 | 98,483,494 | 98.25 | 70,150,697 | 71.23% |
| 12 | ZT20_rep2 | 94,190,011 | 94,164,614 | 98.29 | 67,805,050 | 72.01% |
| 13 | ZT24_rep1 | 100,934,518 | 100,909,158 | 98.26 | 72,773,028 | 72.12% |
| 14 | ZT24_rep2 | 108,544,184 | 108,517,103 | 98.28 | 77,988,632 | 71.87% |
Table 2.
The lists of orthologous genes included in circadian clock pathway.
| AGI | Alias | Description | ID | PER | LAG | AMP |
|---|---|---|---|---|---|---|
| AT1G09570.1 | FHY2,FRE1,HY8,PHYA | phytochrome A | Bra020013 | 24.00 | 2.00 | 3.57 |
| Bra031672 | 18.00 | NA | 0.06 | |||
| AT2G18790.1 | HY3,OOP1,PHYB | phytochrome B | Bra001650 | 24.00 | 13.00 | 0.07 |
| Bra022192 | 22.00 | 7.15 | 3.58 | |||
| AT4G08920.1 | ATCRY1,BLU1,CRY1,HY4,OOP2 | cryptochrome 1 | Bra037880 | 24.00 | 2.00 | 58.64 |
| AT1G04400.1 | AT-PHH1,ATCRY2,CRY2,FHA,PHH1 | cryptochrome 2 | Bra015313 | 12.00 | 0.00 | 9.70 |
| Bra030568 | 10.00 | NA | 0.00 | |||
| AT2G32950.1 | ATCOP1,COP1,DET340,EMB168,FUS1 | transducin/WD40 repeat-like superfamily protein | Bra005541 | 20.00 | 7.00 | 5.27 |
| Bra021818 | 18.00 | NA | 0.04 | |||
| AT1G09530.1 | PAP3,PIF3,POC1 | phytochrome interacting factor 3 | Bra020017 | 24.00 | 5.00 | 3.75 |
| Bra031668 | 24.00 | 5.00 | 2.83 | |||
| AT1G22770.1 | FB,GI | gigantea protein (GI) | Bra024536 | 24.00 | 8.00 | 8.20 |
| AT2G25930.1 | ELF3,PYK20 | hydroxyproline-rich glycoprotein family protein | Bra007774 | 24.00 | 18.00 | 4.14 |
| Bra034284 | 20.00 | 0.00 | 2.22 | |||
| AT2G40080.1 | ELF4 | Protein of unknown function (DUF1313) | Bra000165 | 24.00 | 13.00 | 13.00 |
| Bra004991 | 24.00 | 14.00 | 19.58 | |||
| Bra017035 | 20.80 | 12.58 | 20.58 | |||
| AT5G02840.1 | LCL1(RVE4) | LHY/CCA1-like 1 | Bra005751 | 24.00 | 0.00 | 14.20 |
| Bra005754 | 22.00 | 19.07 | 0.10 | |||
| Bra009562 | 22.00 | 0.00 | 10.42 | |||
| AT3G09600.1 | RVE8 | homeodomain-like superfamily protein | Bra029778 | 24.00 | 2.00 | 19.94 |
| Bra034074 | 24.00 | 2.00 | 10.72 | |||
| AT3G46640.1 | LUX,PCL1 | homeodomain-like superfamily protein | Bra018204 | 24.00 | 16.00 | 4.89 |
| Bra033809 | 22.00 | 14.12 | 8.36 | |||
| AT2G18915.2 | ADO2,LKP2 | LOV KELCH protein 2 | Bra038830 | 24.00 | 3.00 | 1.58 |
| Bra038831 | 24.00 | 2.00 | 1.29 | |||
| Bra038832 | 22.67 | 11.96 | 5.81 | |||
| AT1G01060.1 AT1G01060.4 | LHY,LHY1 | homeodomain-like superfamily protein | Bra030496 | 24.00 | 1.00 | 34.47 |
| Bra033291 | 24.00 | 0.00 | 37.84 | |||
| AT2G46830.1 | CCA1 | circadian clock associated 1 | Bra004503 | 24.00 | 2.00 | 33.20 |
| AT5G61380.1 | APRR1,AtTOC1,PRR1,TOC1 | CCT motif -containing response regulator protein | Bra012964 | 22.67 | 11.96 | 15.09 |
| Bra035933 | 24.00 | 12.00 | 6.58 | |||
| AT5G60100.1 AT5G60100.2 | APRR3,PRR3 | pseudo-response regulator 3 | Bra002512 | 20.00 | 10.00 | 22.85 |
| Bra020263 | 18.00 | NA | NA | |||
| AT5G24470.1 | APRR5,PRR5 | pseudo-response regulator 5 | Bra009768 | 20.00 | 10.00 | 1.19 |
| Bra029407 | 20.00 | 10.00 | 3.28 | |||
| Bra036517 | 20.00 | 8.00 | 2.12 | |||
| AT5G02810.1 | APRR7,PRR7 | pseudo-response regulator 7 | Bra009565 | 24.00 | 8.00 | 25.25 |
| Bra028861 | 22.00 | 10.08 | 40.83 | |||
| AT2G46790.1 | APRR9,PRR9,TL1 | pseudo-response regulator 9 | Bra004507 | 24.00 | 6.00 | 0.11 |
| Bra040484 | 22.67 | 0.62 | 12.55 |
PER: period; LAG: peak time; AMP: amplitude from JCK-CYCLE.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, J.A.; Shim, D.; Kumari, S.; Jung, H.-e.; Jung, K.-H.; Jeong, H.; Kim, W.-Y.; Lee, S.I.; Jeong, M.-J. Transcriptome Analysis of Diurnal Gene Expression in Chinese Cabbage. Genes 2019, 10, 130. https://doi.org/10.3390/genes10020130
AMA Style
Kim JA, Shim D, Kumari S, Jung H-e, Jung K-H, Jeong H, Kim W-Y, Lee SI, Jeong M-J. Transcriptome Analysis of Diurnal Gene Expression in Chinese Cabbage. Genes. 2019; 10(2):130. https://doi.org/10.3390/genes10020130
Chicago/Turabian StyleKim, Jin A., Donghwan Shim, Shipra Kumari, Ha-eun Jung, Ki-Hong Jung, Heesu Jeong, Woe-Yeon Kim, Soo In Lee, and Mi-Jeong Jeong. 2019. "Transcriptome Analysis of Diurnal Gene Expression in Chinese Cabbage" Genes 10, no. 2: 130. https://doi.org/10.3390/genes10020130
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.