Pravastatin and Gemfibrozil Modulate Differently Hepatic and Colonic Mitochondrial Respiration in Tissue Homogenates from Healthy Rats

 and
and 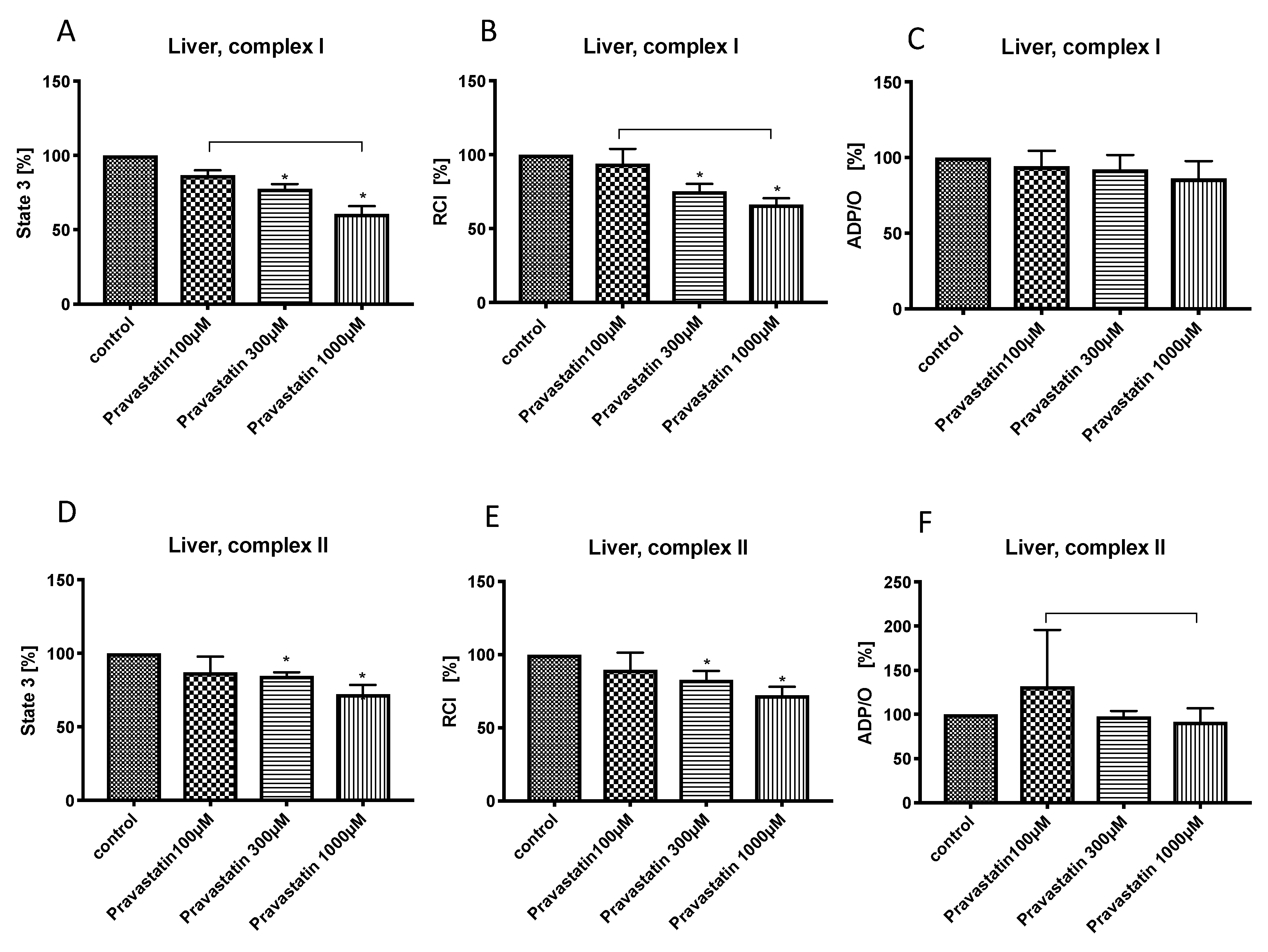{kind=link}
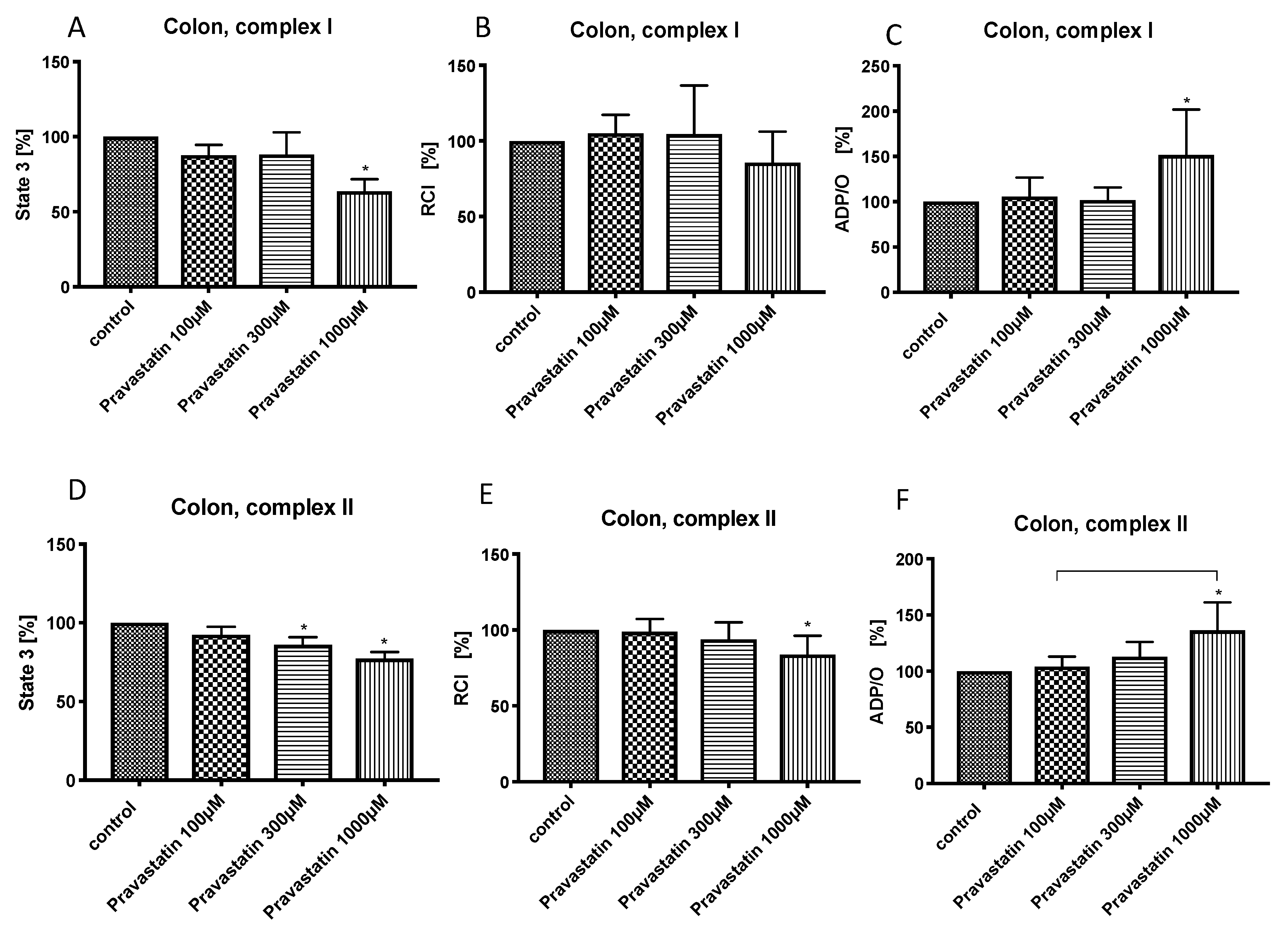{kind=link}
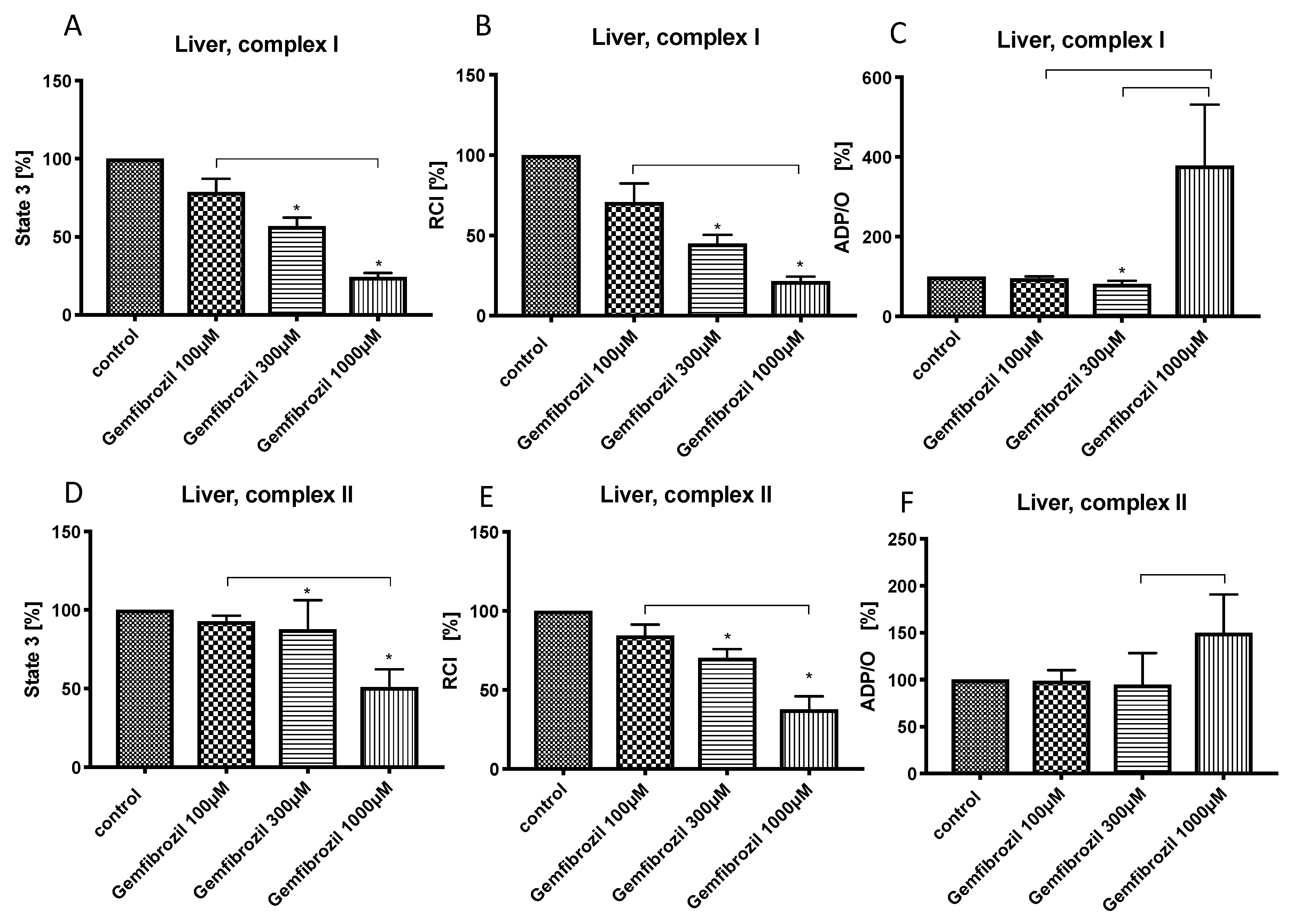{kind=link}
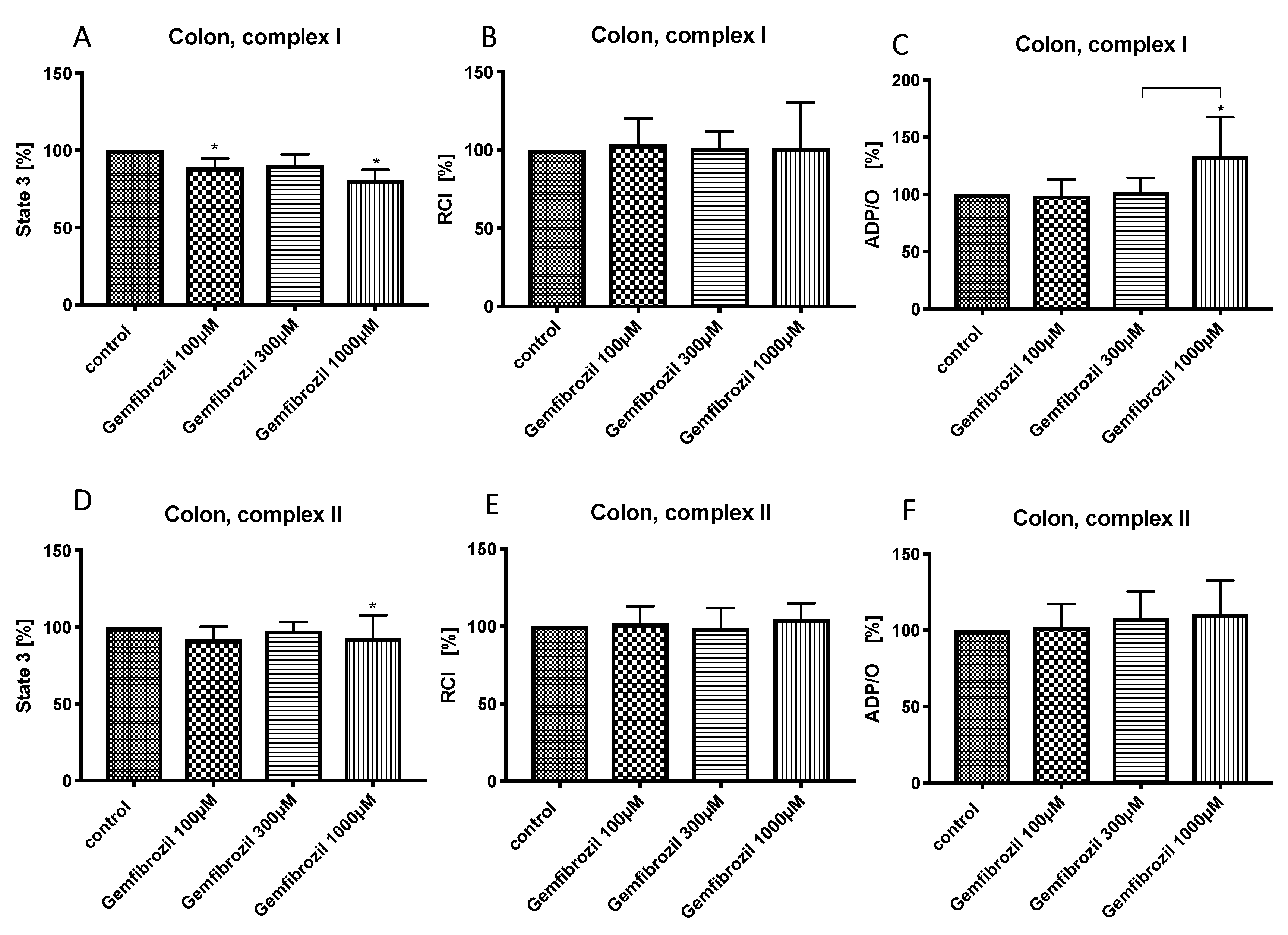{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals:
2.2. Preparation of Liver and Colon Homogenates
2.3. Measurement of Mitochondrial Respiratory Rates
2.3.1. Mitochondrial State 2 Respiration
2.3.2. Mitochondrial State 3 Respiration
2.3.3. General Conditions
2.3.4. Quality Control for the Preparation Procedure
2.3.5. Experimental Conditions
2.4. Statistical Analysis
3. Results
3.1. Effects of Pravastatin on Hepatic Mitochondrial Respiration
3.2. Effects of Pravastatin on Colonic Mitochondrial Respiration
3.3. Effects of Gemfibrozil on Hepatic Mitochondrial Respiration
3.4. Effects of Gemfibrozil on Colonic Mitochondrial Respiration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Godoy, J.C.; Niesman, I.R.; Busija, A.R.; Kassan, A.; Schilling, J.M.; Schwarz, A.; Alvarez, E.A.; Dalton, N.D.; Drummond, J.C.; Roth, D.M.; et al. Atorvastatin, but not pravastatin, inhibits cardiac Akt/mTOR signaling and disturbs mitochondrial ultrastructure in cardiac myocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 1209–1225. [Google Scholar] [CrossRef]
- Terblanche, M.; Almog, Y.; Rosenson, R.S.; Smith, T.S.; Hackam, D.G. Statins and sepsis: Multiple modifications at multiple levels. Lancet Infect. Dis. 2007, 7, 358–368. [Google Scholar] [CrossRef]
- Piepoli, M.F.; Hoes, A.W.; Agewall, S.; Albus, C.; Brotons, C.; Catapano, A.L.; Cooney, M.-T.; Corrà, U.; Cosyns, B.; Deaton, C.; et al. 2016 European Guidelines on cardiovascular disease prevention in clinical practice: The Sixth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (constituted by representatives of 10 societies and by invited experts) Developed with the special contribution of the European Association for Cardiovascular Prevention & Rehabilitation (EACPR). Eur. Heart J. 2016, 37, 2315–2381. [Google Scholar] [PubMed]
- Wiel, E.; Lebuffe, G.; Robin, E.; Gasan, G.; Corseaux, D.; Tavernier, B.; Jude, B.; Bordet, R.; Vallet, B. Pretreatment with peroxysome proliferator-activated receptor alpha agonist fenofibrate protects endothelium in rabbit Escherichia coli endotoxin-induced shock. Intensive Care Med. 2005, 31, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Goya, K.; Sumitani, S.; Xu, X.; Kitamura, T.; Yamamoto, H.; Kurebayashi, S.; Saito, H.; Kouhara, H.; Kasayama, S.; Kawase, I. Peroxisome proliferator-activated receptor alpha agonists increase nitric oxide synthase expression in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.; Takemura, T.; Eriksson, P.; Hamsten, A. Effects of Fibrate Compounds on Expression of Plasminogen Activator Inhibitor-1 by Cultured Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1577–1581. [Google Scholar] [CrossRef]
- Frost, R.J.; Otto, C.; Geiss, H.C.; Schwandt, P.; Parhofer, K.G. Effects of atorvastatin versus fenofibrate on lipoprotein profiles, low-density lipoprotein subfraction distribution, and hemorheologic parameters in type 2 diabetes mellitus with mixed hyperlipoproteinemia. Am. J. Cardiol. 2001, 87, 44–48. [Google Scholar] [CrossRef]
- Davidson, M.H. Statin/fibrate combination in patients with metabolic syndrome or diabetes: Evaluating the risks of pharmacokinetic drug interactions. Expert Opin. Drug Saf. 2006, 5, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Hedrington, M.S.; Davis, S.N. Peroxisome proliferator-activated receptor alpha-mediated drug toxicity in the liver. Expert Opin. Drug Metab. Toxicol. 2018, 14, 671–677. [Google Scholar] [CrossRef]
- Scatena, R.; Bottoni, P.; Vincenzoni, F.; Messana, I.; Martorana, G.E.; Nocca, G.; De Sole, P.; Maggiano, N.; Castagnola, M.; Giardina, B. Bezafibrate Induces a Mitochondrial Derangement in Human Cell Lines: A PPAR-Independent Mechanism for a Peroxisome Proliferator. Chem. Res. Toxicol. 2003, 16, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Mullen, P.J.; Zahno, A.; Lindinger, P.; Maseneni, S.; Felser, A.; Krähenbühl, S.; Brecht, K. Susceptibility to simvastatin-induced toxicity is partly determined by mitochondrial respiration and phosphorylation state of Akt. Biochim. Biophys. Acta 2011, 1813, 2079–2087. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, P.; Török, M.; Zahno, A.; Waldhauser, K.M.; Brecht, K.; Krähenbühl, S. Toxicity of statins on rat skeletal muscle mitochondria. Cell. Mol. Life Sci. CMLS 2006, 63, 2415–2425. [Google Scholar] [CrossRef] [PubMed]
- Päivä, H.; Thelen, K.M.; Van Coster, R.; Smet, J.; De Paepe, B.; Mattila, K.M.; Laakso, J.; Lehtimäki, T.; von Bergmann, K.; Lütjohann, D.; et al. High-dose statins and skeletal muscle metabolism in humans: A randomized, controlled trial. Clin. Pharmacol. Ther. 2005, 78, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Nadanaciva, S.; Rana, P.; Beeson, G.C.; Chen, D.; Ferrick, D.A.; Beeson, C.C.; Will, Y. Assessment of drug-induced mitochondrial dysfunction via altered cellular respiration and acidification measured in a 96-well platform. J. Bioenerg. Biomembr. 2012, 44, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.C.; Busanello, E.N.B.; de Oliveira, D.N.; Catharino, R.R.; Oliveira, H.C.F.; Vercesi, A.E. Coenzyme Q10 or Creatine Counteract Pravastatin-Induced Liver Redox Changes in Hypercholesterolemic Mice. Front. Pharmacol. 2018, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.S.; Chalasani, N. Lipid-lowering agents that cause drug-induced hepatotoxicity. Clin. Liver Dis. 2007, 11, 597–613. [Google Scholar] [CrossRef]
- Bouitbir, J.; Charles, A.-L.; Echaniz-Laguna, A.; Kindo, M.; Daussin, F.; Auwerx, J.; Piquard, F.; Geny, B.; Zoll, J. Opposite effects of statins on mitochondria of cardiac and skeletal muscles: A “mitohormesis” mechanism involving reactive oxygen species and PGC-1. Eur. Heart, J. 2012, 33, 1397–1407. [Google Scholar] [CrossRef]
- Brunmair, B.; Lest, A.; Staniek, K.; Gras, F.; Scharf, N.; Roden, M.; Nohl, H.; Waldhäusl, W.; Fürnsinn, C. Fenofibrate impairs rat mitochondrial function by inhibition of respiratory complex I. J. Pharmacol. Exp. Ther. 2004, 311, 109–114. [Google Scholar] [CrossRef]
- Zhou, S.; Wallace, K.B. The effect of peroxisome proliferators on mitochondrial bioenergetics. Toxicol. Sci. Off. J. Soc. Toxicol. 1999, 48, 82–89. [Google Scholar] [CrossRef]
- Zungu, M.; Young, M.E.; Stanley, W.C.; Essop, M.F. Chronic treatment with the peroxisome proliferator-activated receptor alpha agonist Wy-14,643 attenuates myocardial respiratory capacity and contractile function. Mol. Cell. Biochem. 2009, 330, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Andreux, P.A.; Houtkooper, R.H.; Auwerx, J. Pharmacological approaches to restore mitochondrial function. Nat. Rev. Drug Discov. 2013, 12, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Herminghaus, A.; Eberhardt, R.; Truse, R.; Schulz, J.; Bauer, I.; Picker, O.; Vollmer, C. Nitroglycerin and Iloprost Improve Mitochondrial Function in Colon Homogenate Without Altering the Barrier Integrity of Caco-2 Monolayers. Front. Med. 2018, 5, 291. [Google Scholar] [CrossRef] [PubMed]
- Herminghaus, A.; Buitenhuis, A.J.; Schulz, J.; Vollmer, C.; Scheeren, T.W.L.; Bauer, I.; Picker, O.; Truse, R. Propofol improves colonic but impairs hepatic mitochondrial function in tissue homogenates from healthy rats. Eur. J. Pharmacol. 2019, 853, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Herminghaus, A.; Papenbrock, H.; Eberhardt, R.; Vollmer, C.; Truse, R.; Schulz, J.; Bauer, I.; Weidinger, A.; Kozlov, A.V.; Stiban, J.; et al. Time-related changes in hepatic and colonic mitochondrial oxygen consumption after abdominal infection in rats. Intensive Care Med. Exp. 2019, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Cámara-Lemarroy, C.R.; Guzman-DE LA Garza, F.J.; Cordero-Perez, P.; Ibarra-Hernandez, J.M.; Muñoz-Espinosa, L.E.; Fernandez-Garza, N.E. Gemfibrozil attenuates the inflammatory response and protects rats from abdominal sepsis. Exp. Ther. Med. 2015, 9, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S. HMG CoA reductase inhibitor accelerates aging effect on diaphragm mitochondrial respiratory function in rats. Biochem. Mol. Biol. Int. 1998, 46, 923–931. [Google Scholar]
- Jeger, V.; Djafarzadeh, S.; Jakob, S.M.; Takala, J. Mitochondrial function in sepsis. Eur. J. Clin. Invest. 2013, 43, 532–542. [Google Scholar] [CrossRef]
- Siekmeier, R.; Gross, W.; März, W. Determination of pravastatin by high performance liquid chromatography. Int. J. Clin. Pharmacol. Ther. 2000, 38, 419–425. [Google Scholar] [CrossRef]
- McTavish, D.; Sorkin, E.M. Pravastatin. A review of its pharmacological properties and therapeutic potential in hypercholesterolaemia. Drugs 1991, 42, 65–89. [Google Scholar] [CrossRef]
- Hatanaka, T. Clinical pharmacokinetics of pravastatin: Mechanisms of pharmacokinetic events. Clin. Pharmacokinet. 2000, 39, 397–412. [Google Scholar] [CrossRef]
- Yamazaki, M.; Tokui, T.; Ishigami, M.; Sugiyama, Y. Tissue-selective uptake of pravastatin in rats: Contribution of a specific carrier-mediated uptake system. Biopharm. Drug Dispos. 1996, 17, 775–789. [Google Scholar] [CrossRef]
- Hatanaka, T.; Honda, S.; Sasaki, S.; Katayama, K.; Koizumi, T. Pharmacokinetic and pharmacodynamic evaluation for tissue-selective inhibition of cholesterol synthesis by pravastatin. J. Pharmacokinet. Biopharm. 1998, 26, 329–347. [Google Scholar] [CrossRef]
- Omar, M.A.; Wilson, J.P. FDA adverse event reports on statin-associated rhabdomyolysis. Ann. Pharmacother. 2002, 36, 288–295. [Google Scholar] [CrossRef]
- Vladutiu, G.D.; Simmons, Z.; Isackson, P.J.; Tarnopolsky, M.; Peltier, W.L.; Barboi, A.C.; Sripathi, N.; Wortmann, R.L.; Phillips, P.S. Genetic risk factors associated with lipid-lowering drug-induced myopathies. Muscle Nerve 2006, 34, 153–162. [Google Scholar] [CrossRef]
- Herminghaus, A.; Barthel, F.; Heinen, A.; Beck, C.; Vollmer, C.; Bauer, I.; Weidinger, A.; Kozlov, A.V.; Picker, O. Severity of polymicrobial sepsis modulates mitochondrial function in rat liver. Mitochondrion 2015, 24, 122–128. [Google Scholar] [CrossRef]
- Evans, J.R.; Forland, S.C.; Cutler, R.E. The effect of renal function on the pharmacokinetics of gemfibrozil. J. Clin. Pharmacol. 1987, 27, 994–1000. [Google Scholar] [CrossRef]
- Busse, K.H.; Hadigan, C.; Chairez, C.; Alfaro, R.M.; Formentini, E.; Kovacs, J.A.; Penzak, S.R. Gemfibrozil concentrations are significantly decreased in the presence of lopinavir-ritonavir. J. Acquir. Immune Defic. Syndr. 2009, 52, 235–239. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herminghaus, A.; Laser, E.; Schulz, J.; Truse, R.; Vollmer, C.; Bauer, I.; Picker, O. Pravastatin and Gemfibrozil Modulate Differently Hepatic and Colonic Mitochondrial Respiration in Tissue Homogenates from Healthy Rats. Cells 2019, 8, 983. https://doi.org/10.3390/cells8090983
Herminghaus A, Laser E, Schulz J, Truse R, Vollmer C, Bauer I, Picker O. Pravastatin and Gemfibrozil Modulate Differently Hepatic and Colonic Mitochondrial Respiration in Tissue Homogenates from Healthy Rats. Cells. 2019; 8(9):983. https://doi.org/10.3390/cells8090983
Chicago/Turabian StyleHerminghaus, Anna, Eric Laser, Jan Schulz, Richard Truse, Christian Vollmer, Inge Bauer, and Olaf Picker. 2019. "Pravastatin and Gemfibrozil Modulate Differently Hepatic and Colonic Mitochondrial Respiration in Tissue Homogenates from Healthy Rats" Cells 8, no. 9: 983. https://doi.org/10.3390/cells8090983
APA StyleHerminghaus, A., Laser, E., Schulz, J., Truse, R., Vollmer, C., Bauer, I., & Picker, O. (2019). Pravastatin and Gemfibrozil Modulate Differently Hepatic and Colonic Mitochondrial Respiration in Tissue Homogenates from Healthy Rats. Cells, 8(9), 983. https://doi.org/10.3390/cells8090983