Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
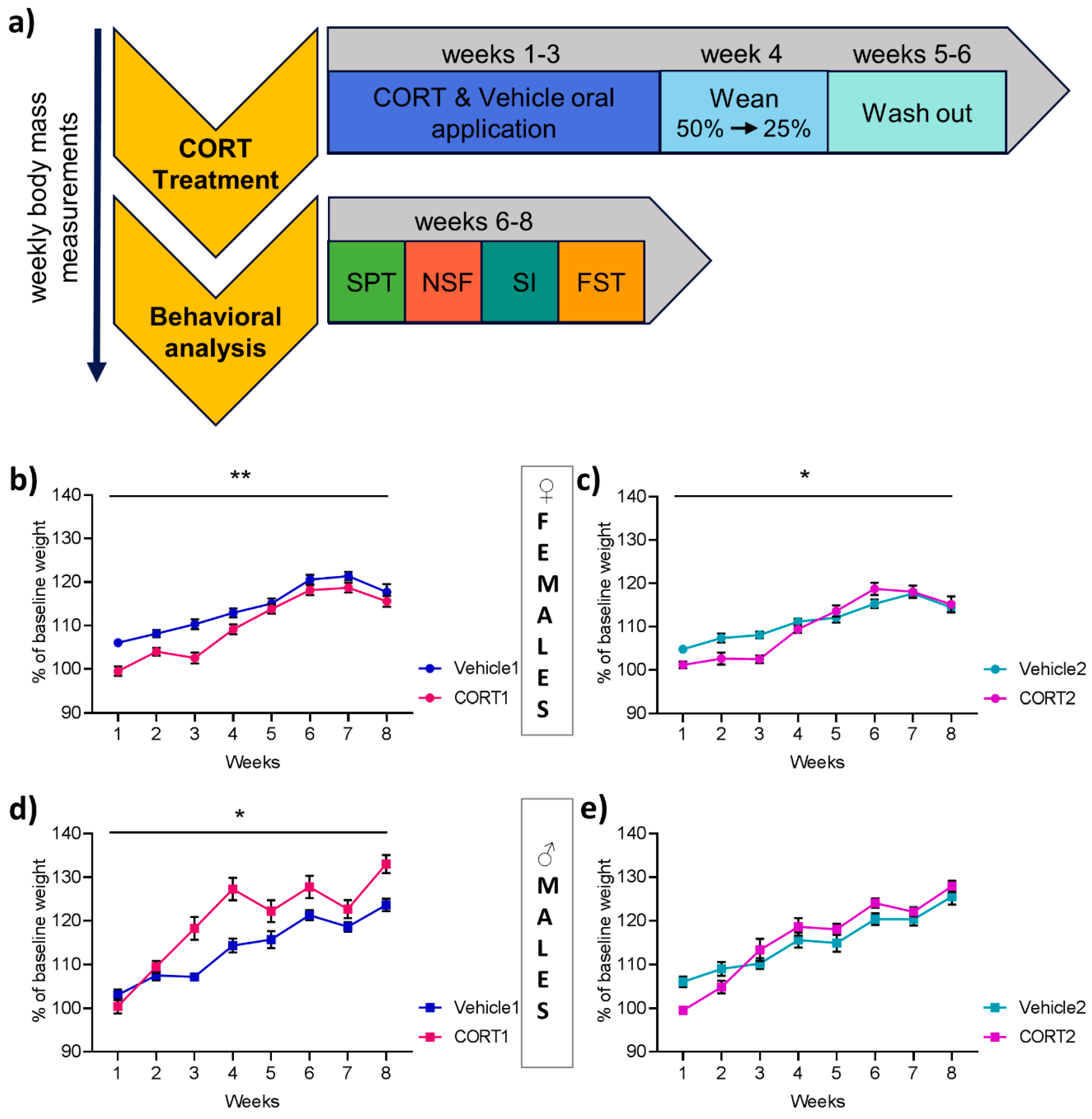
2.2. CORT Treatment
2.3. Behavioral Analysis
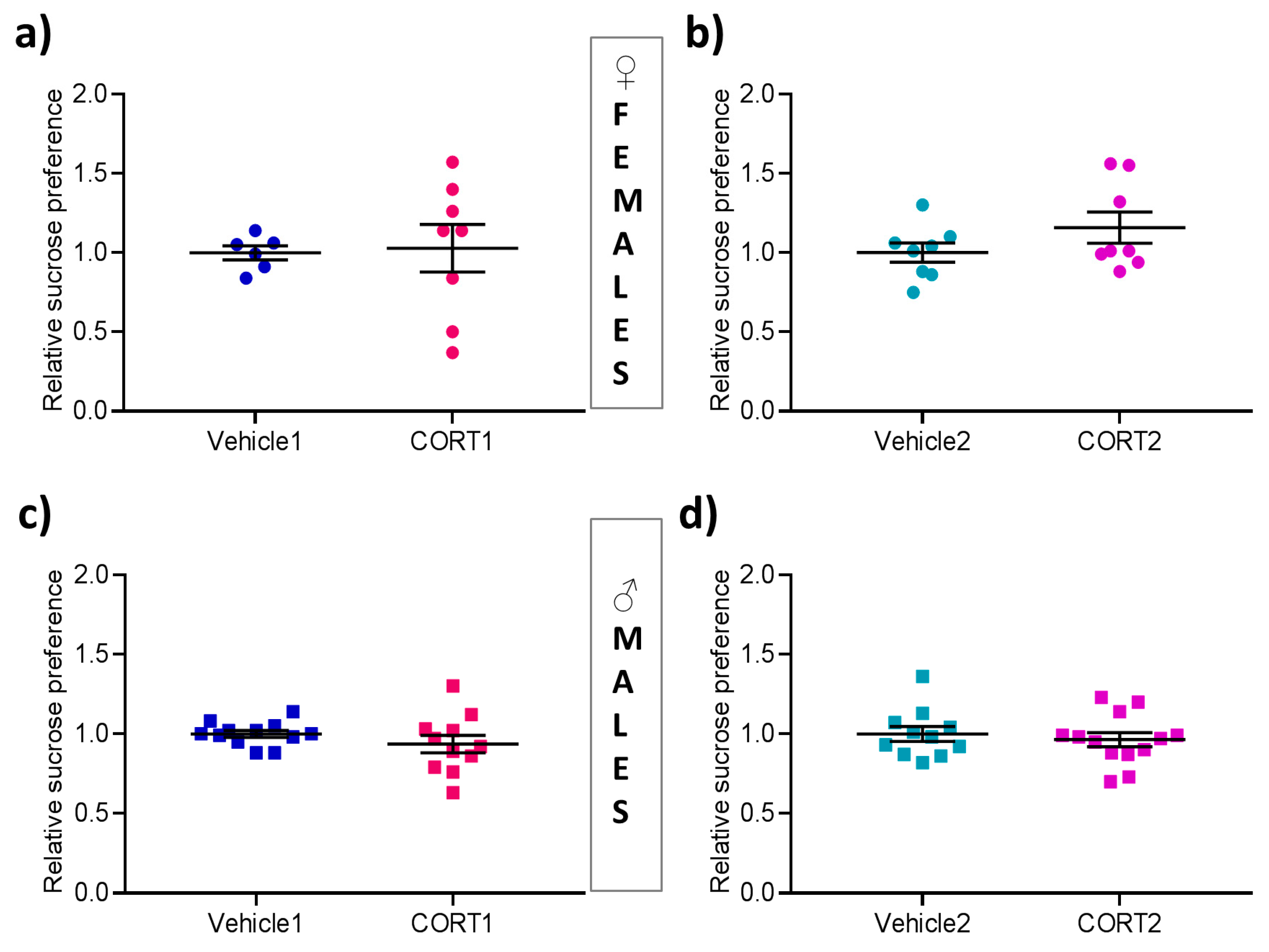
2.3.1. Sucrose Preference Test (SPT)
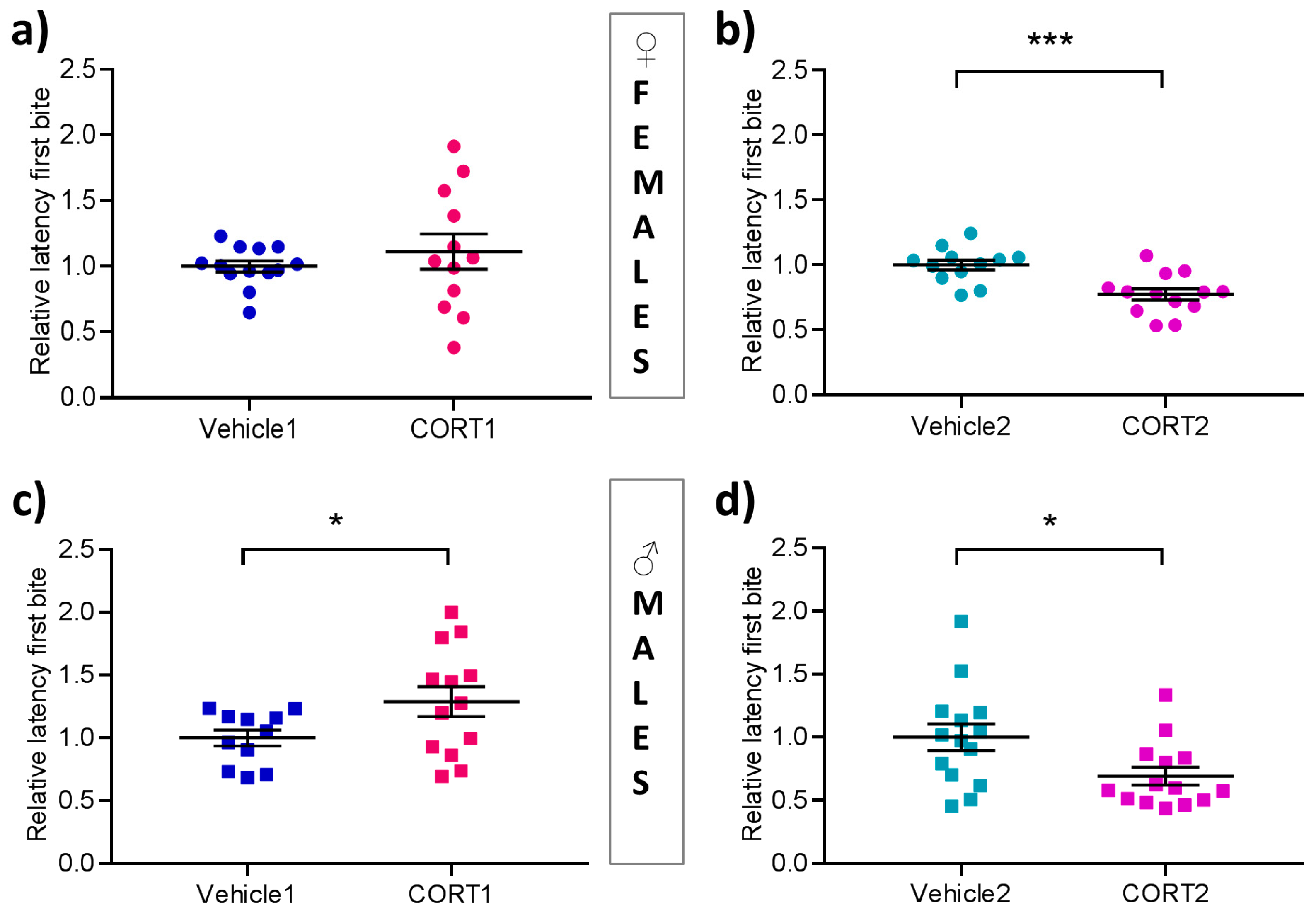
2.3.2. Novelty Suppressed Feeding (NSF)
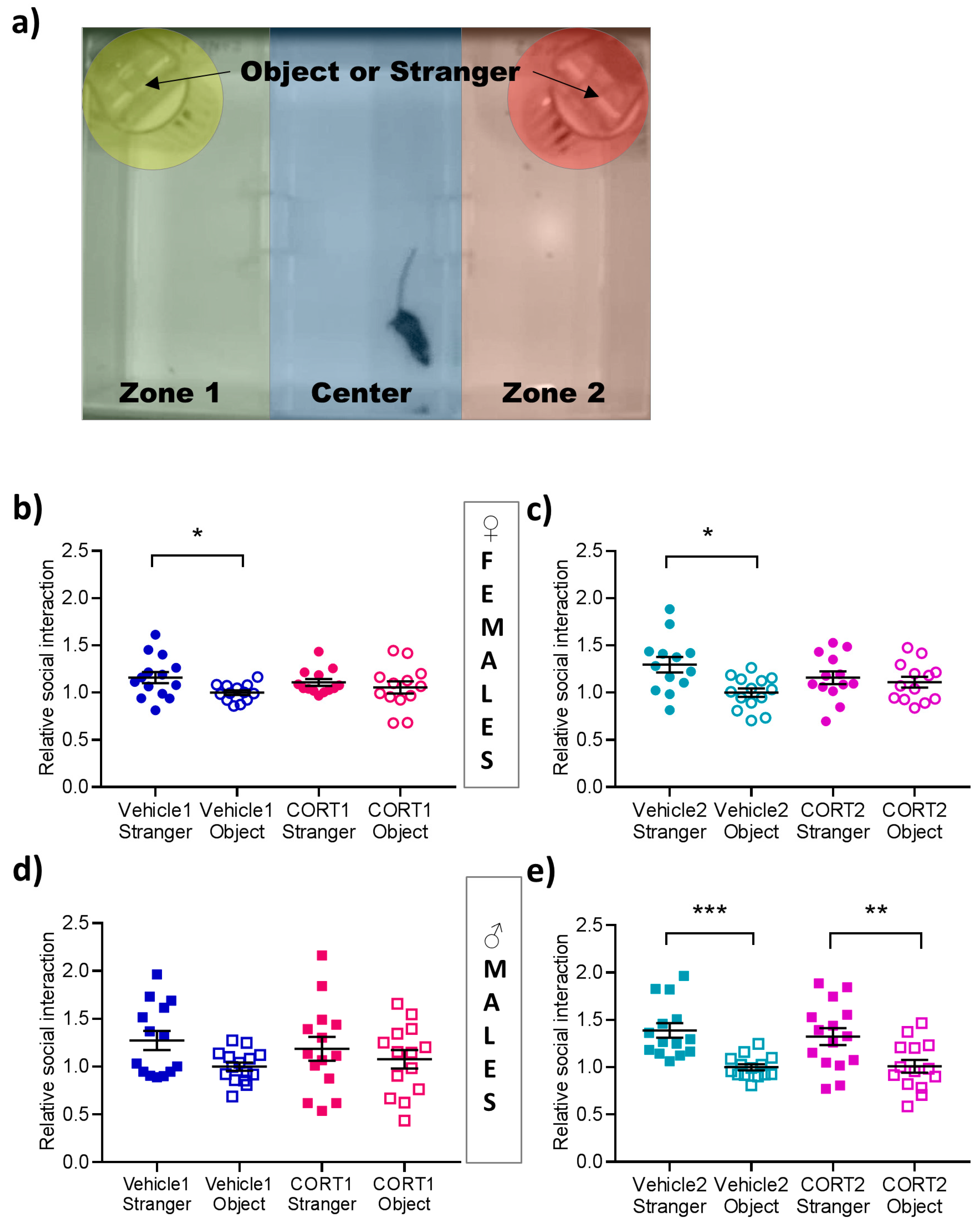
2.3.3. Social Interaction (SI)
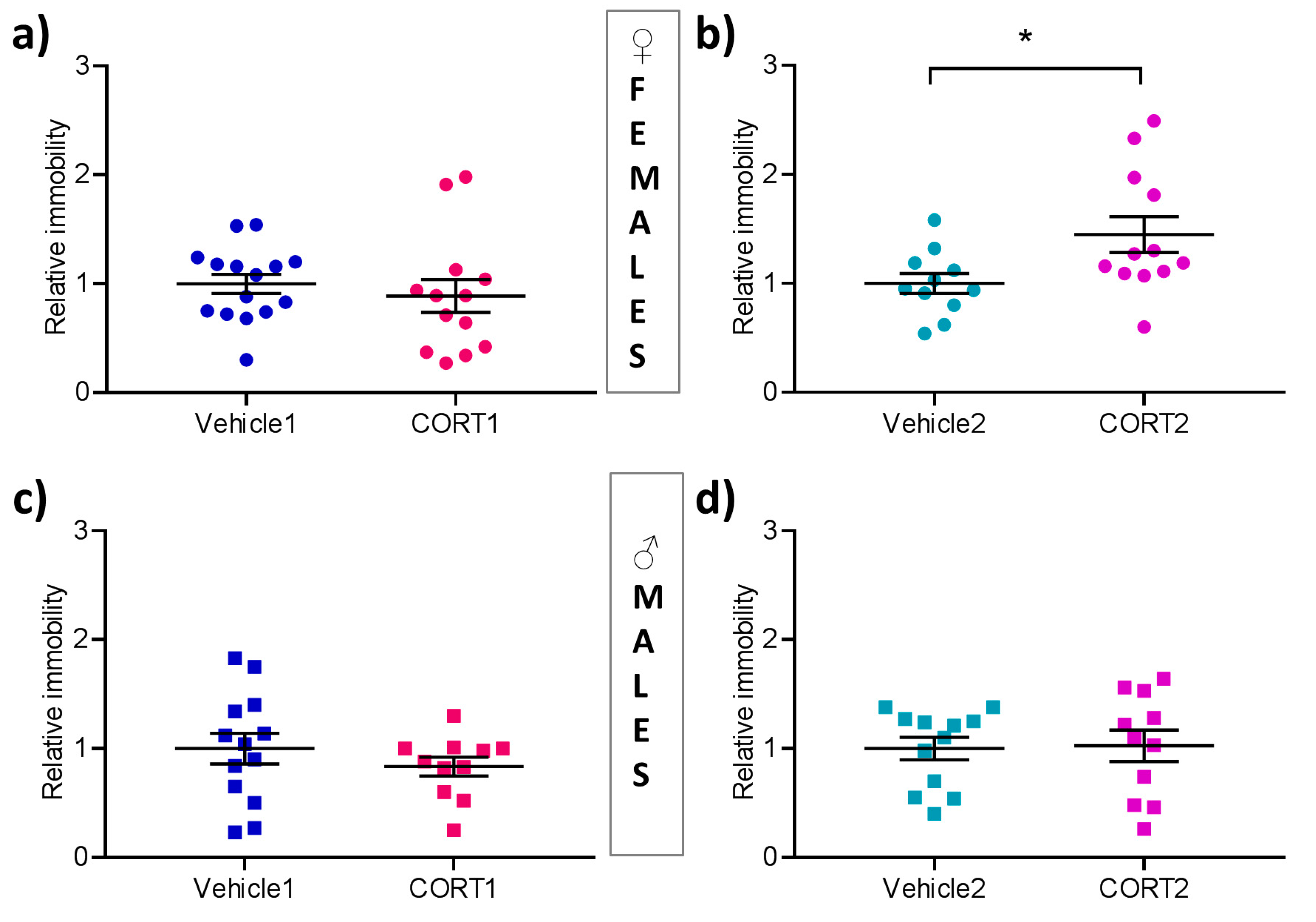
2.3.4. Forced Swim Test (FST)
2.4. Statistical Analysis
3. Results
3.1. Effect of CORT Treatment on Bodyweight Increase
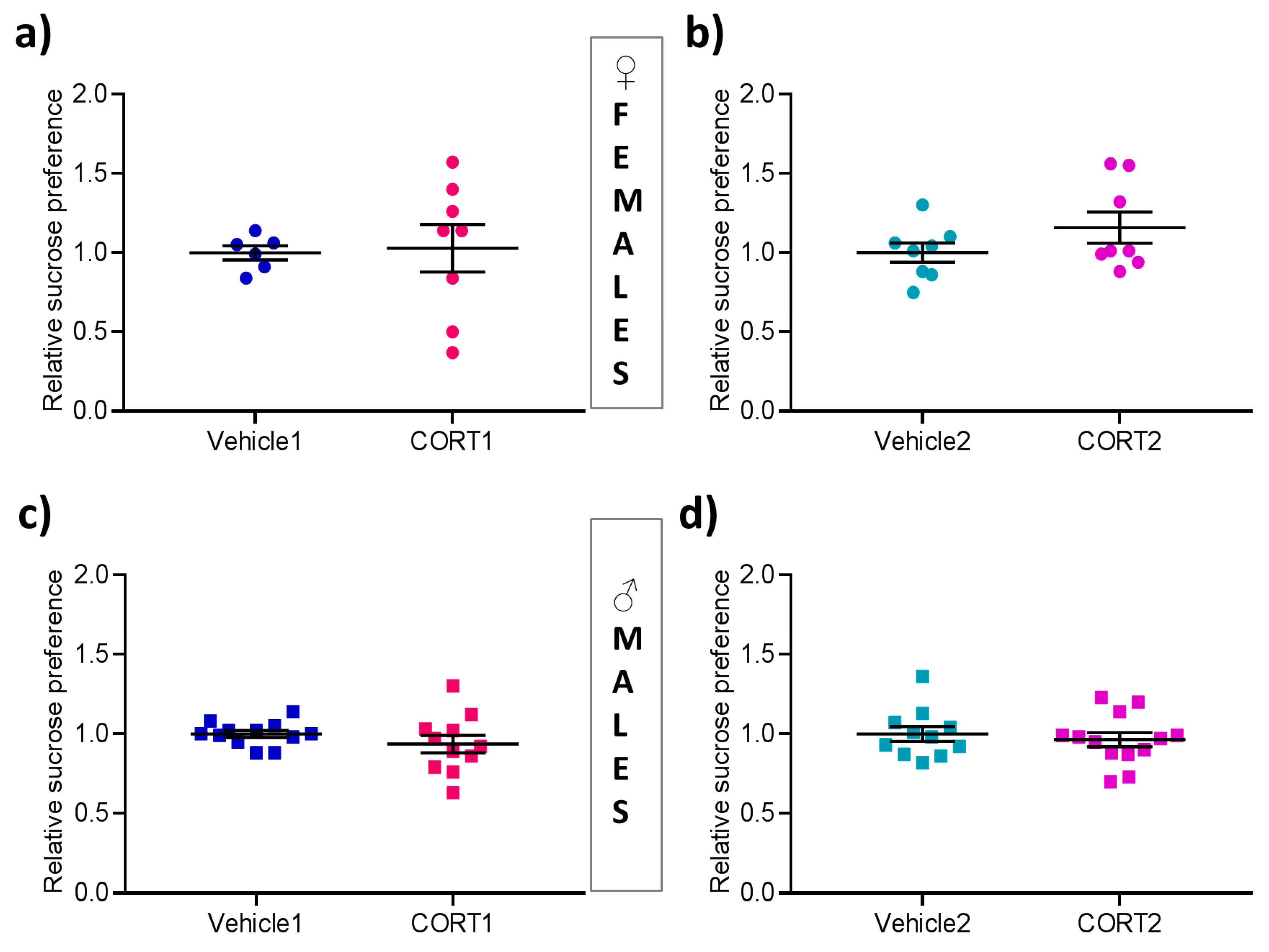
3.2. No Impact of CORT Treatment on Sucrose Preference
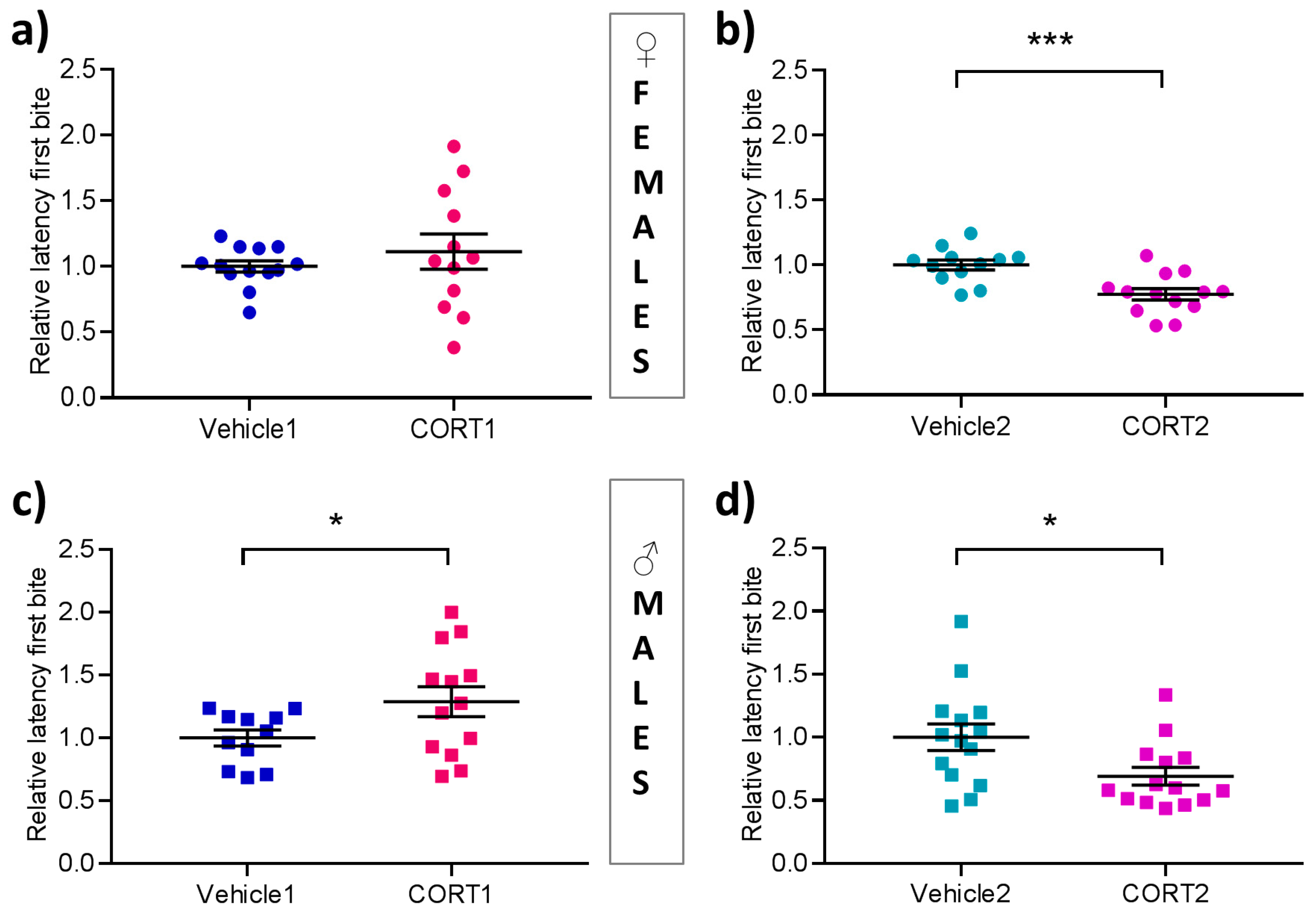
3.3. Altered Hyponeophagia in CORT Mice
3.4. Impaired Social Interaction in CORT Mice
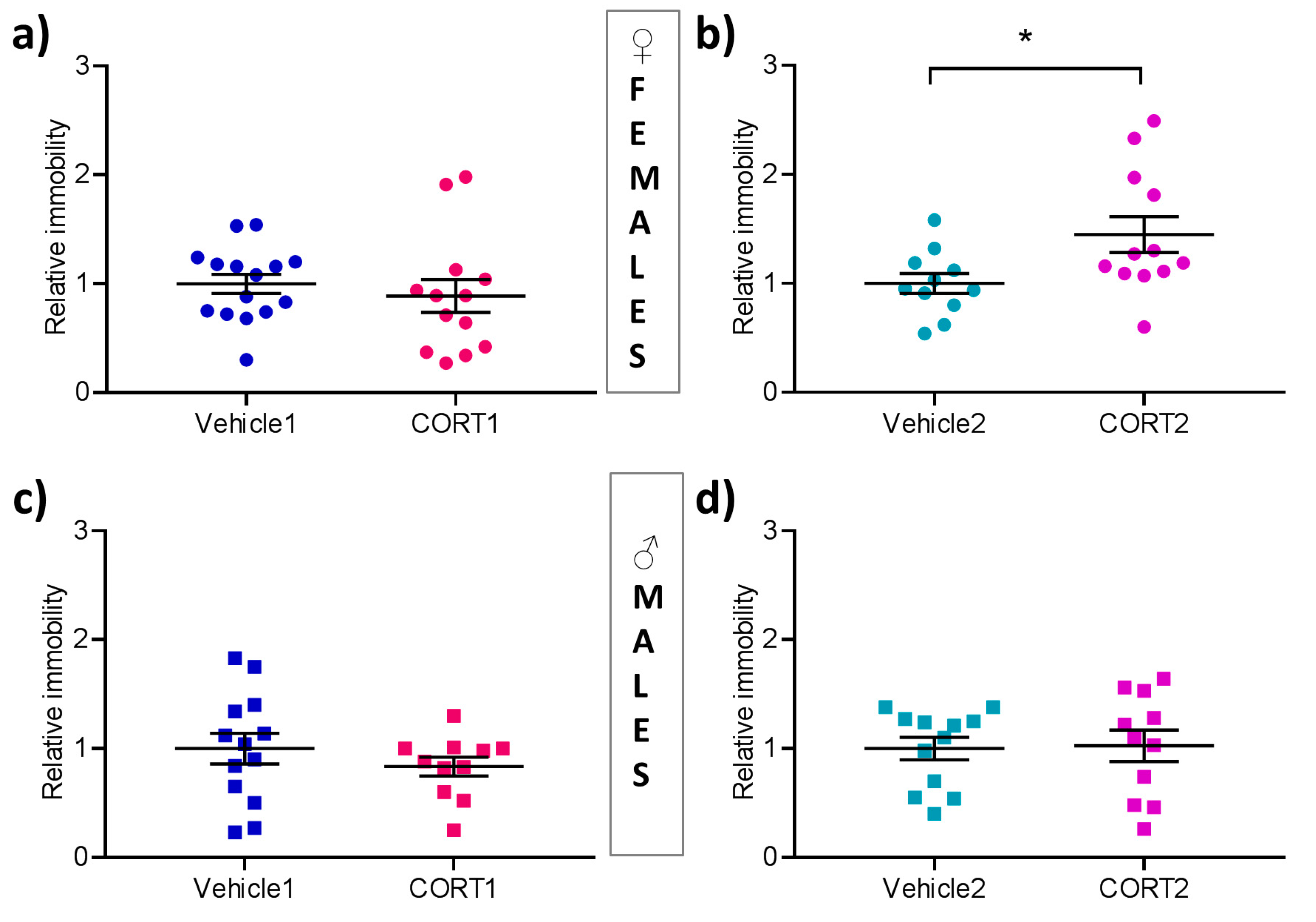
3.5. CORT2 Females Present with Increased Behavioral Despair
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WHO | Depression. Available online: http://www.who.int/mental_health/management/depression/en/ (accessed on 20 February 2019).
- American Psychiatric Association. DSM-5 Task Force. In Diagnostic and statistical manual of mental disorders: DSM-5, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 9780890425541. [Google Scholar]
- Pollak, D.D.; Rey, C.E.; Monje, F.J. Rodent models in depression research: Classical strategies and new directions. Ann. Med. 2010, 42, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.R. Why is depression more prevalent in women? J. Psychiatry Neurosci. 2015, 40, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Shansky, R.M. Are hormones a “female problem” for animal research? Science 2019, 364, 825–826. [Google Scholar] [CrossRef] [PubMed]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheggi, S.; De Montis, M.G.; Gambarana, C. Making Sense of Rodent Models of Anhedonia. Int. J. Neuropsychopharmacol. 2018, 21, 1049–1065. [Google Scholar] [CrossRef] [PubMed]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The Forced Swim Test as a Model of Depressive-like Behavior. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [PubMed]
- Samuels, B.; Hen, R. Novelty-suppressed feeding in the mouse. Mood Anxiety Relat. Phenotypes Mice: Charact. Using Behav. Testsvolume Ii 2011, 107–121. [Google Scholar]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal Relationship between Stressful Life Events and the Onset of Major Depression. AJP 1999, 156, 837–841. [Google Scholar] [CrossRef]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of Depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M.; Schatzberg, A.F. HPA Axis in Major Depression: Cortisol, Clinical Symptomatology, and Genetic Variation Predict Cognition. Mol. Psychiatry 2017, 22, 527–536. [Google Scholar] [CrossRef]
- Varghese, F.P.; Brown, E.S. The Hypothalamic-Pituitary-Adrenal Axis in Major Depressive Disorder: A Brief Primer for Primary Care Physicians. Prim. Care Companion J. Clin Psychiatry 2001, 3, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.-C.; Cao, X.; Das, M.; Zhu, X.-H.; Gao, T.-M. Behavioral animal models of depression. Neurosci. Bull. 2010, 26, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abelaira, H.M.; Réus, G.Z.; Quevedo, J. Animal models as tools to study the pathophysiology of depression. Revista brasileira de psiquiatria (São Paulo, Brazil: 1999) 2013, 35 (Suppl. 2), S112–S120. [Google Scholar] [CrossRef] [PubMed]
- Gourley, S.L.; Taylor, J.R. Recapitulation and reversal of a persistent depression-like syndrome in rodents. Curr Protoc Neurosci 2009, Chapter 9, Unit 9 32. 49, 9–32. [Google Scholar] [CrossRef] [PubMed]
- Gourley, S.L.; Kiraly, D.D.; Howell, J.L.; Olausson, P.; Taylor, J.R. Acute Hippocampal Brain-Derived Neurotrophic Factor Restores Motivational and Forced Swim Performance After Corticosterone. Biological Psychiatry 2008, 64, 884–890. [Google Scholar] [CrossRef] [Green Version]
- Gourley, S.L.; Wu, F.J.; Kiraly, D.D.; Ploski, J.E.; Kedves, A.T.; Duman, R.S.; Taylor, J.R. Regionally-specific regulation of pERK1/2 MAP kinase in a model of antidepressant-sensitive chronic depression. Biol Psychiatry 2008, 63, 353–359. [Google Scholar] [CrossRef] [PubMed]
- David, D.J.; Samuels, B.A.; Rainer, Q.; Wang, J.-W.W.; Marsteller, D.; Mendez, I.; Drew, M.; Craig, D.A.; Guiard, B.P.; Guilloux, J.-P.P.; et al. Neurogenesis-Dependent and -Independent Effects of Fluoxetine in an Animal Model of Anxiety/Depression. Neuron 2009, 62, 479–493. [Google Scholar] [CrossRef] [Green Version]
- Monje, F.J.; Cabatic, M.; Divisch, I.; Kim, E.-J.; Herkner, K.R.; Binder, B.R.; Pollak, D.D. Constant darkness induces IL-6-dependent depression-like behavior through the NF-κB signaling pathway. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 9075–9083. [Google Scholar] [CrossRef]
- Savalli, G.; Diao, W.; Schulz, S.; Todtova, K.; Pollak, D.D. Diurnal oscillation of amygdala clock gene expression and loss of synchrony in a mouse model of depression. Int. J. Neuropsychopharmacol. 2015, 18. [Google Scholar] [CrossRef]
- Britton, D.R.; Britton, K.T. A sensitive open field measure of anxiolytic drug activity. Pharmacol. Biochem. Behav. 1981, 15, 577–582. [Google Scholar] [CrossRef]
- Mineur, Y.S.; Picciotto, M.R.; Sanacora, G. Antidepressant-Like Effects of Ceftriaxone in Male C57BL/6J Mice. Biol. Psychiatry 2007, 61, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.; Fernando, P.; Cicvaric, A.; Berger, A.; Pollak, A.; Monje, F.J.; Pollak, D.D. Long-term effects of maternal immune activation on depression-like behavior in the mouse. Translational psychiatry 2014, 4, e363. [Google Scholar] [CrossRef] [PubMed]
- Pollak, D.D.; Monje, F.J.; Zuckerman, L.; Denny, C.A.; Drew, M.R.; Kandel, E.R. An animal model of a behavioral intervention for depression. Neuron 2008, 60, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Stopa, L.R.S.; de Souza, C.F.; Santos, G.F.; Martins, A.B.; Ferreira, R.N.; de Andrade, F.G.; Leite, C.M.; Zaia, D.A.M.; Zaia, C.T.B.V.; Uchoa, E.T. Sex differences in glucocorticoids-induced anabolic effects in rats. Physiol. Behav. 2019, 209, 112587. [Google Scholar] [CrossRef] [PubMed]
- Van Donkelaar, E.L.; Vaessen, K.R.D.; Pawluski, J.L.; Sierksma, A.S.; Blokland, A.; Cañete, R.; Steinbusch, H.W.M. Long-Term Corticosterone Exposure Decreases Insulin Sensitivity and Induces Depressive-Like Behaviour in the C57BL/6NCrl Mouse. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Mekiri, M.; Gardier, A.M.; David, D.J.; Guilloux, J.-P. Chronic corticosterone administration effects on behavioral emotionality in female c57bl6 mice. Exp. Clin. Psychopharmacol. 2017, 25, 94–104. [Google Scholar] [CrossRef]
- Varholick, J.A.; Bailoo, J.D.; Palme, R.; Würbel, H. Phenotypic variability between Social Dominance Ranks in laboratory mice. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bartolomucci, A.; Palanza, P.; Sacerdote, P.; Ceresini, G.; Chirieleison, A.; Panerai, A.E.; Parmigiani, S. Individual housing induces altered immuno-endocrine responses to psychological stress in male mice. Psychoneuroendocrinology 2003, 28, 540–558. [Google Scholar] [CrossRef]
- Bundgaard, C.J.; Kalliokoski, O.; Abelson, K.S.; Hau, J. Acclimatization of Mice to Different Cage Types and Social Groupings with Respect to Fecal Secretion of IgA and Corticosterone Metabolites. In Vivo 2012, 26, 883–888. [Google Scholar]
- Moy, S.S.; Nadler, J.J.; Perez, A.; Barbaro, R.P.; Johns, J.M.; Magnuson, T.R.; Piven, J.; Crawley, J.N. Sociability and preference for social novelty in five inbred strains: An approach to assess autistic-like behavior in mice. Genesbrain Behav. 2004, 3, 287–302. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of Social Interaction Behaviors. J. Vis. Exp 2011. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, K.L.; Merriweather, M.Y.; Yuva-Paylor, L.A.; Paylor, R. The use of behavioral test batteries: Effects of training history. Physiol. Behav. 2001, 73, 705–717. [Google Scholar] [CrossRef]
- Dalla, C.; Shors, T.J. Sex differences in learning processes of classical and operant conditioning. Physiol. Behav. 2009, 97, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crusio, W.E. Troublesome variability in mouse studies. Nat. Neurosci. 2009, 12, 1075. [Google Scholar] [CrossRef] [PubMed]
- Kappel, S.; Hawkins, P.; Mendl, M.T. To Group or Not to Group? Good Practice for Housing Male Laboratory Mice. Anim. (Basel) 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- An, X.-L.; Zou, J.-X.; Wu, R.-Y.; Yang, Y.; Tai, F.-D.; Zeng, S.-Y.; Jia, R.; Zhang, X.; Liu, E.-Q.; Broders, H. Strain and sex differences in anxiety-like and social behaviors in C57BL/6J and BALB/cJ mice. Exp. Anim. 2011, 60, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Balcombe, J.P. Laboratory environments and rodents’ behavioural needs: a review. Lab Anim 2006, 40, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, X.-Y.; Zhu, Q.-B.; Li, J.; Shi, L.-G.; Wu, J.-L.; Zhang, Q.-J.; Huang, M.-L.; Bao, A.-M. Sex differences in the stress response in SD rats. Behav. Brain Res. 2015, 284, 231–237. [Google Scholar] [CrossRef]
- DeSantis, S.M.; Baker, N.L.; Back, S.E.; Spratt, E.; Ciolino, J.D.; Maria, M.M.-S.; Dipankar, B.; Brady, K.T. GENDER DIFFERENCES IN THE EFFECT OF EARLY LIFE TRAUMA ON HYPOTHALAMIC–PITUITARY–ADRENAL AXIS FUNCTIONING. Depress. Anxiety 2011, 28, 383–392. [Google Scholar] [CrossRef]
- Bao, A.-M.; Hestiantoro, A.; Van Someren, E.J.W.; Swaab, D.F.; Zhou, J.-N. Colocalization of corticotropin-releasing hormone and oestrogen receptor-α in the paraventricular nucleus of the hypothalamus in mood disorders. Brain 2005, 128, 1301–1313. [Google Scholar] [CrossRef]
- Wang, S.-S.; Kamphuis, W.; Huitinga, I.; Zhou, J.-N.; Swaab, D.F. Gene expression analysis in the human hypothalamus in depression by laser microdissection and real-time PCR: The presence of multiple receptor imbalances. Mol. Psychiatry 2008, 13, 786–799. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, J.; McEwen, B.S. Sex in the brain: Hormones and sex differences. Dialogues Clin Neurosci 2016, 18, 373–383. [Google Scholar] [PubMed]
- Admin NIH Guidelines. Office of Science Policy. Available online: https://osp.od.nih.gov/biotechnology/nih-guidelines/ (accessed on 13 July 2019).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CORT1 | CORT2 | |||
|---|---|---|---|---|
| Behavioral Test | Females | Males | Females | Males |
| SPT |  | | | |
| NSF |  |  |  | |
| SI |  | | | |
| FST | | | | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, S.; Gureczny, S.; Reisinger, S.N.; Horvath, O.; Pollak, D.D. Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice. Cells 2019, 8, 1018. https://doi.org/10.3390/cells8091018
Berger S, Gureczny S, Reisinger SN, Horvath O, Pollak DD. Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice. Cells. 2019; 8(9):1018. https://doi.org/10.3390/cells8091018
Chicago/Turabian StyleBerger, Stefanie, Sarah Gureczny, Sonali N. Reisinger, Orsolya Horvath, and Daniela D. Pollak. 2019. "Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice" Cells 8, no. 9: 1018. https://doi.org/10.3390/cells8091018
APA StyleBerger, S., Gureczny, S., Reisinger, S. N., Horvath, O., & Pollak, D. D. (2019). Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice. Cells, 8(9), 1018. https://doi.org/10.3390/cells8091018